

 PDF
PDF Citation
Citation Print
Print



Introduction
Tumor-infiltrating lymphocytes (TILs) have consistently been reported to play an important role in breast cancer [1–5]. TILs have a strong prognostic and predictive significance, particularly in triple-negative breast cancer (TNBC). CD8+ cytotoxic TILs are activated by the T cell receptor-recognition of a specific peptide, which is generally generated from endogenous proteins, and are presented by a major histocompatibility complex class I (MHC I) on the surface of tumor cells [6]. The recognition of these peptides by cytotoxic CD8+ TILs triggers a series of events that can result in tumor cell lysis. A better understanding of TILs and related features could facilitate the development of efficient immunotherapeutic approaches in breast cancer.
MHC I proteins are membrane proteins that are expressed on almost all nucleated cells and are encoded by human leukocyte antigen (HLA)-A, -B, and -C genes. The expression of HLAs varies from tissue to tissue and is largely stimulated by interferon (IFN) signaling. The downregulation of HLAs is frequently observed in tumors and is reported to be correlated with disease progression [7]. Aberrant HLA expression in tumor cells might be caused by alterations in HLA gene transcription, the translation of HLA mRNA, or post-translational modifications. Torigoe et al. [8] established a monoclonal anti-pan HLA class I antibody suitable for the immunostaining of formalin-fixed tissue and found a high rate (85%, 35 out of 41 cases) of HLA downregulation in breast cancer compared with other malignancies (20%–42%). Since HLA expression on tumor cells is important for the function of TILs, the downregulation of HLA might compromise the effective immune response in patients with breast cancer. Moreover, recent studies have reported increased IFN signaling in cancer cells and their association with a good response to anthracycline-based chemotherapy in breast cancer [9]. However, HLA expression, the level of IFN signaling activation, and their relationship in normal breast tissue and various subtypes of breast cancer have not been extensively studied.
We previously demonstrated the differential expression of HLA-ABC in breast cancer. HLA-ABC protein expression was negatively correlated with estrogen receptor (ER) protein expression but was not significantly correlated with human epidermal growth factor receptor 2 (HER2) protein expression [10,11]. HLA-ABC expression was higher in TNBC and hormone receptor (HR)−/HER2+ breast cancers than in HR+ breast cancers, was positively correlated with TILs, and was associated with better clinical outcomes in breast cancer patients [1,10,11]. In The Cancer Genome Atlas (TCGA) data analysis, HLA-A gene expression was positively correlated with CD8B gene expression but was not significantly correlated with the total number of mutations. Instead, HLAs, CD3, and CD8 gene expression were positively correlated with IFN receptor genes and the IFN-inducible MxA gene [10]. Additionally, MxA protein expression was higher in TNBC than in other types of breast cancer, was positively correlated with TIL levels, and was associated with better clinical outcomes [12]. Therefore, it can be hypothesized that ER activity, IFN signaling, and MHC I expression regulate one another and influence TIL influx.
The current study aimed to clarify the mechanisms of the association of MHC I with estrogen and IFN signaling.
Materials and Methods
1. Cell lines, cultures, drug treatments, and plasmid transfections
This study used breast cancer cell lines obtained from ATCC, including ERα+ (MCF-7 and T47D) and ERα− (MDA-MB-231). The MCF-7 cells were maintained in Dulbecco’s Modified Eagle Medium (cat No. 11995, Life Technologies, Grand Island, NY) supplemented with 10% fetal bovine serum (cat No. 16000, Invitrogen, Carlsbad, CA) and 1% penicillin/streptomycin (cat No. 15140, Invitrogen). The T47D and MDA-MB-231 cells were grown in Roswell Park Memorial Institute 1640 (Gibco, El Paso, TX) supplemented with 10% fetal bovine serum and 1% penicillin/streptomycin. All cells were cultured at 37°C in the presence of 5% CO2. The cells were starved for 24 hours and treated with ICI (1 to 10 μM, fulvestrant, Sigma-Aldrich, St. Louis, MO), IFN-γ (100 units/mL, R&D Systems, Minneapolis, MN), or β-estradiol (1 nM, Sigma-Aldrich) in 2 mL of medium for an appropriate time. The cells were then used in the protein expression assays.
For the ESR1 plasmid transfection, ERα-cells were plated and cultured in a 6-well plate at 90% confluency and transfected with 2.5 μg of hESR-GFP (cat No. #28230, Addgene, Cambridge, MA) using 3.75 μL of Lipofectamine 3000 reagent (Life Technologies) and 5 μL of P3000 reagent (Life Technologies) per well according to the manufacturer’s protocol.
2. Protein isolation and Western blotting
The cells were lysed with RIPA buffer, and the Pierce BCA Protein Assay Reagent Kit (cat No. 23225, Thermo Fisher, Waltham, MA) was used to measure the protein concentration. Approximately 10 μg of protein was separated by 10% sodium dodecyl sulfate polyacrylamide gel electrophoresisand transferred to a PVDF membrane (Millipore, Bedford, MA). S1 Table summarizes the antibodies used for protein detection. Equal loading of the protein samples was verified with an antibody to β-actin. Immunoreactive signals were detected with the Promega Western Blot Detection System (cat No. W1008, Promega, Madison, WI).
