

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Taurine, a nonessential amino acid, is a frequently used ingredient in energy drinks, as it improves athletic and mental performance. This amino acid is also an essential regulator of physiological processes and pathologies, such as metabolism and cancer development; this aspect was either unknown or ignored until the last decade, when it attracted the attention of scientists [1-3].
Taurine was discovered in Germany in 1827 by Tiedemann and Gmelin [4], who isolated it from ox (Bos taurus) bile. Ten years later, Demarcay [5] named this amino acid taurine, and 20 years later, Jacobsen and Smith [6] found that its structure contains sulfur. Since then, the natural occurrence of taurine has been recognized in a wide variety of invertebrate and vertebrate tissues [6] in some plants, algae, and fungi [7]. In mammals, except felines and foxes, which cannot synthesize taurine, the total taurine pool is balanced between synthesis and absorption from food. In humans, taurine plays a functional role in vital organs, such as the brain, eyes, kidneys, and heart. It performs several primary physiological functions, including osmotic regulation, and has antioxidant, antiapoptotic, and anti-inflammatory effects [1]. The mechanisms through which taurine exerts these effects are not completely known, but are currently being unveiled. This review aims to provide an overview of taurine–bone interactions based on discoveries in the last decade. We summarize how this amino acid affects bone development and functions, as well as orthopedic disorders, including bone cancer.
Go to : 
PROPERTIES OF TAURINE
Taurine (2-aminoethane-1-sulfonic acid) is a sulfur-containing amino acid with a β-amino group and an acidic sulfonic group (R-SO3H) separated by two methylene (CH2) moieties. Unlike several other amino acids, taurine does not contain a carboxyl group; therefore, it is considered a nonproteinogenic amino acid. Taurine´s molecular formula is C2H7NO3S; its exact molecular mass is 125.014664 g/mol; its melting point is 300°C; and its solubility in water is 80.7 mg/mL. Although taurine has an isoelectric point of 5.15, it is considered a zwitterion at physiological pH, with a pKa value of 1.05 and pKb value of 9. Taurine is commercially manufactured from ethylene oxide or monoethanolamine [8].
Go to :
BIOSYNTHESIS OF TAURINE
The primary source of taurine for humans is the diet, especially seafood and meat [9]. Human milk or formula compensates for insufficient synthesis in infants and children, which develops until adulthood. Endogenous synthesis in adults occurs in discrete amounts, ranging from 0.4 to 1.0 mmol per day, depending on protein intake and hepatic function [10]. De novo production of taurine has been mainly found in the liver and brain; however, other tissues, including the kidney, skeletal muscle, white and brown adipose tissues, pineal gland, and retina, can also synthesize it [6]. There are three putative pathways for taurine production from cysteine. The first is the cysteine sulfinate pathway (I), which includes two reactions: decarboxylation of cysteine sulfinate to hypotaurine by cysteine sulfinate decarboxylase, and oxidation of hypotaurine to taurine by hypotaurine dehydrogenase [11]. In pathway (II), cysteine sulfinate is converted to cysteate and taurine by cysteine dioxygenase and cysteate decarboxylase, respectively. The third pathway (III) uses cysteamine produced by the degradation of coenzyme A, and finally, cysteamine is transformed into hypotaurine by 2-aminoethanethiol dioxygenase (Fig. 1) [12,13].
