

 PDF
PDF Citation
Citation Print
Print



INFLAMMATION AND DIABETES
Free fatty acids and pro-inflammatory cytokines from the adipose tissue have a negative effect on the intracellular insulin signaling pathway. Free fatty acids activate atypical protein kinase C (PKC), IκB kinase (IKK-β), c-jun N-terminal kinase, leading to the serine/threonine phosphorylation of insulin receptor substrate (IRS) 1 and the inhibition of its signaling.1) As a result, free fatty acids contribute to the insulin resistance in target tissues. Free fatty acids can also promote the secretion of pro-inflammatory cytokines by binding to Toll-like receptors.
The very first experimental evidence connecting inflammation to insulin resistance was the report showing that the overexpression of tumor necrosis factor alpha (TNFα) induced insulin resistance.2),3) Since that original report, numerous proteins secreted by adipose tissues were found to be associated with insulin resistance, including monocyte chemoattractant protein-1 (MCP-1), plasminogen activator inhibitor-1, interleukin (IL)-6, resistin, leptin, visfatin, angiotensinogen, retinol binding protein 4 (RBP4), serum amyloid A, and many others. TNFα secreted by the adipose tissue promotes the lipolysis of adipose tissue and inhibits insulin signaling. Furthermore, the neutralization of TNFα reverses insulin resistance in mouse models; however, this phenomenon has not been observed in humans. IL-6 increases the expression of suppressor of cytokine signaling 3 (SOCS-3) and inhibits IRS signaling through the induction of SOCS-3.4)
TNFα receptors, IL-1 receptors, Toll-like receptors, and advanced glycation end-product receptors mediate the intracellular inflammatory process by stimulating IKK-β/nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) and JNK signaling. Furthermore, reactive oxygen species, endoplasmic reticulum (ER) stress, ceramide, and PKC isoforms have been implicated in the regulation of intracellular inflammation.1)
In addition to insulin target tissues, pancreatic islet inflammation could be detrimental to the pancreatic function, resulting in diabetes progression and insulin secretory dysfunction. In the pancreatic islets of type 2 diabetes patients, inflammatory cell infiltration, amyloid deposition, and the expression of high cytokine levels can be detected, eventually leading to the development of fibrosis. This type of pancreatic inflammation is commonly observed as a result of obesity, with cytokines, hyperglycemia, and dyslipidemia further contributing to this process. Moreover, high expression levels of IL-1β promote insulin secretory dysfunction and beta-cell apoptosis; however, in obese type 2 diabetes patients, IL-1β antagonists have been shown to decrease islet inflammation, as well as blood glucose levels.5) This increased IL-1β expression has also been observed in a gene array study of human islets from type 2 diabetes patients.6)
INFLAMMATION AND FATTY LIVER
Recently, the prevalence of non-alcoholic fatty liver disease (NAFLD) has been increasing and it is commonly detected in obese patients. Inflammation also plays an important role in the progression of NAFLD; furthermore, the expression of intra-hepatic inflammatory markers increases proportionally relative to the degree of obesity. Since inflammation can be induced by increased lipid deposits in the adipose tissue, similar processes occur in the liver tissue. In obese individuals, free fatty acids and pro-inflammatory cytokines enter through the portal vein, promoting inflammation in the liver and inducing selective hepatic insulin resistance. In the liver, these cytokines are known to activate Kupffer cells, cells responsible for the induction of the inflammatory processes. In addition to Kupffer cells, other immune cells, such as natural killer cells, B cells, T cells, dendritic cells, and hepatic stellate cells, can also mediate inflammation, resulting in the progression of NAFLD to steatohepatitis and liver cirrhosis.