3. Gene Expression Omnibus data analysis
We analyzed one dataset from the Gene Expression Omnibus (GEO) database that included Affymetrix RNA microarray analysis data from MCF-7 breast cancer cells that were transfected with ESR1 shRNA (5′-GCTTCAGGCTACCATTATGttcaagagacataATGGTAGCCTGAAGCttttttacgcgt-3′) (accession No. GDS4061) [13]. The fold changes of ESR1, HLA-A, and MxA mRNA expressions were calculated.
4. Patients and tissue specimens
A total of 126 patients who were diagnosed with ER+/HER2− invasive ductal carcinoma were randomized to receive either estrogen modulator treatment or chemotherapy for 24 weeks as their neoadjuvant systemic therapy [14]. We analyzed the clinicopathologic data of the patients and the HLA-ABC and ER protein expressions and TIL levels in the pre-neoadjuvant biopsy tissues and the post-neoadjuvant resected tissues.
5. Histological evaluation
The histologic type was defined based on the 2019 World Health Organization classification criteria, and the histologic grade was assessed using the modified Bloom-Richardson classification [15]. The hematoxylin and eosin–stained slides were histopathologically analyzed for TILs (defined as the percentage of the invasive carcinoma’s stroma that was infiltrated by lymphocytes in 10% increments; if less than 10% of the stroma was infiltrated by TILs, 1% or 5% criteria were used; all available full sections were evaluated), histological subtype and grade, tumor size, pT category, pN category, and lymphovascular invasion [1,16]. The tumor response to neoadjuvant systemic therapy was evaluated based on the Miller-Payne grade (1, no change; 2, up to 30% reduction; 3, 30%–90% reduction; 4, more than 90% reduction; 5, no residual malignant cells) [17]. A pathologic complete response (pCR) was defined as the absence of residual invasive cancer cells in the breast and lymph nodes [18].
6. Tissue microarray construction and immunohistochemical evaluation
All 126 patients were checked for ER, progesterone receptor (PR) and HER2 expression and Ki-67 labeling index both in the pre-neoadjuvant biopsy tissues and the post-neoadjuvant resected tissues by immunohistochemistry, except 9 cases in whom the residual tumor cells were few or even did not exist at all in post-neoadjuvant resected tissues. ER and PR levels were regarded as positive if there was at least 1% positive tumor nuclei staining. Additionally, the Allred score, which is the sum of the intensity score (0–3) and the proportion score (0–5), was calculated for the ER and PR. HR+ tumors were defined as those determined to be ER-positive and/or PR-positive. HER2-overexpressing tumors were defined as those with scores of 3+ according to the immunohistochemistry or gene amplification by silver in situ hybridization. Ki-67 labeling index in the tumor cells was measured by eyeball estimation and dichotomized into < 20% and ≥ 20%.
Among the 126 patients, 56 were available for formalin-fixed, paraffin-embedded tissue blocks of both biopsies and resected specimens at the time of this study. Each resected tissue sample was arrayed in three 1-mm diameter cores to minimize tissue loss and overcome tumor heterogeneity. Full sections of biopsy tissues and tissue microarray sections of the resected tissues were stained with an automatic immunohistochemical staining device (Benchmark XT, Ventana Medical Systems, Tucson, AZ). The HLA-ABC staining was semi-quantitatively evaluated as the H-score, which is the product of the actual percentage of positive-stained cells and the intensity score (0–3); the H-score can range from 0–300. We then categorized HLA-ABC expression in tumor cells as one of two levels (negative, H-score < 50; positive, H-score ≥ 50).
S2 Table summarizes the antibodies used for the immunohistochemical staining.
7. Statistical analysis
All statistical analyses were performed using R ver. 3.2.3 [19]. The Wilcoxon rank-sum test, chi-square test, Fisher exact test, and log-rank test were used as appropriate. All tests were two-sided, and statistical significance was set at 5%.
Results
1. The expression of HLA-ABC in breast cancer cell lines treated with ICI or IFN-γ
We evaluated the expression of the HLA-ABC protein in the ER-positive cell lines (MCF-7 and T47D) and in an ER-negative cell line (MDA-MB-231) by Western blot analysis. The baseline HLA-ABC protein expression was higher in the MDA-MB-231 cell line than in the MCF-7 and T47D cell lines (Fig. 1A). Next, we treated the MCF-7 cells with 1 to 10 μM ICI, which is an ER downregulator, for 24 or 48 hours. The ER protein expression decreased and HLA-ABC increased with ICI treatment under all conditions (Fig. 1B). When the MCF-7 cells were treated with 100 units/mL of IFN-γ with or without 1 nM of estradiol for 48 hours, the HLA-ABC protein expression was markedly increased (Fig. 1C).
