

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Mitochondria play a pivotal function in cellular energy supply, where adenosine triphosphate (ATP) is generated by the mitochondrial respiratory chain in a process referred to as oxidative phosphorylation [1]. A decline in mitochondrial function, reflected by diminished electron transport chain activity, is related to many common degenerative diseases, including cancer [2], diabetes [3], neurodegeneration [4], and other aging-associated diseases [1].
One of the most informative mitochondrial function tests is cellular respiration quantification [5]. In particular, the oxygen consumption rate (OCR), expressed in pmol/min, can estimate the ability of mitochondria to synthesize ATP and perform its functions [6]. For epidemiological studies that have practical limitations concerning the availability of bio-specimens, OCR levels of leukocytes and platelets can be used as a surrogate marker to investigate mitochondrial function in humans [7]. Several small-scale clinical studies have measured the OCR of leukocytes or platelets to compare OCR levels between patients and control groups [8–13].
Recently, low-dose environmental chemicals have been suspected to induce mitochondrial dysfunction [14–16]. Among them, persistent organic pollutants (POPs), especially organochlorine pesticides (OCPs) and polychlorinated biphenyls (PCBs), have garnered interest because chronic exposure to low-dose POPs is a new risk factor for many mitochondrial dysfunction-related diseases, including type 2 diabetes mellitus (T2DM) and dementia [17,18]. As typical examples of lipophilic chemical mixtures, POPs exhibit long half-lives with strong lipophilicity, biodegradation resistance, and bioaccumulation in the fatty tissue of living organisms [18].
To the best of our knowledge, no previous study has evaluated the possible relationship between POPs and OCR in humans. This study aimed to evaluate the association between OCR and serum OCP and PCB concentrations in the general population. In addition, we evaluated the effects of low-dose OCPs and PCBs on OCR in in vitro cell models.
METHODS
Study population
This study was conducted with 323 subjects aged between 30 and 70 years who visited Kyungpook National University Hospital (Daegu, Republic of Korea) for medical checkups between November 2015 and February 2018. Patients diagnosed with malignant tumors, myocardial infarction, stroke, or renal failure within the last 5 years were excluded from the study. The study was approved by the Institutional Review Board of Kyungpook National University Hospital (IRB No. KNUH 2013-12-016). Informed consent was obtained from all participants.
Human study
Demographic factors, physical measurements, and health-related behaviors
Trained interviewers collected information on demographic factors, health-related behaviors, and physical measurements. All subjects were asked to fast overnight for at least 8 hours before venous blood sampling. Approximately 25 mL of fresh whole blood was collected from each subject, and 20 mL was immediately sent to a laboratory for OCR analysis. Serum was collected after centrifugation of 5 mL of blood, and the serum samples were stored in a freezer at −80°C until POP analysis. The height and weight of all subjects were measured while the subjects wore light clothes and no shoes. Body mass index (BMI) was calculated as weight divided by the square of the height. T2DM and hypertension were defined based on the physician’s diagnosis.
OCR measurement
1) Isolation of platelets and peripheral blood mononuclear cells
Platelets and peripheral blood mononuclear cells (PBMCs) were isolated from 20 mL of fresh blood within 2 hours of blood collection. For platelet separation, platelet-rich plasma was obtained by centrifugation of blood at 500 relative centrifugal force (rcf) for 15 minutes. The platelet-rich plasma was centrifuged for an additional 10 minutes at 1,500 rcf to collect platelets. Isolated platelets were cleaned and reconstituted with phosphate-buffered saline (P4417, Sigma-Aldrich, St. Louis, MO, USA) containing 1 μg/mL prostaglandin I2 (P6188, Sigma-Aldrich), which inhibits platelet activation.
For PBMC separation, density-gradient separation using Histopaque-1077 (10771, Sigma-Aldrich) and isolation by the buffy coat was performed. Tubes containing blood from each subject were centrifuged at 700 rcf for 30 minutes at room temperature. To isolate PBMCs, a magnetic-activated cell sorting separator was used [19]. The isolated PBMCs were washed with L-glutamine (Gibco 11835-030, Thermo Fisher Scientific, Waltham, MA, USA) and cultured in RPMI 1640 medium (Gibco 11879-020, Thermo Fisher Scientific) supplemented with penicillin (100 U/mL), streptomycin (100 mg/mL), and 10% fetal bovine serum.