Several studies described the connection between inflammatory cytokines and the development of NAFLD. For example, adipocyte-specific fatty acid-binding protein (A-FABP) is expressed in adipocytes and is associated with insulin resistance, metabolic syndrome, and type 2 diabetes. It has been reported that the levels of A-FABP are positively associated with the NAFLD stage, the levels of TNFα, insulin resistance index, and metabolic syndrome.7) In another study, Park et al.8) investigated whether insulin resistance and systemic inflammatory markers could be used as independent predictors of NAFLD in non-obese men. In this cross-sectional study, multivariate analysis indicated that an increase in C-reactive protein (CRP) per 1 standard deviation (SD; 1.48 mg/L) and homeostatic model assessment-insulin resistance per 1 SD (0.63) were independent risk factors for NAFLD.8)
We investigated the connection between silent mating type information regulation 2 homolog (sirtuin, SIRT), NAFLD, and insulin resistance.9) The SIRT family consists of nicotinamide adenine dinucleotide-dependent enzymes that modify proteins via de-acetylation. For example, SIRT1 promotes fatty acid oxidation in the liver by activating adenosine monophosphateactivated protein kinase; however, the connection between exendin-4 and SIRT1 expression has not been previously reported. Therefore, we used high-fat diet-induced obesity mouse model (C57BL/6J), as well as cell culture models, to investigate whether the protective effects of exendin-4 on fatty liver involved SIRT. We observed that exendin-4 increased SIRT1 expression in mice with high-fat diet-induced NAFLD by stimulating fatty acid oxidation.10) Subsequently, we also showed that exendin-4 treatment elevated SIRT1 expression, leading to the reduction of ER stress parameters in hepatocytes11)
ADIPOSE TISSUE AND INFLAMMATION
The adipose tissue secrets numerous factors that are involved in metabolism, inflammation, and immunity. Inflammatory mediators produced by the adipose tissue are known to contribute to the pathogenesis of diabetes and cardiovascular diseases. For example, serum levels of inflammatory cytokines are different between healthy and obese subjects, suggesting the indirect role of these mediators. Blood levels of pro-inflammatory cytokines are also associated with strokes and cardiovascular mortality.12) For example, MCP-1 is highly expressed in the adipose tissues of obese subjects; however, the blood levels of MCP-1, IL-6, RBP4, and visfatin are decreased if these obese subjects were to lose their body weight. When morbidly obese subjects lose weight, the reduction of both adipose tissues and adipose tissue macrophages is observed.13) Therefore, the expansion of adipose tissues can induce inflammatory processes in the body and contribute to the progression of metabolic and cardiovascular diseases.
INFLAMMATORY BIOMARKERS AND METABOLIC DISEASES
Clinical studies clearly demonstrate the connection between subclinical inflammation and the development of diabetes and cardiovascular diseases. For example, in the crosssectional and longitudinal studies, the blood levels of high sensitivity CRP (hsCRP), an inflammatory marker, were closely correlated with the metabolic syndrome components.14),15) It was reported that there was significant correlation between hsCRP and the blood glucose levels even in the euglycemic state, with the glucose levels and hsCRP positively correlated in 10,059 healthy Korean individuals.16) In Rancho-Bernardo study, the baseline levels of IL-6 and hsCRP were significantly correlated with the severity of carotid intima-media thickness,17) while baseline blood TNFα levels were associated with the later progression of glycemic status and NAFLD.18),19) Furthermore, weight gain for several years was correlated with increased inflammation, and the elevated levels of inflammatory markers, such as haptoglobin, ceruloplasmin, alpha1-antitrypsin, orosomucoid, and fibrinogen, at baseline could be used as the predictors of future weight gain.20)