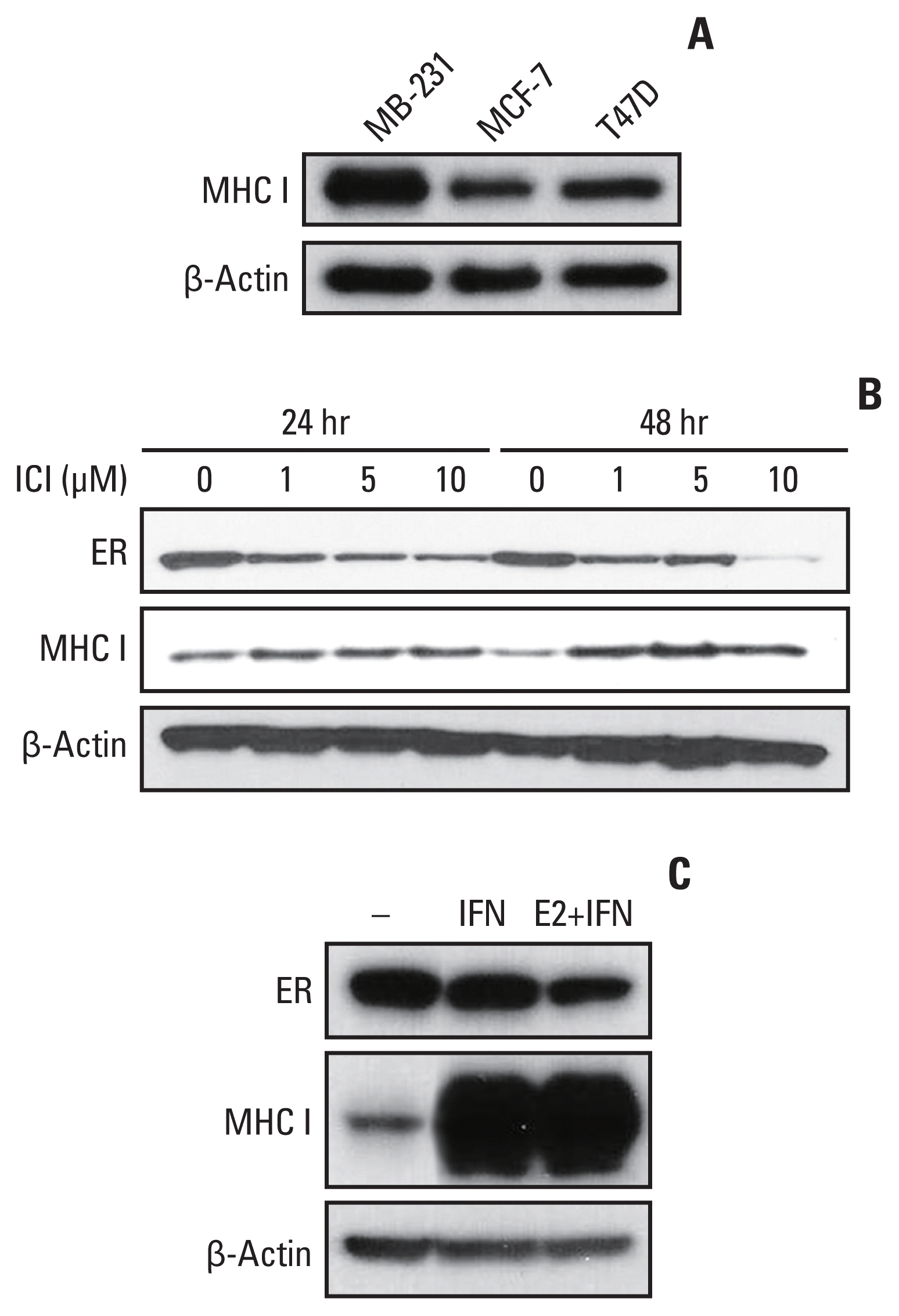
Fig. 1
The effect of estrogen signaling and interferon signaling on HLA-ABC expression. (A) The baseline HLA-ABC protein expression is higher in ER-negative MDA-MB-231 cell lines than in ER-positive MCF-7 and T47D cell lines. (B) After 24 or 48 hours of ICI treatment (1 to 10 μM), the ER protein expression decreased, and HLA-ABC protein increased in MCF-7 cells. (C) The HLA-ABC protein expression increased in MCF-7 cells 48 hours after 100 units/mL of IFN-γ treatment. ER, estrogen receptor; HLA, human leukocyte antigen; IFN, interferon; MHC I, major histocompatibility complex class I.
![]()
2. The expression of HLA-ABC in the ER-negative breast cancer cell line transfected with hESR-GFP
We also observed changes in the HLA-ABC protein expression in the ER-negative MDA-MB-231 breast cell line. Twenty-four or 48 hours after transfection with the hESR-GFP plasmid, the MDA-MB-231 cells showed ER expression and decreased HLA-ABC protein levels (Fig. 2A). When the MDA-MB-231 cells were treated with 100 units/mL of IFN-γ with or without 1 nM of estradiol for 48 hours, HLA-ABC protein expression was markedly increased (Fig. 2B).