2) Bioenergetic profiles of platelets and PBMCs
Respirometry of platelets and PBMCs was performed using an XF24 analyzer (Seahorse Bioscience, Billerica, MA, USA). We used freshly isolated platelets and PBMCs obtained directly after blood collection without cryopreservation. Platelets and PBMCs were immobilized onto a Cell-Tak (354241, Corning Inc., Corning, NY, USA) coated Seahorse XF24 cell culture microplate in XF assay medium supplemented with 10 mM glucose (103576-100, Agilent Technologies, Santa Clara, CA, USA) and 1 mM pyruvate (103578-100, Agilent Technologies). Bioenergetic profiles were analyzed in quadruplicates. Oligomycin (1.5 μM, Sigma-Aldrich), carbonyl cyanide-4-(trifluor-omethoxy), phenylhydrazone (3 to 4.5 μM, C2759, Sigma-Aldrich), and rotenone (1 μM, A8674, Sigma-Aldrich) were used to inhibit mitochondrial respiration in this assay. We measured basal, maximal, and reserve OCRs, which were normalized to the total protein amounts.
Measurement of POPs
Serum samples stored in a freezer at −80°C were transferred to a laboratory at Hanyang University (Ansan, Korea) for POP analysis. The samples were purified and concentrated for analysis. POP analysis was performed using a high-resolution gas chromatography/high-resolution mass spectrometry system (HRGC/HRMS; JMD-700D, JEOL, Tokyo, Japan). A total of 35 types of POPs (17 PCBs and 18 OCPs) were identified. The PCBs identified were PCB18, PCB28, PCB33, PCB52, PCB54, PCB70, PCB101, PCB105, PCB118, PCB128, PCB138, PCB153, PCB170, PCB180, PCB187, PCB195, and PCB206. The OCPs identified were α-hexachlorocyclohexane (HCH), β-HCH, γ-HCH, δ-HCH, o,p′-dichlorodiphenyldichloroethylene (DDE), p,p′-DDE, o,p′-dichlorodiphenyldichloroethane (DDD), p,p′-DDD, o,p′-dichlorodiphenyltrichloroethane (DDT), p,p′-DDT, trans-chlordane, cis-chlordane, oxychlordane, trans-nonachlor, cis-nonachlor, heptachlor, heptachlor epoxide, and hexachlorobenzene. Among them, five PCBs (PCB118, PCB138, PCB153, PCB180, and PCB187) and four OCPs (β-HCH, p,p′-DDT, p,p′-DDE, and trans-nonachlor) for which at least 60% of the subjects had concentration values above the limit of detection (LOD) were included in statistical analyses. The LOD was defined as three times the signal-to-noise ratio. The detection rates and distributions of the selected POP concentrations are provided in Supplementary Table 1. When both wet and lipid-adjusted concentrations were evaluated, their results were similar, so we present wet concentrations as the main result.
In vitro experiment
The effects of low-dose β-HCH (CAS No. 1024-57-3, AccuStandard, New Haven, CT, USA), p,p′-DDT (CAS No. 50-29-3, AccuStandard), and Arochlor-1254 (CAS No. 11097-69-1, AccuStandard) on the OCR were evaluated in human platelets, human PBMCs, and Jurkat T-cells (Clone E6-1, ATCC TIB-152, ATCC, Manassas, VA, USA), using an XF96 analyzer (Seahorse Bioscience). The concentration range of the three compounds was 0.1 pM to 100 nM. As cell viability in in vitro environments differed by cell type, treatment duration varied by cell type. Since metabolic and OCR control of platelets is difficult in in vitro environments owing to the lack of a nucleus in the cells, the OCR for platelets was measured after treatment for 6 and 24 hours. The PBMC OCR was measured at 24 and 72 hours, while that for Jurkat T-cells was measured after treatment for 2 and 7 days. Viability was assessed using the Cell Counting Kit 8 (Dojindo Laboratories, Kumamoto, Japan).
Statistical analyses
We primarily used the summary measures of PCBs (∑PCBs) and OCPs (∑OCPs) because the general population is simultaneously exposed to a mixture of these compounds, and there are high correlations among serum concentrations of individual compounds. We presented results of basal OCR for the human studies because other OCRs did not show clear patterns. Since the basal platelet and PBMC OCR values showed skewed distributions, logarithmic changes were used for statistical analyses. The final results were presented as geometrical means of OCR values.
The summary measures were determined by summing the ranks of the individual congeners of the five PCBs and four OCPs, and classified into quintile groups. The last quintile included a broad range of concentrations; therefore, we further categorized the last quintile using 90th and 95th percentile cut-off points. Summary measures using absolute concentrations of each compound were not considered since these measures were largely determined by a few compounds with high concentrations. Also, we provided results on individual compounds.