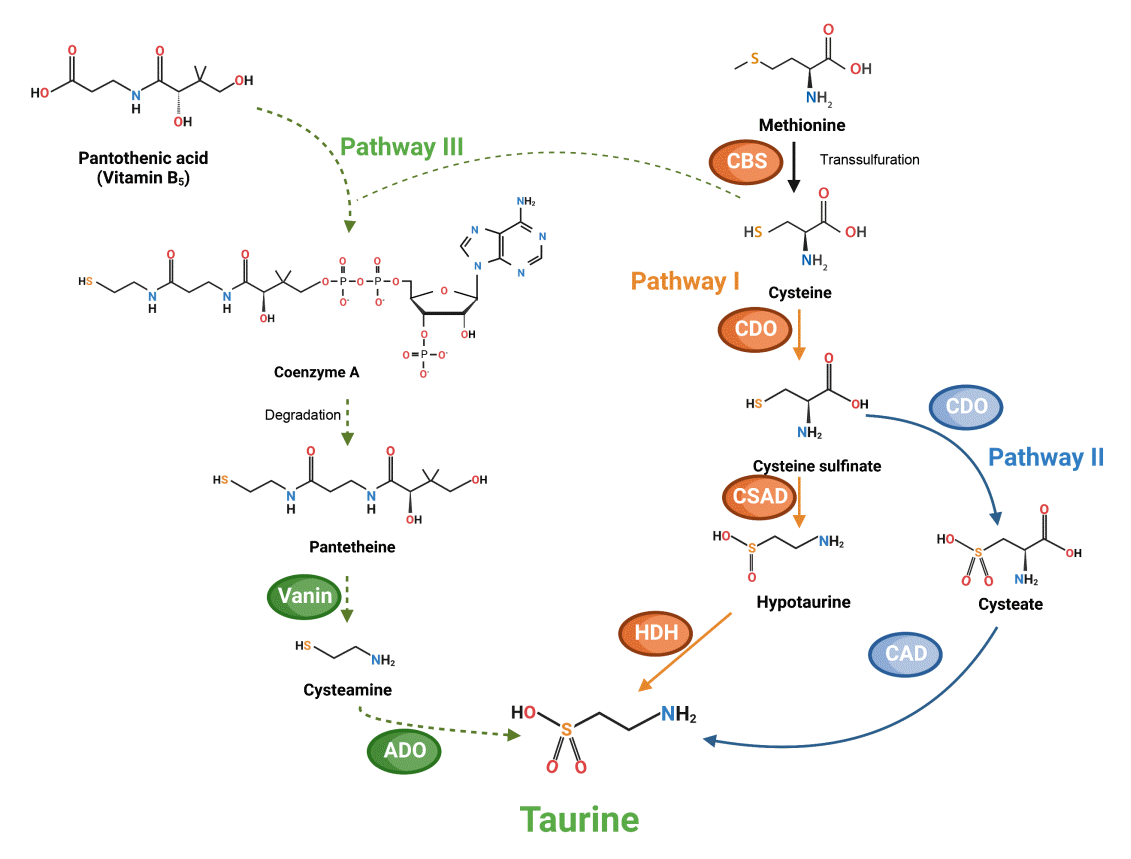 | Fig. 1.Biosynthetic pathways of taurine in humans. Orange and blue arrows indicate synthesis pathways (I) and (II) from cysteine, respectively. Synthesis pathway (III) is indicated in green arrows [13] (created with BioRender.com). CBS, cystathionine β-synthase; CDO, cysteine dioxygenase; CSAD, cysteine sulfinate decarboxylase; HDH, hypotaurine dehydrogenase; CAD, candysteic acid decarboxylase; ADO, 2-aminoethanethiol dioxygenase.
|
Go to :
ABSORPTION, DISTRIBUTION, AND EXCRETION OF TAURINE
The taurine supply in the human body is covered by intestinal uptake and biosynthesis, mainly in the liver. After oral ingestion, intestinal transport moves taurine by two carriers: protoncoupled amino acid transporter 1 (PAT1) and sodium- and chloride-dependent taurine transporter (TAUT). PAT1 has pH-dependent activity on the millimolar order and functions as the primary transporter at higher taurine concentrations; this protein also transports glycine, alanine, and proline [14]. TAUT is a beta-alanine transporter; unlike PAT1, its taurine affinity is in the micromolar range, but with low transport capacity [15]. In addition to endogenous synthesis, taurine content in the liver (approximately 10 mM) is maintained by portal vein transport and taurine conjugated with bile acids, which are products of cholesterol catabolism. Taurine metabolism involves the formation of 5-glutamyl-taurine and taurocholate, and its excretion occurs in the kidney through urine or by the enterohepatic cycle through feces conjugated to bile acids (Fig. 2) [13,14,16].