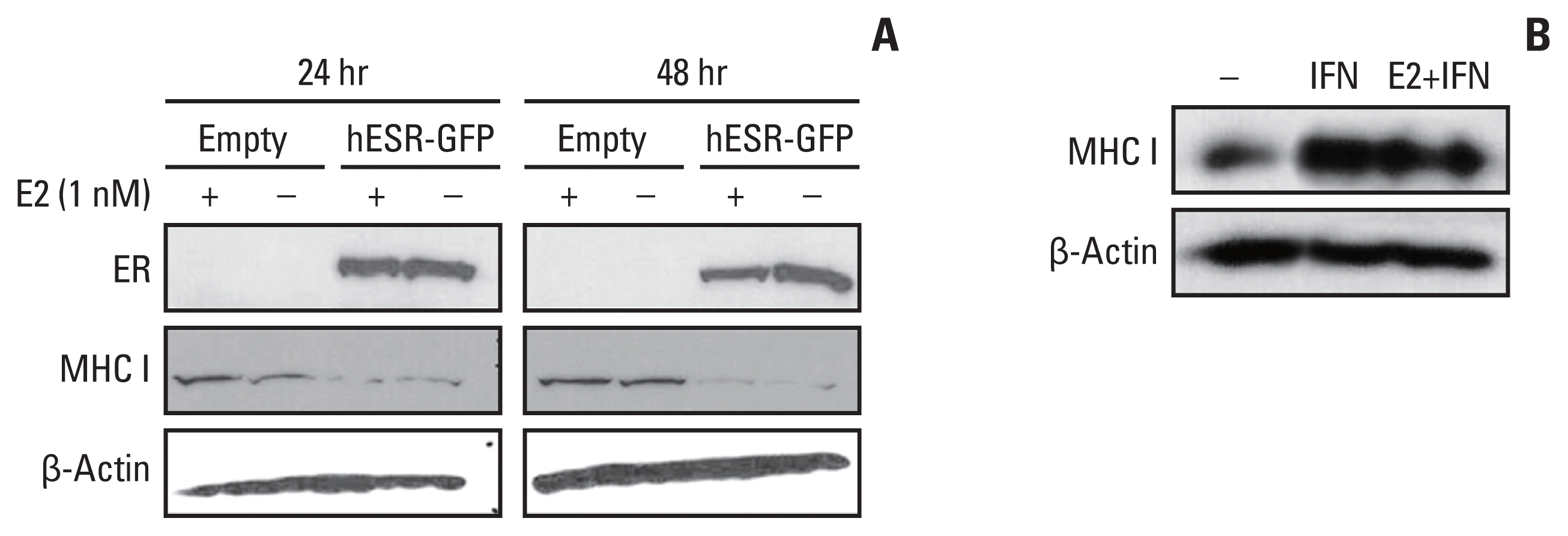
Fig. 2
HLA-ABC protein expression in the MDA-MB-231 cells. (A) Twenty-four or 48 hours after transfection with the hESR-GFP plasmid, MDA-MB-231 cells show ER expression and decreased HLA-ABC protein levels. (B) The HLA-ABC protein expression was markedly increased 48 hours after 100 units/mL of IFN-γ treatment in MDA-MB-231 cells. ER, estrogen receptor; HLA, human leukocyte antigen; IFN, interferon; MHC I, major histocompatibility complex class I.
![]()
3. The change of HLA-A and MxA mRNA expressions in breast cancer cell lines in the GEO data
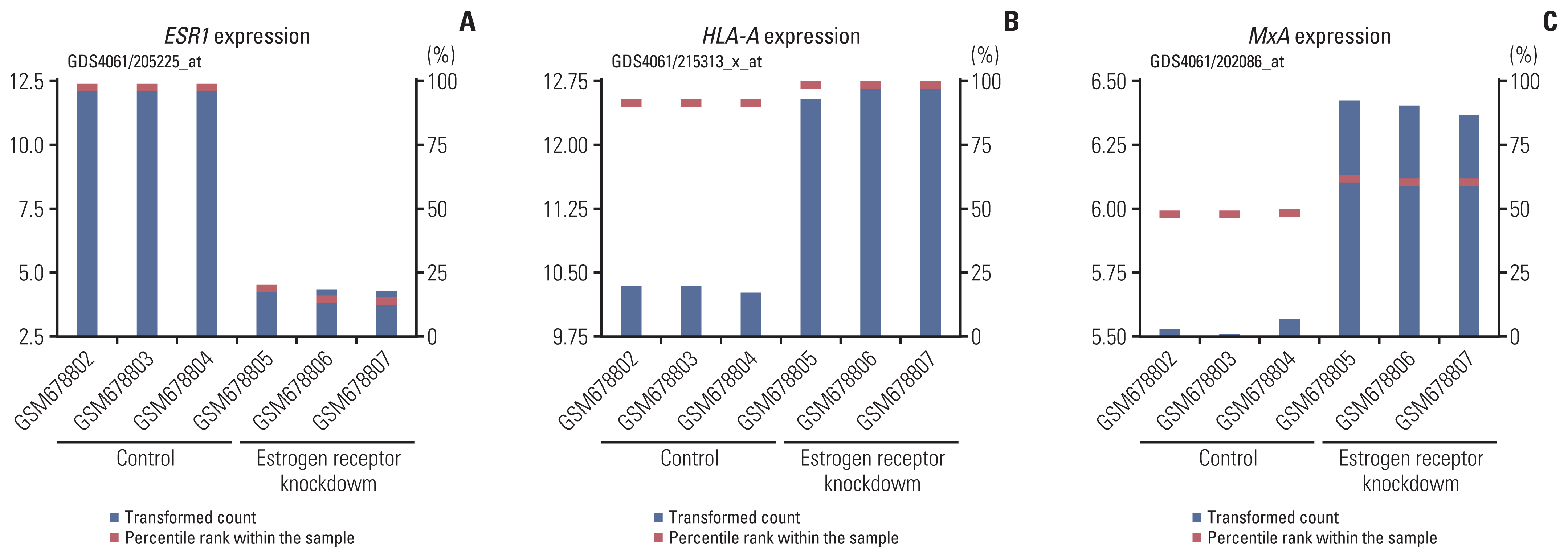
We also analyzed the HLA-A and MxA mRNA expressions in breast cancer cells from the GEO database. When the MCF-7 cells were transfected with ESR1 shRNA, the ESR1 mRNA expression decreased, and the HLA-A and MxA mRNA expressions increased (Fig. 3). The fold changes were 0.360, 1.225, and 1.156 for ESR1, HLA-A, and MxA, respectively.