According to quintiles of summary measures, basal platelet and PBMC OCR values were estimated using the general linear model. The covariates were age (continuous, years), sex, BMI (continuous, kg/m2), current smoker (yes, no), current drinker (yes, no), moderate or vigorous exercise (yes, no), physician-diagnosed hypertension (yes, no), and physician-diagnosed T2DM (yes, no). Furthermore, we performed stratified analyses by age (<60, ≥60 years), sex (men, women), BMI (<25, ≥25 kg/m2), T2DM (yes or no), and hypertension (yes or no). Statistical analyses were performed using SAS version 9.4 (SAS Inc., Cary, NC, USA).
RESULTS
General characteristics of study subjects
The platelet OCR was measured in 323 study subjects, while PBMC OCR was measured in 261 subjects. The mean±stan-dard deviation of basal platelet and PBMC OCR was 9.38± 5.47 and 43.08±52.54 pmol/min/μg, respectively. There was no correlation between these two measures (r=0.05, P=0.420). Supplementary Table 2 shows demographic, health behavior-related, and clinical characteristics of study subjects. Among the 323 participants, the mean age was 54.5 years, 47.7% were men, and the mean BMI was 24.8 kg/m2. The characteristics of the 261 subjects with information on PBMC OCR were similar to those of all study subjects.
Association between general characteristics and basal OCR
Table 1 shows the mean values of the basal platelet or PBMC OCR according to general characteristics. In the case of platelets, women showed significantly lower basal OCR levels than men (P=0.010), and subjects with hypertension had significantly lower basal OCR levels than those without hypertension (P=0.018). However, age, BMI, cigarette smoking, alcohol consumption, exercise, and T2DM were not associated with the basal platelet OCR level.
On the other hand, the pattern of the mean values of basal PBMC OCR according to general characteristics differed from that of mean values of basal platelet OCR. Age was significantly associated with basal PBMC OCR level (P=0.013); subjects aged ≥60 years showed higher OCR levels than those aged <60 years. Moreover, diabetic subjects had significantly lower basal OCR levels than those without T2DM (P=0.011). However, sex, BMI, cigarette smoking, alcohol consumption, exercise, and hypertension, were not significantly associated with the basal PBMC OCR levels.