We also investigated the impact of inflammation on the correlation between insulin resistance and mortality in 165,849 apparently healthy Korean adults in a longitudinal study. During the follow-up period of 1,417,325.6 person-years, there was a correlation between the levels of hsCRP and all-cause and cancer-related mortality risks, irrespective of insulin resistance status. This correlation was more evident in the case of cancer-related mortality.21) The Da Qing Diabetes Study showed that there was a significant association between the baseline hsCRP levels and 10-year all-cause mortality.22) Furthermore, the meta-analysis demonstrated that elevated hsCRP levels could predict the risk of all-cause and cardiovascular mortality in the general population.23)
INFLAMMATION AS A TARGET OF METABOLIC DISEASES
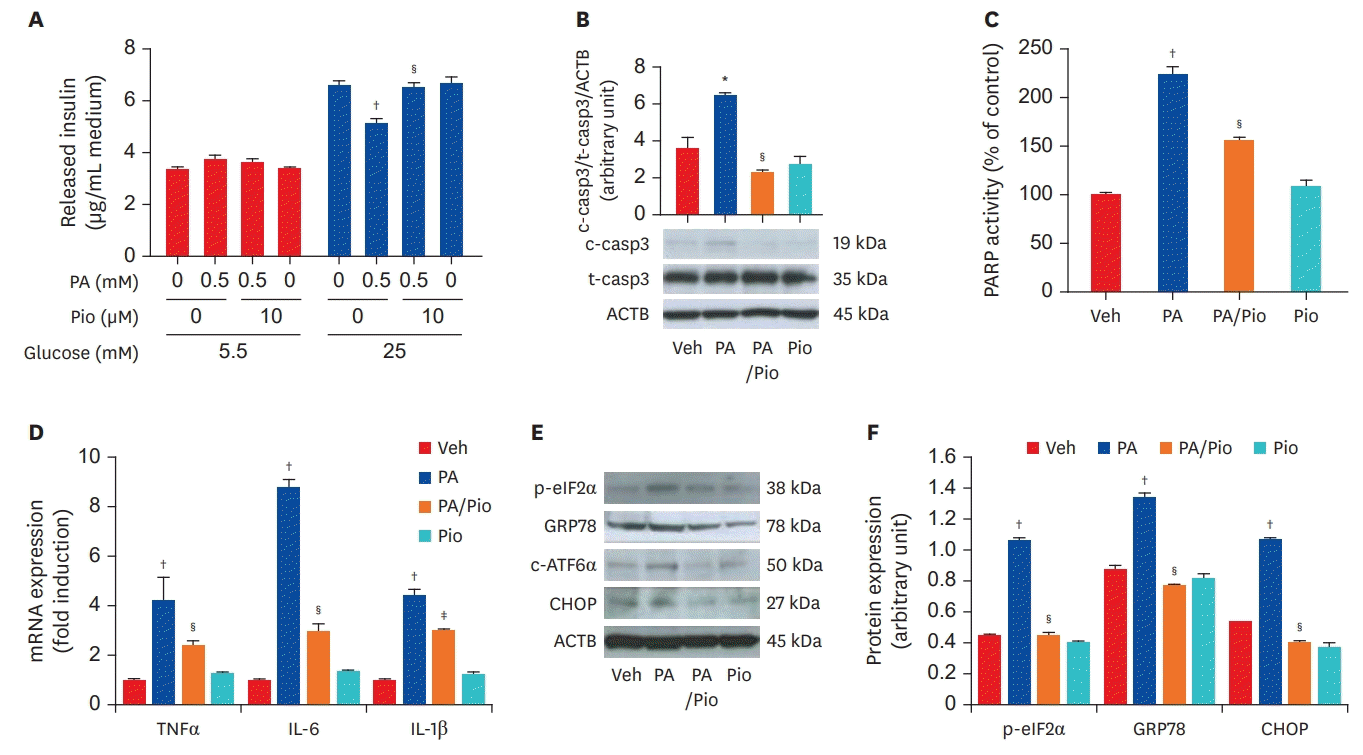
Thiazolidinediones, anti-diabetic drugs, has anti-inflammatory effects in addition to its anti-hyperglycemic properties. It can inhibit NF-κB activity and reduce inflammatory signaling. Furthermore, peroxisome proliferator-activated receptor alpha (PPARα), PPARδ, liver X receptor are nuclear receptors that also have anti-inflammatory effects. We previously reported that pioglitazone, a PPARγ agonist, significantly reduced inflammation and ER stress in a mouse insulinoma cell line (Figure 1).24) The anti-inflammatory effect of pioglitazone was also confirmed in vivo using pancreatic islets of high-fat-fed KK-Ay mice. In these experiments, pioglitazone prevented β-cell apoptosis by reducing ER stress and inflammation.24) Statins also inhibit the activation of NF-κB, activator protein-1, hypoxiainducible factor 1-alpha, and reduce the expression of cytokines and CRP. In 1876, Ebstein25) reported that high dose salicylate had glucose-lowering effects; these glucose-reducing effects of salicylate were due to the inhibition of NF-κB signaling. High doses of aspirin (up to 7 g/day) have been shown to lower the blood levels of free fatty acids, triglycerides, glucose, and induce numerous metabolic effects, such as the inhibition of hepatic glucose production and the stimulation of insulin-dependent glucose transport. However, high doses of aspirin also increase bleeding, limiting its use. In randomized controlled trials, salsalate, a non-acetylated salicylate, was reported to have glucose-lowering effect, although renal safety issues may limit its clinical application.26) Additional evidence is needed to further assess the effect of salsalate on metabolic diseases.
Lifestyle intervention plays a very important role in the reduction of low-grade inflammation. The Action for Health in Diabetes trial, which examined the effect of intensive lifestyle intervention (ILI) on cardiovascular mortality in overweight/obese type 2 diabetic subjects, found that CRP was significantly reduced in the ILI group.27)
CONCLUSION
Inflammation and insulin resistance observed in obesity lead to metabolic diseases. Current research is focused on targeting inflammation, since the reduction of inflammation would improve metabolic diseases. Basic lifestyle interventions, such as regular exercise and healthy eating, are very important even in the absence of weight reduction and should be regarded as essential treatment methods.
 XML Download
XML Download