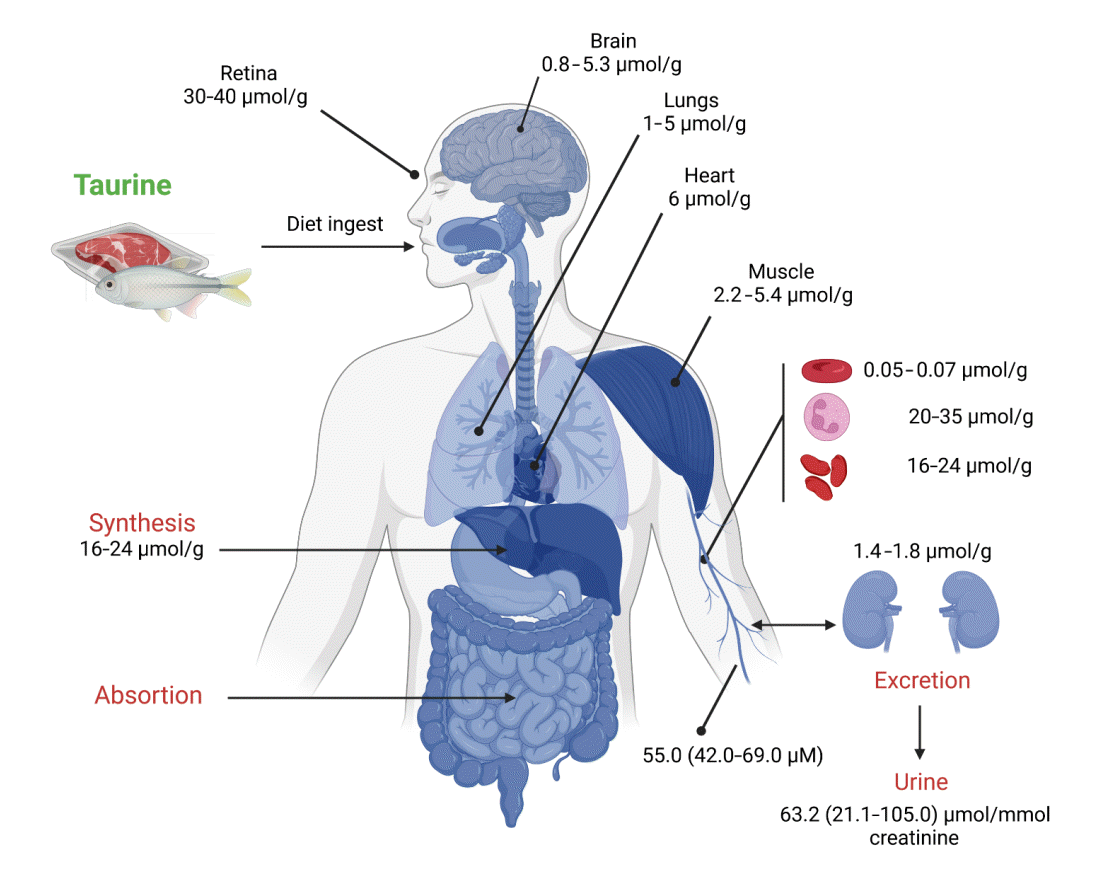 | Fig. 2.Human taurine tissue content, distribution, and excretion. The plasma level of taurine is the sum of the dietary intake and the endogenous synthesis by the liver, while the kidney is the primary organ for its excretion. Tissues with remarkably high taurine accumulation include the retina, heart, lung, and skeletal muscle, as well as some hematopoietic cells such as erythrocytes, platelets, and white blood cells. Data represents the amount per day. Data taken from [13] (created with BioRender.com).
|
Go to :
CELLULAR FUNCTIONS OF TAURINE
Antioxidant activity
Taurine is frequently classified as an antioxidant; it scavenges reactive oxygen species (ROS), reduces ROS generation, and alleviates the harmful effects of oxidative stress. Free radical scavenging by taurine depends on its concentration; it is effective against peroxyl radicals (ROO•), superoxide anion (O2•−), nitric oxide (NO•), and peroxynitrite (ONOO−), but it does not scavenge H2O2 [17]. However, the main antioxidant property of taurine is attributed to its mitochondrial protective effect, which blocks ROS generation [18]. Taurine reacts with uridines of mitochondrial tRNAs to form 5-taurinomethyluridine (τm5U) and 5-taurinomethyl-2-thiouridine (τm5s2U), thereby improving the codon-anticodon interaction between the uridine-uridine-guanosine (UUG) codon and taurine-modified adenosine-adenosineuridine, optimizing the translation of encoded proteins that are rich in UUG regions of subunits 5 (mt-ND5) and 6 (mt-ND6) of the respiratory chain complex I [19]. Another antioxidant mechanism of taurine involves normalizing the enzymatic machinery against oxidative stress in the cell. Many studies have described the restoration of levels of catalase, glutathione peroxidase, glutathione, superoxide dismutase (SOD), and thioredoxin reductase by taurine treatment after exposure to oxidative inducers [1,20], with the most recent studies focusing on diabetes-induced oxidative stress [21]. In contrast, the generation of taurochloramine (TauCl) and taurobromide (TauBr) by the reaction of taurine hypochlorous and hypobromous acids derived from the myeloperoxidase system in activated neutrophils is an indirect antioxidant mechanism of this amino acid [22]. Taurine haloamines significantly induce antioxidant enzyme expression, promoting the nuclear translocation of nuclear factor E2-related factor 1 (Nrf2) (Fig. 3) [23]. The following sections describe additional properties of these taurine analogs.