4. The changes of ER and HLA-ABC expressions, Ki-67 labeling index and TIL levels after neoadjuvant chemotherapy or estrogen modulators in breast cancer patients
The clinicopathologic characteristics of the 126 patients who randomly received chemotherapy (adriamycin and cyclophosphamide) or estrogen modulator treatment (tamoxifen and goserelin) as their neoadjuvant systemic therapy and following surgical resection were analyzed (Table 1). Before the neoadjuvant therapy, there were no differences between the two groups in patient age, cT and cN categories, ER Allred score, Ki-67 labeling index, and TIL levels in biopsy specimens. After neoadjuvant therapy, no significant difference was observed in the pCR ratio (p=0.058). However, the chemotherapy group showed significantly lower ypT and ypN categories (p < 0.001 and p=0.016, respectively) and a more reduced tumor burden according to the Miller-Payne grade (p < 0.001). The estrogen modulator group showed more frequent lymphovascular invasion (p=0.046), a lower ER Allred score (p=0.027), higher Ki-67 labeling index (p=0.004) and higher TIL levels (p < 0.001) in resected specimens than the neoadjuvant chemotherapy group (Figs. 4, 5).
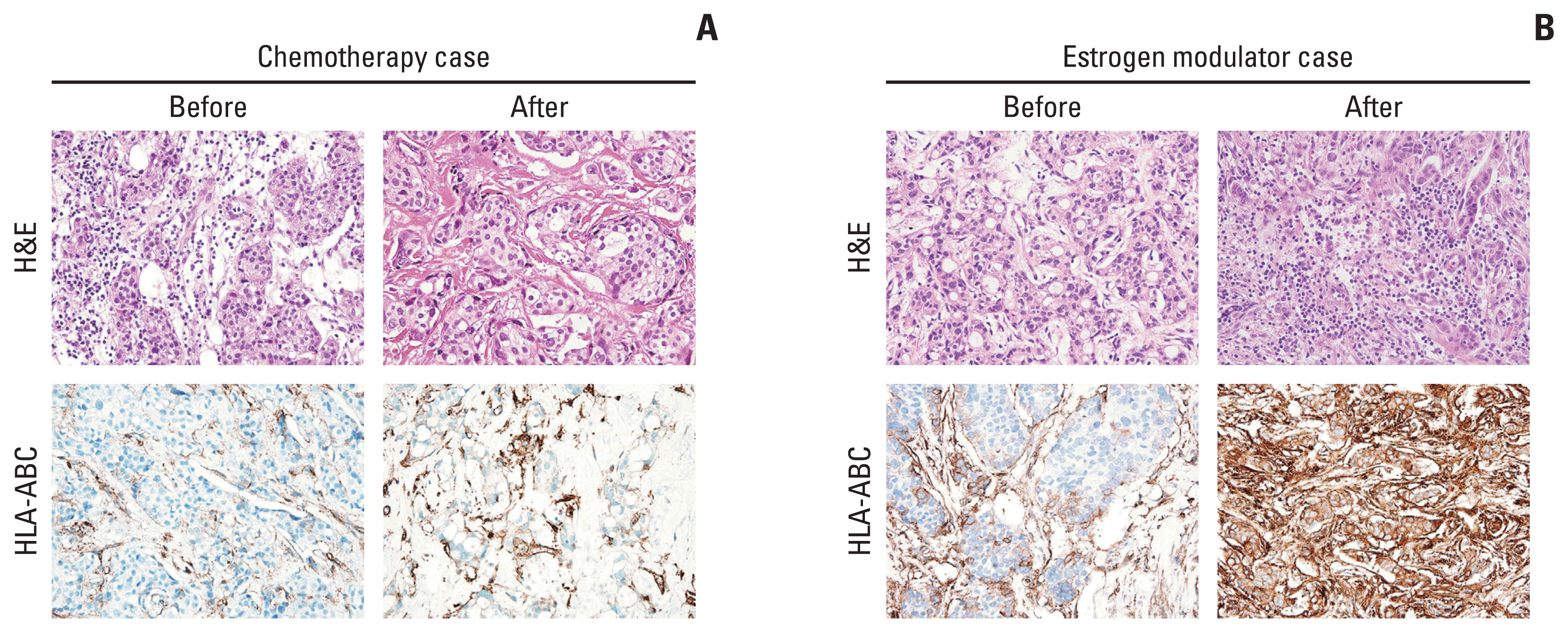
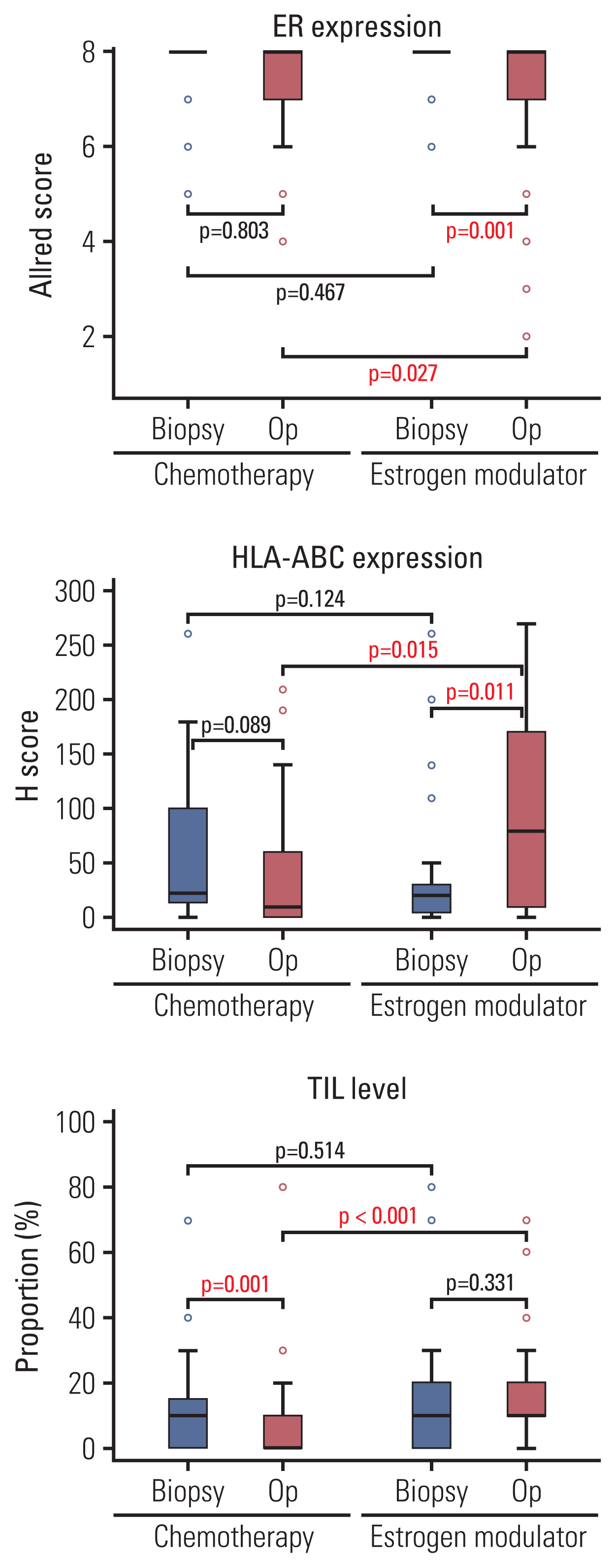
Fig. 4
The microscopic images of breast tissue before and after chemotherapy or estrogen modulator treatment. In the patient who received neoadjuvant chemotherapy (A), the quantity of TILs decreased and HLA-ABC expression was similar in the resection specimen compared with the pre-neoadjuvant biopsy specimens. By contrast, the TIL levels and HLA-ABC expression increased after estrogen modulator treatment in the patient who received estrogen modulator treatment (Nolvadex and Zoladex) (B) (A and B, ×400). HLA, human leukocyte antigen; TIL, tumor-infiltrating lymphocyte.
![]()
Fig. 5
The changes of ER and HLA-ABC expression and TIL levels. The TIL levels significantly decreased after chemotherapy, while the expression of ER decreased after estrogen modulator treatment. Compared with the chemotherapy group, HLA-ABC expression increased significantly after estrogen modulator treatment (Op). ER, estrogen receptor; HLA, human leukocyte antigen; Op, operation; TIL, tumor-infiltrating lymphocyte.