Association between serum concentrations of POPs and basal OCR
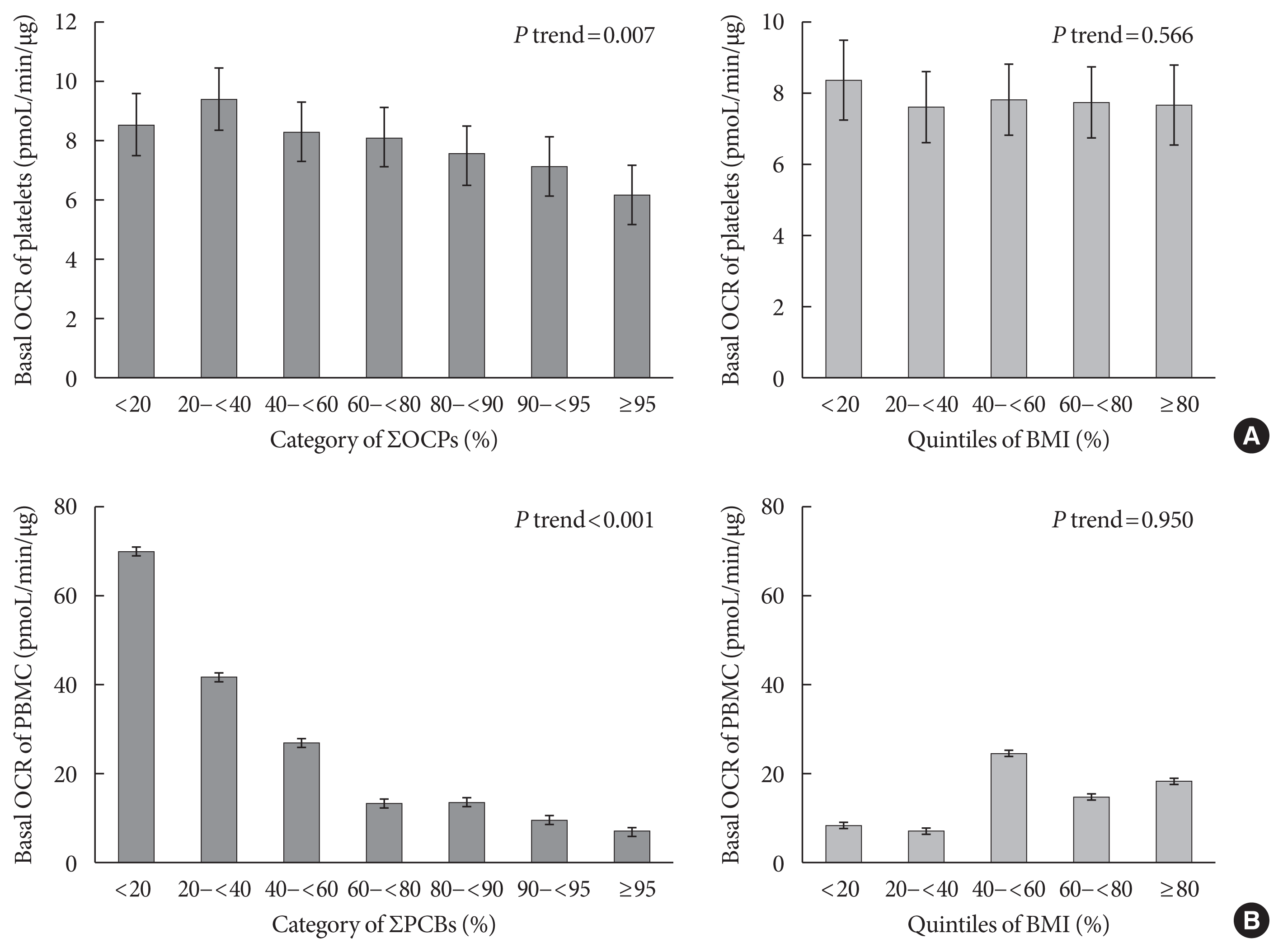
Table 2 shows the association of summary measures of PCBs or OCPs with basal platelet or PBMC OCR levels. In platelets, the summary measures of OCPs were significantly and inversely associated with mean basal OCR levels. For example, according to the quintiles of ∑OCPs, the mean basal platelet OCR levels were 8.6, 9.6, 8.2, 8.0, and 7.1 pmol/min/μg (P trend=0.019). This inverse association persisted even after adjustment for age, gender, BMI, cigarette smoking, alcohol consumption, exercise, hypertension, and T2DM. However, there was no association between ∑PCBs and platelet OCR levels.
Meanwhile, PCB summary measures demonstrated remarkably strong inverse associations with the mean basal PBMC OCR levels (Table 2). According to the quintiles of ∑PCBs, the mean basal OCR levels were 70.1, 41.0, 26.4, 13.2, and 10.5 pmol/min/μg (P trend <0.001). Adjusting for all covariates did not change the strong inverse associations. On the contrary, there was no association between the ∑OCP serum concentrations and the mean basal PBMC OCR levels.
In addition, we compared POPs with obesity in the relationships with platelet or PBMC OCR levels in detail (Fig. 1). For these comparisons, the last quintile of OCP or PCB summary measures were further categorized with 90% and 95% cut-off points. Although POPs showed clearer decreasing trends in the last quintile, BMI was not related to platelet or PBMC OCR levels. Additional analyses focusing on individual compounds revealed inverse associations between OCPs and basal platelet OCR levels for β-HCH and trans-chlordane, whereas the inverse associations between PCBs and basal PBMC OCR levels were observed for all PCB congeners (Supplementary Table 3).
Stratification analysis
Table 3 presents the associations between the summary measure of OCPs and mean platelet OCR levels stratified by age (<60, ≥60 years), sex (men, women), BMI (<25, ≥25 kg/m2), T2DM (yes, no), and hypertension (yes, no). The inverse associations were observed in all subjects; however, they were pronounced in subjects aged ≥60 years, men, BMI <25 kg/m2, and subjects without T2DM. On the other hand, strong inverse association between the summary measures of PCBs and mean basal PBMC OCR levels was observed in all subgroups stratified by age, sex, BMI, T2DM, and hypertension (Table 4).