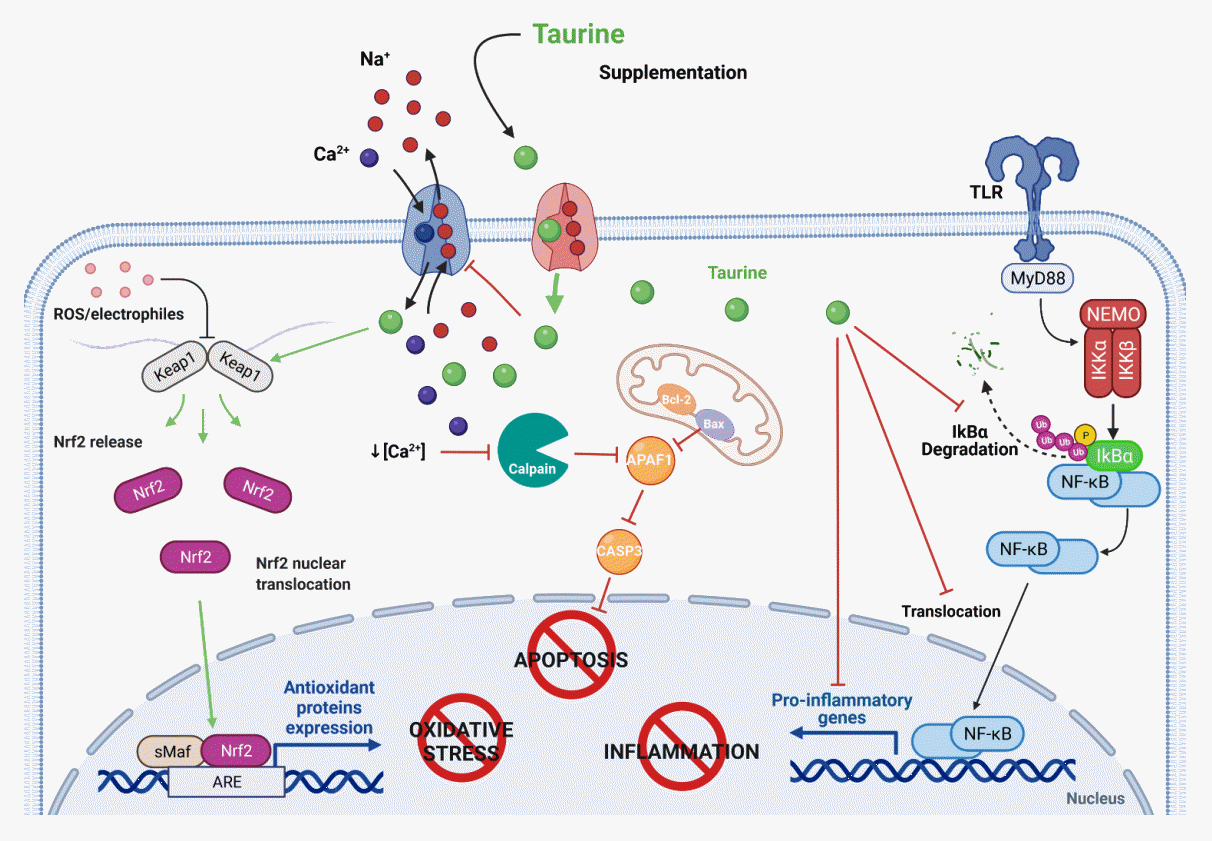 | Fig. 3.Cytoprotective roles of taurine against cell damage. Antioxidant activity: taurine facilitates the release of the nuclear factor E2-related factor (Nrf2), a redox-sensitive transcription factor, by the Kelch-like ECH-associated protein 1 (Keap1), and the translocation of Nrf2 to the nucleus and its binding to the antioxidant response element (ARE) activates the gene transcription of antioxidant enzymes. Anti-apoptotic activity: taurine’s contribution to calcium homeostasis blocks the activation of the calpain-dependent apoptotic cascade. Anti-inflammatory activity: taurine represses the degradation of the inhibitor of nuclear factor-κB (NF-κB) alpha (IκBα) and keeps NF-κB inactivated; it also prevents its translocation to the nucleus and reduces the transcription of genes encoding proinflammatory cytokines (created with BioRender.com). APAF1, apoptosis protease-activating factor-1; Bax, Bcl-2-associated X protein; BCl-2, B-cell lymphoma 2; CASP3, caspase-3; Ikβα, nuclear factor of kappa light polypeptide gene enhancer in B-cells inhibitor, alpha; Keap1, Kelch-like ECH-associated protein 1; MyD88, innate immune signal transduction adaptor; Nrf2, NEMO, NF-kappa-B essential modulator; sMaF, musculoaponeurotic fibrosarcoma; TLR, Toll-like receptor.