![]()
Table 1
Comparison of clinicopathologic variables according to neoadjuvant systemic therapy in breast cancer patients
| Variable | Neoadjuvant systemic therapy | p-valuea) | |
|---|---|---|---|
| Chemotherapy (n=65) | Estrogen modulator (n=61) | ||
| Pre-neoadjuvant | |||
| Age at diagnosis (yr) | 43 (38–46) | 42 (38–46) | 0.784 |
| cT | |||
| 1 | 12 (18.6) | 7 (11.5) | 0.522 |
| 2 | 42 (64.6) | 42 (68.9) | |
| 3 | 10 (15.4) | 12 (19.7) | |
| 4 | 1 (1.5) | 0 | |
| cN | |||
| 0 | 1 (1.5) | 0 | > 0.99 |
| 1 | 54 (83.1) | 52 (85.2) | |
| 2 | 4 (6.2) | 4 (6.6) | |
| 3 | 6 (9.2) | 5 (8.2) | |
| ER Allred score in biopsy | 8 (8–8) | 8 (8–8) | 0.467 |
| Ki-67 labeling index in biopsy | |||
| < 20% | 24 (36.9) | 18 (29.5) | 0.488 |
| ≥ 20% | 41 (63.1) | 43 (70.5) | |
| HLA expression in biopsy | |||
| Negative | 17 (65.4) | 24 (80.0) | 0.243 |
| Positive | 9 (34.6) | 6 (20.0) | |
| TIL level in biopsy | 10 (0–12.5) | 10 (0–20) | 0.514 |
| Post-neoadjuvant | |||
| Histologic grade | |||
| 1 | 4 (6.9) | 3 (5.0) | 0.358 |
| 2 | 46 (79.3) | 53 (88.3) | |
| 3 | 8 (13.8) | 4 (6.7) | |
| ypT | |||
| 0 | 7 (10.8) | 1 (1.6) | < 0.001 |
| 1 | 32 (49.2) | 12 (19.7) | |
| 2 | 20 (30.8) | 37 (60.7) | |
| 3 | 6 (9.2) | 9 (14.8) | |
| 4 | 0 | 2 (3.3) | |
| ypN | |||
| 0 | 8 (12.3) | 2 (3.3) | 0.016 |
| 1 | 39 (60.0) | 27 (44.3) | |
| 2 | 15 (23.1) | 23 (37.7) | |
| 3 | 3 (4.6) | 9 (14.8) | |
| Miller-Payne grade | |||
| 1 | 2 (3.1) | 20 (32.8) | < 0.001 |
| 2 | 10 (15.4) | 24 (39.3) | |
| 3 | 36 (55.4) | 14 (23.0) | |
| 4 | 10 (15.4) | 2 (3.3) | |
| 5 | 7 (10.8) | 1 (1.6) | |
| Pathologic complete response | |||
| No | 60 (92.3) | 61 (100) | 0.058 |
| Yes | 5 (7.7) | 0 | |
| LVI | |||
| Absent | 36 (55.4) | 22 (36.1) | 0.046 |
| Present | 29 (44.6) | 39 (63.9) | |
| ER Allred score in resected tissue | 8 (7.25–8) | 8 (7–8) | 0.027 |
| Ki-67 labeling index in resected tissue | |||
| < 20% | 50 (86.2) | 36 (61.0) | 0.004 |
| ≥ 20% | 8 (13.8) | 23 (39.0) | |
| HLA expression in resected tissue | |||
| Negative | 17 (68.0) | 10 (35.7) | 0.038 |
| Positive | 8 (32.0) | 18 (64.3) | |
| TIL in resected tissue | 0 (0–10) | 10 (10–20) | < 0.001 |
![]()
Next, we evaluated HLA-ABC expression in both the biopsy and resection tissues of 56 patients by immunohistochemistry (Table 1). Before neoadjuvant therapy, the HLA-ABC positivity in tumor cells did not differ significantly between the two groups (p=0.243). After neoadjuvant systemic therapy, however, the estrogen modulator group showed higher HLA-ABC positivity than the chemotherapy group (p=0.038).
The overall survival and recurrence-free survival between the two groups did not differ significantly (p=0.396 and p=0.758, respectively) (S3 Fig.).
Discussion
Recently, the significance of TILs, particularly that of CD8+ cytotoxic T cells, in breast cancer has been revealed. Higher TIL level is known to be associated with longer patient survival and better response to chemotherapy [4,11,20]. Higher TIL level also correlates with programmed death-ligand 1 (PD-L1) expression [21], which may predict response to immune checkpoint inhibitor treatment [22]. Immune checkpoint inhibitors pembrolizumab and atezolizumab recently have been approved in TNBC with PD-L1 expression by FDA. However, HR+ breast cancers are still considered to be immunologically cold, and it is not yet hopeful whether immunotherapy can be effective in patients with those cancers [23].
The expression of MHC I proteins on the tumor cell surface is essential for CD8+ T cells to act, and several reports have documented the positive relationship between TIL levels and MHC I expression. We previously reported that TILs were more abundant in tumors with a stronger expression of HLA-ABC. We also reported that the HLA-ABC expression of the tumor cells was positively correlated with TIL levels in consecutive series of primary breast cancers and TNBC cohorts [10]. Although some may think that high HLA-ABC expression is due to a high mutation rate and more immunogenic mutations, we previously revealed that the total number of mutations was not associated with HLA-A expression in the tumor, and Spranger et al. [24] also reported that the density of nonsynonymous somatic mutations is not significantly associated with T cell related gene expression.