In vitro study
Fig. 2 shows a summary of the in vitro study. Cells were treated with the three POPs Arochlor-1254, β-HCH, and p,p′-DDT, ranging in concentration from 0.1 pM to 100 nM, mimicking low-dose exposure that was similar to that in humans. The OCR levels were measured at 6 and 24 hours for human platelets, 24 and 48 hours for human PBMCs, and days 2 and 7 for Jurkat T-cells owing to different viabilities by cell type. The viability of PBMCs was modestly affected by exposure to PCB (p,p′-DDT and β-HCH), while exposure to Arochlor-1254 at 10−9 to 10−11 M for 24 hours increased the viability of PBMCs. Prolonged exposure to DDT for 7 days significantly decreased the viability of Jurkat T-cells (Supplementary Fig. 1). After 6 hours of treatment with the three POPs, the basal platelet OCR levels significantly decreased only at a concentration of 100 nM of Arochlor-1254 and p,p′-DDT. After 24 hours, the basal platelet OCR levels showed a non-linear increasing trend. Maximal platelet OCR levels did not show clear patterns. For PBMCs, the basal OCR levels were higher after 24 hours of treatment with β-HCH and p,p′-DDT than in the control group, with a non-linear trend. However, this trend disappeared after 72 hours of treatment. Treatment of PBMCs with Arochlor-1254 did not show any clear effect when compared with the control group. There was little effect of the three compounds on maximal OCR levels.
In contrast, apparent detrimental effects of POPs on OCR levels in Jurkat T-cells could be observed on day 7. The basal and maximal OCR levels at day 7 were significantly lower after treatment with Arochlor-1254 and β-HCH, than in the control group. Low basal OCR levels were observed after treatment with 100 pM to 100 nM of these compounds; however, there was no linear dose-response relationship. The effect of p,p′-DDT on Jurkat T-cell OCR levels at day 7 could be evaluated only with the 0.1 pM concentration as other doses led to cell death. Both basal and maximal Jurkat T-cell OCR levels significantly decreased even with 0.1 pM of p,p′-DDT. In contrast to the day 7 results, at day 2 the basal and maximal Jurkat T-cell OCR levels were higher after treatment with POPs than in the control group, especially for β-HCH.
DISCUSSION
This study demonstrated interesting relationships between chronic POP exposure and mitochondrial function in both humans and in vitro cells, even though the patterns differed by cell and POP types. The mean PBMC OCR levels strongly decreased as the serum PCB concentration increased, and this relationship was similarly observed in all subgroups stratified by age, sex, BMI, T2DM, and hypertension. However, in platelets, serum OCP concentrations were inversely associated with mean basal platelet OCR levels. This association was more pronounced with old age, in men, non-obese subjects, and subjects without T2DM.
One possible explanation for the complex results is that the biological effects of OCPs and PCBs may differ depending on the cell type. As there was no correlation of basal OCR levels between platelets and PBMCs, mitochondrial function may vary, depending on cell type, in the same individual. The in vitro experiment results showed similar complexity. The effect of POPs on OCR levels differed depending on cell type, chemical compound, dose, and treatment duration. Furthermore, non-linear dose-responses were observed in the low-dose range of chemicals. For example, certain low-dose ranges of Arochlor-1254, β-HCH, and p,p′-DDT even increased platelet and PBMC OCR levels at 24-hour, and of Jurkat T-cells at day 2.
Among the cell types we used, only Jurkat T-cells could be used to evaluate the effect of POPs on OCR levels at day 7 of treatment. Because all biological effects of low-dose POPs in humans reflect chronic effects, it is reasonable to assume that Jurkat T-cell results at day 7 would be the most relevant results to humans. Among the three compounds, Arochlor-1254 and β-HCH at a concentration range of 100 pM to 100 nM significantly decreased the basal and maximal OCR levels at day 7 non-linearly. For p,p′-DDT, the extremely low-dose of 0.1 pM decreased the mean Jurkat T-cell OCR levels at day 7. In the human study, p,p′-DDT did not show any significant results in the analyses focusing on individual compounds. However, it is important to note that any epidemiological findings on POPs cannot be interpreted from the viewpoint of individual compounds, which were directly measured and demonstrated statistical significance because humans are exposed to a mixture of various lipophilic chemicals [18]. Therefore, the direct comparison of individual compounds between the in vitro and human studies is objectionable.
Several pharmaceutical drugs have long been recognized as mitochondrial toxins [20]. Moreover, there is emerging evidence that common environmental chemicals can also induce mitochondrial dysfunction [14]. Various environmental chemicals can act as mitochondrial toxins at low doses through diverse mechanisms such as effects on mitochondrial fusion and fission, structural damage, and oxidative phosphorylation dysfunction [14,21,22]. However, certain low doses of chemical exposure can improve mitochondrial function through multiple stress responses, particularly when the exposure is transient [23,24]. In fact, findings from the present in vitro study corroborate this hormetic effect of POP compounds. As the exposure pattern of low-dose environmental chemicals in humans is typically chronic, mimicking chronic exposure is crucial to evaluate the effects of environmental chemicals in in vitro or animal experimental studies with certainty.