|
Calcium homeostasis
Taurine can preserve intracellular calcium (Ca2+) concentrations by inhibiting influx from extracellular routes or releasing it from intracellular pools. One of the known mechanisms underlying this effect is the extrusion of taurine and sodium (Na+) from the cell via taurine/Na+ symport, which limits Ca2+ overload through Na+/Ca2+ exchange in ischemia-reperfusion injuries [24]. This effect suggests the therapeutic use of taurine in surgery for cardiac damage, heart transplantation, and myocardial infarction [25]. Taurine also regulates the activity of Ca2+-ATPase in the sarcoplasmic reticulum, ultimately maintaining Ca2+ concentrations, which has implications for diastolic dysfunction of the heart or impaired fibers type I and II of the skeletal muscle [26,27]. The role of taurine in calcium-sensitive excitation-contraction in muscular pathologies has been used to test the hypothesis that increasing taurine intake can improve skeletal muscle function in a mouse model of Duchenne muscular dystrophy [28]. Taurine also modulates the calcium influx produced by coupling glutamate to N-methyl-d-aspartate receptors, preventing membrane depolarization by opening chloride channels or inhibiting L-, P/Q-, and N-type voltage-gated Ca2+ channels [29].
Cell death
Closely related to its antioxidant activity, taurine maintains normal cellular function and regulates apoptosis (Fig. 3). The interruption of the respiratory chain by a deficiency in complex I has been demonstrated in taurine-deficient models. In fact, mitochondrial damage leads to increased membrane permeability, inducing cytochrome c release, apoptosome formation, and activation of caspase-9 and caspase-3 [30]. Taurine also inhibits apoptosis induced by toxic agents, such as tert-butyl hydroperoxide [31], acrylamide [32], and arsenic [33], in cell cultures that activate the phosphatidylinositol 3-kinase pathway/Akt and inhibit nuclear translocation of nuclear factor-kappa B (NF-κB). Interestingly, taurine supplementation protects osteocyte-like cells from cell death triggered by ROS in vitro [34]. This non-classical amino acid is also involved in neuronal cell death protection against elevated circulating corticosteroids due to chronic stress, by positively regulating extracellular signal-regulated kinase and Nrf2 [35]. Therefore, taurine has been suggested as a potential neuroprotective agent, particularly for glaucoma treatment [36]. More recently, taurine has been demonstrated to limit arsenic-induced cell death by downregulating pyroptosis and autophagy in vitro and in vivo, thereby reducing hepatotoxicity [37]. In addition, taurine has been proposed as a potential therapeutic agent for treating alcohol-induced cell death and nonalcoholic fatty liver disease resulting from obesity [38,39].
Energy metabolism
Although taurine biosynthesis is limited in humans, it also plays a vital role in energy metabolism. Under taurine deficiency, skeletal and heart muscles, liver, and adipose tissue show metabolic and physical dysfunction, highlighting the downregulation of genes involved in fatty acid metabolism [40]. Taurine supplementation in the diet can also strengthen energy metabolism for muscle performance, cardiac function, and liver activity [25,41,42]. For example, in obesity models, taurine treatment lowers fatty acid oxidation, decreases weight, and improves hyperglycemia and hyperinsulinemia. Details of these effects and their molecular mechanisms have recently been compiled elsewhere [40,43].
Inflammation
Although the antioxidant properties of the taurine derivatives, TauCl and TauBr, have already been noted, their anti-inflammatory properties are remarkable. TauCl inhibits the overproduction of inflammatory mediators, such as tumor necrosis factor-α, interleukin-1, -2, -6, -8, 10, and -12, monocyte chemo-attractant protein-1 and -2, cyclooxygenase (COX)-2, and prostaglandin E2 (PGE2) [44]. The regulation of all these molecules converges on the inactivation of IkappaB kinase, which phosphorylates the inhibitor kappa B-alpha, thereby preventing its degradation and promoting its union with NF-κB involved in the synthesis of proinflammatory mediators (Fig. 3) [45]. The beneficial effects of taurine have recently been explained by the brain injury and inflammation mechanisms in patients with type 2 diabetes [21,46]. Furthermore, new insights have been gained into how the microbiome regulates taurine and healthy aging. Ahmadi et al. [47] reported that a human-origin probiotic cocktail from a healthy infant gut prevented high-fat diet-induced inflammation in elderly mice in a manner dependent on increased taurine abundance in the gut, favoring healthy aging. Additionally, topically applied taurine can regulate skin inflammation caused by ultraviolet light exposure, enabling this amino acid to be suitable for the amelioration of dermatitis and other skin pathologies [48]. The anti-inflammatory role of taurine has been addressed in other conditions, such as adult mouse lungs exposed to cigarette smoke [49], carbon tetrachloride-induced testicular and kidney damage [50], and ulcerative colitis-colorectal cancer [51]. As the inflammatory effect of taurine has become markedly relevant, it is now being used as a biopharmaceutical coupled to collagen sponges to improve wound healing and tissue proliferation [52]. Furthermore, evidence from clinical trials has shown that taurine can significantly decrease the levels of protein C reactive and malondialdehyde 56 days after its supplementation, thereby exhibiting beneficial effects on inflammatory and oxidative stress [53].