Instead, some evidence has indicated that MHC I expression is related to ER expression and IFN signaling. In our previous study, HLA-ABC expression was negatively associated with ER expression in a consecutive breast cancer cohort and normal breast tissue, and HLA mRNA expression was positively correlated with IFN-associated gene expression in a TCGA and Cancer Cell Line Encyclopedia data analysis [10]. We also reported that the high expression of IFN-mediator MxA in the tumor cells was positively associated with TIL levels, CD8+ cell number, and stronger HLA-ABC expression and was an independent prognostic factor for better disease-free survival in breast cancer [12]. Although one article has reported that ERα signaling modulates IFN-γ inducible MHC II expression through class II transactivator in breast cancer cells [25], the mechanism of the relationship between HLA-ABC expression, ER activity, and IFN signaling in breast cancers has never been reported.
In the current study, we revealed that the expressions of HLA-ABC protein and HLA-A mRNA in breast cancer cells are negatively affected by ER signaling in vitro. We insist that this is a novel and important finding in addition to previously known tumor-intrinsic oncogenic pathways that have been suggested to be associated with the reduction of immune reactions, such as the WNT/β-catenin pathway, the mitogen-activated protein kinase pathway, the phosphoinositide 3-kinase/AKT pathway, MYC upregulation, and CDK4/6 activation [26]. The baseline HLA-ABC protein expression was higher in ER-negative cells than in ER-positive cells, and HLA-ABC protein expression was increased after ICI treatment and decreased after ER overexpression. HLA-A mRNA expression was also increased after ESR1 shRNA transfection. We also revealed a positive association between HLA-ABC and IFN signaling and a negative association between ER and IFN signaling. Breast cancer cells showed increased HLA-ABC protein expression after IFN treatment and increased MxA mRNA expression after ESR1 shRNA transfection. This is the first study that analyzed the relationship between HLA-ABC expression, ER activity, and IFN signaling in breast cancer in vitro.
We also analyzed the breast cancer tissues of patients who received either neoadjuvant chemotherapy or estrogen modulator therapy. We revealed that the HLA-ABC expression in breast cancer cells increased and ER expression decreased after estrogen modulator treatment compared with chemotherapy. Estrogen modulator treatment negatively regulate ER signaling in breast cancer. By combining the results of the in vitro experiments, our data suggested that ER signaling was downregulated by estrogen modulator treatment, and this caused the increase of HLA-ABC expression in tumor cells. We also analyzed the TIL levels in biopsies and resection tissues from the two groups. TIL levels were significantly decreased in resected tissue after neoadjuvant chemotherapy compared with biopsy tissue, which is concordant with a previous publication [27]. Although the HLA-ABC expression in tumor cells was increased in the estrogen modulator treatment group, we did not identify a significant change in TIL levels in this group. This is possibly due to the insufficient duration of neoadjuvant estrogen modulator therapy. Further studies with a longer duration of neoadjuvant treatment are necessary. However, estrogen modulator therapy group still showed significantly higher TIL level in resected tissue than chemotherapy group. Park et al. [28] reported that immune-stimulation after neoadjuvant chemotherapy is associated with pCR in breast cancer. Therefore, we suggest that higher HLA-ABC expression and TIL level in estrogen modulator therapy group may have a positive effect on prognosis of patients.
We found that Ki-67 labeling index was decreased both after neoadjuvant chemotherapy and after neoadjuvant estrogen modulator treatment, but chemotherapy group showed significantly lower Ki-67 labeling index than estrogen modular treatment group in resected tumor tissue. This might be explained by the fact that conventional chemotherapeutic agents are generally toxic to proliferative cells. Considering that chemotherapy group showed remarkable decrease of Ki-67 labeling index but did not show significant change of HLA-ABC expression, we suggest that increase of HLA-ABC expression in estrogen modulator group is rather associated with downregulation of ER signaling than decreased proliferation activity.
This study has several limitations. First, our data do not include in vitro results with estrogen treatment, the key molecule in ER signaling, or with tamoxifen treatment, an important selective ER modulator drug. Second, molecular mechanisms and mediators on how HLA-ABC expression and IFN signaling are regulated by ER signaling must be identified. Therefore, further experiments are necessary to more precisely define these molecular mechanisms. We suggest that more significant results might be obtained by future studies with more patient samples and a longer follow-up duration.
In conclusion, this study demonstrated that MHC I expression and TIL levels were affected by ER pathway modulation and IFN treatment. Therefore, we suggest that downregulation of ER signaling might induce immune reaction in HR+ breast cancer. Further studies elucidating the mechanism of MHC I regulation could suggest a way to boost TIL influx in cancer and increase the efficacy of immunotherapy in treatment of HR+ breast cancer patients.
 XML Download
XML Download