Among various environmental chemicals, chemical mixtures such as POPs and heavy metals are of utmost concern for humans because of their long half-lives [25]. Importantly, these chemicals tend to accumulate in the mitochondria because of the lipophilicity of POPs and the cationic property of heavy metals [14]. It is well-known that covalently linking drugs with a lipophilic cation such as an alkyl triphenylphosphonium moiety can increase drug concentration in mitochondria by 100 to 1,000 times than that in the medium [26].
Generally, aging [1], cigarette smoking [27], obesity [28,29], diabetes [3,28], and hypertension [29] are known to be pathophysiologically associated with mitochondrial dysfunction. However, in the current study, the associations of general or clinical characteristics with basal OCR levels of platelets and PBMCs were not clear. Although there were low basal platelet OCR levels in subjects with hypertension and low basal PBMC OCR levels in diabetic subjects, other characteristics were not notably associated with the basal platelet or PBMC OCR levels.
Inconsistent findings in humans might be related to the dynamic nature of mitochondrial functions. Although mitochondrial function varies among individuals and both genetic and environmental factors contribute to their variability, the mitochondrial function of an individual is also flexible and adaptable to meet the energy demands of a particular cell type under quiescent conditions and during biological activation [30]. For example, OCR levels in peripheral blood cells among patients with certain mitochondrial dysfunction-related diseases, such as sickle cell anemia, autism, acute pancreatitis, and septic shock, were compared with those of control groups in small-scale clinical studies [8–11,13]. The results demonstrated that OCR levels of patient groups were not consistently lower than those of control groups. Increased oxidative stress in patients likely adds to energy demand, resulting in adaptive changes in mitochondrial dynamics/composition to meet ATP demand over time.
We observed higher basal OCR levels in PBMCs in elderly subjects than in younger subjects. It was an unexpected finding because it is generally well-known that mitochondrial function is impaired in numerous cell types with aging [31]. However, previous epidemiological studies using OCR levels of peripheral blood cells have reported inconsistent associations between age and mitochondrial function. For example, one study reported reduced reserve capacity in monocytes among older adults compared to younger adults [32]. However, age was not associated with the mitochondrial function of platelets or lymphocytes in other studies [33,34]. Unlike peripheral blood cells, aging has been consistently associated with a reduction in mitochondrial respiration in skeletal muscle [35–37].
Recent advances in the measurement of cell bioenergetics in peripheral blood allow the determination of parameters that affect metabolism in intact cells, allocation of oxygen consumption to ATP synthesis, and overall oxidative phosphorylation capacity. Some researchers suggest that monitoring the bioenergetic health state of immune cells and platelets from blood can be a potentially useful diagnostic tool for evaluating an individual’s overall bioenergetic health and in epidemiological studies [19]. However, our study demonstrated no correlation of basal OCR levels between platelets and PBMCs, suggesting the possibility of different OCR pattern levels for different cell types. In fact, it is practically difficult to measure OCR levels in diverse cell types in epidemiological studies. If OCR levels are measured in only one cell type in peripheral blood, they may not have utility as a surrogate marker of general mitochondrial function in humans.
This study has several limitations. First, it was cross-sectional, which does not allow for the determination of a temporal correlation. Nonetheless, the possibility that mitochondrial function could affect serum PCB and OCP concentrations is unlikely and epidemiological findings were supported by in vitro experiments. Second, although we measured the basal OCR levels in both platelets and PBMCs, the value of the summation of activity in a population of PBMCs rather than OCR levels in discrete subsets of PBMCs can be problematic because the pattern of OCR levels in peripheral blood cells differed depending on the PBMC type [13].
Despite these limitations, this is the first human study that evaluated the association of chronic exposure to low-dose POPs with mitochondrial function, measured as OCR. In addition, the findings from the in vitro study corroborated the findings from the human studies. Overall, these studies suggest the possibility that chronic exposure to low-dose POPs can be harmful to humans, specifically by impairing oxidative phosphorylation in the mitochondria. Mitochondrial dysfunction may be the primary mechanism that can explain recent findings linking low-dose POPs and various chronic diseases in humans.
 XML Download
XML Download