Osmotic regulation
Osmoregulation in cells involves the maintenance of salt and water balance, with adjustment of their osmolyte content to minimize injury to the membrane under osmotic disequilibrium. Unlike charged osmolytes, taurine is a neutral zwitterion released or introduced by cells to compensate for deviations from normotonicity, which can occur by different mechanisms [54]. Swelling-induced taurine efflux is energy-independent, temperature-sensitive, and pH-sensitive, and is regulated by non-hydrolytic adenosine triphosphate (ATP) binding via a volume-regulated anion channel [55]. Although taurine efflux is believed to be Ca2+-independent, it was recently demonstrated that it depends on Ca2+ in an insensitive fraction of human retinal Müller cells [56]. Under conditions of hypertonicity, taurine accumulation is mediated by TAUT based on in vivo studies and cultured astrocytes, which has been suggested to be a concerted uptake/biosynthesis interaction; however, this notion has not been well established [57]. Fig. 3 summarizes the functions of taurine in cell protection.
Go to :
TAURINE AND BONE
Bone is a mineralized connective tissue that constitutes the skeletal system and provides mechanical support and protection for organs and bone marrow, as well as conferring strength, toughness, and capacity for continuous growth and remodeling throughout post-fetal life [58]. Osteoblasts, osteoclasts, and osteocytes, the bone cells, are immersed in an organic and inorganic matrix composed of collagen type I, a smaller proportion of non-collagen proteins, and nanosized hydroxyapatite crystals [59].
Two processes are involved in bone growth: intramembranous ossification and endochondral ossification. Intramembranous ossification involves the transition from mesenchymal cells to osteoblasts, mainly by the action of the Runt-related transcription factor 2 (RUNX2) [60], while endochondral ossification involves the formation of a hyaline cartilage template, which is replaced by mineralized bone tissue by the action of SRY-box transcription factor (SOX9) [61]. Once the bone has formed, bone remodeling is described as either the formation of bone by osteoblasts or resorption by osteoclasts.
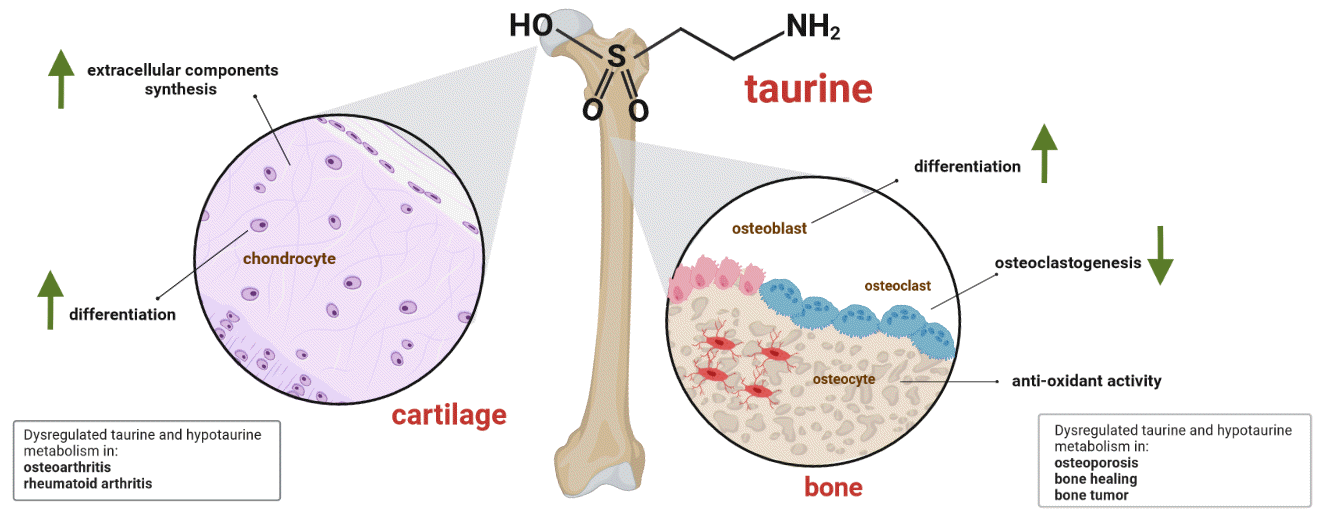
Different amino acids act as precursors of bioactive macromolecules, such as second messengers, cytokines, or neurotransmitters, that control various cellular processes that affect the skeletal system [62]. Although taurine is not a structural component of proteins, it is metabolically involved in many processes that influence bone development [63]. Interestingly, the influence of taurine on bone growth was elucidated less than 10 years ago by Roman-Garcia, who found that vitamin B12 deficiency induced growth retardation and osteoporosis in a mice model parallel with low taurine production by the liver. This effect results in the abrogation of the growth hormone–insulin-like growth factor I axis (GH/IGF-I) and GH resistance. As GH controls taurine synthesis in a vitamin-B12 dependent manner, this amino acid is consequently an upstream regulator of IGF-I production in the liver and a regulator in osteoblasts [64]. Although this study helps to understand the role of taurine in bone development at the molecular level, the impact of taurine at the macro level and on the microstructure has scarcely been explored. Furthermore, few studies have been conducted in the last two decades. At the time of this review, the only study that evaluated the impact of taurine ingestion on human bones revealed significantly higher serum osteocalcin levels in patients with type II diabetes than in controls, suggesting that taurine may modulate bone mineralization [65]. This finding is supported by studies using animal models, which revealed the positive effects of taurine on bone development. For instance, in a mouse model of protein malnutrition, increases in growth plate thickness and improvement in bone volume, trabecular number, connection density, and total porosity were observed after taurine consumption for 12 weeks. Taurine is proposed to exert these effects by elevating IGF-1 levels and promoting the phosphorylation of Janus kinase 2-signal transducer and activator of transcription 5, as these markers are enhanced in the serum, liver, and tibia-growth plate [66]. Consistent with this study, male rats fed a diet containing 2% taurine for 6 weeks had significantly higher femur bone mineral content (BMC)/weight than control rats, suggesting that taurine may have a beneficial effect on bone even though the markers, alkaline phosphatase (ALP) and osteocalcin were not affected [67]. The third study involved the effect of taurine supplementation in the diet of broiler birds, which, after 42 days, presented notably higher levels of serum ALP and bone-specific ALP than control birds. In an analysis of BMC, birds fed a taurine-supplemented diet had the highest percentage of phosphorous, calcium, and phosphate composition, indicating that taurine supplementation boosted the bone mineral composition of meat-type poultry birds [68].
In contrast to previous studies, the oral administration of taurine to mice for 8 weeks decreased cortical and trabecular bone thickness and reduced the density of secondary osteons [69]. Although this previous study suggests the negative effect of prolonged taurine exposure on the bone tissue microstructure, the lack of sufficient studies prevents the drawing of conclusions. Undoubtedly, the effect of taurine on the typical bone structure must be studied thoroughly, as most analyses have focused on its positive effect on damage caused by osteoporosis; such studies are described in the next section.
Taurine and osteoblasts
Osteoblasts are bone-building cells that synthesize collagen matrix and mineralize bone. The mesenchymal progenitors of osteoblasts are activated by specific transcription factors that operate at a distinct time point during differentiation, defining various stages of the osteoblast lineage. The two master transcription regulators in osteoblast differentiation are SOX9 and RUNX2, the first of which mediates the differentiation of mesenchymal stem cells into chondrogenic progenitors but is not expressed in mature osteoblasts. In contrast, RUNX2 is necessary to differentiate osteoblasts from precursors [70]. The regulation of RUNX2 involves several pathways, such as Hedgehog, Notch, Wnt, bone morphogenetic protein (BMP), and fibroblast growth factor (FGF) signaling [71]. Besides SOX9 and RUNX2, other transcription factors, such as osterix, play essential roles in osteoblastic differentiation.
The relationship between taurine and osteoblasts is being elucidated. To date, two studies have shown that taurine stimulates osteogenesis. In MG-63 human osteoblast-like cells, taurine was recently shown to increase the expression levels of RUNX2 and intermediates of the BMP pathway, such as SMAD-1,5,8. These effects enhance the activity of ALP, which is a marker of osteogenic differentiation (Fig. 4) [72]. In mesenchymal stem cells, taurine promotes osteoblastogenesis, increasing ALP activity and calcium deposition owing to the upregulation of osteopontin, RUNX2, and osterix mRNA [73]. Taurine also exerted cytoprotective effects on osteoblasts under stress, as demonstrated in osteoblast-like UMR-106 cells. Pre-treatment with taurine can evade apoptosis induced by H2O2, an effect attributed to the overexpression of Nrf2 and β-catenin, which reduce oxidative damage [74]. In contrast, taurine was found to reduce apoptosis in MC3T3-E1 cells (osteoblast precursor cells) by inhibiting cytochrome C release and caspase-3 and caspase-9 activation [75].
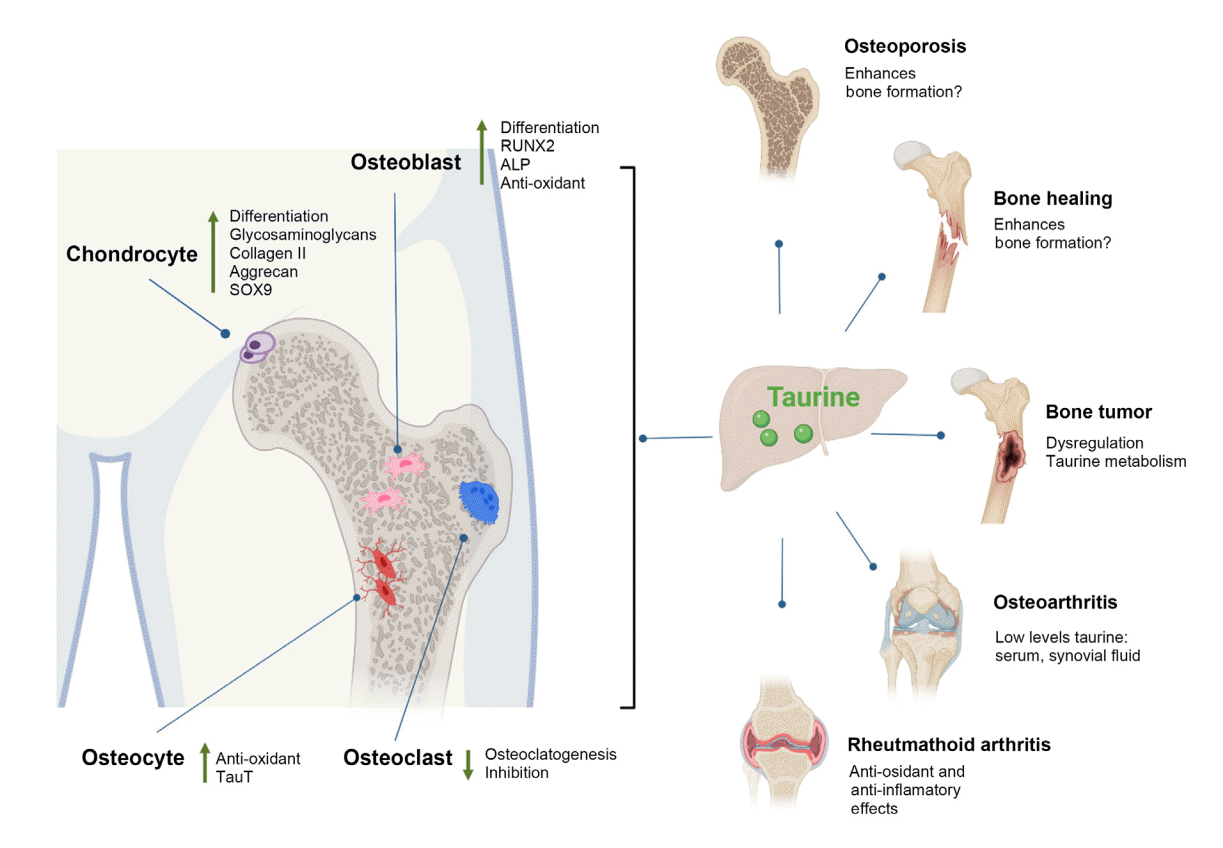 | Fig. 4.Potential actions of taurine in bone and cartilage. Taurine promotes osteogenesis by upregulating Runt-related transcription factor 2 (RUNX2), as well as increasing alkaline phosphatase (ALP) activity and calcium deposition by osteoblasts; it also plays an antioxidant role in osteoblasts and osteocytes. In osteoclastogenesis, taurine exerts an inhibitory function. In chondrogenic differentiation, taurine promotes the synthesis of extracellular matrix components such as glycosaminoglycans and collagen, in addition to increasing the expression of SRY-box transcription factor 9 (SOX9). In bone and cartilage health, taurine promotes the formation of bone calluses in the consolidation process and promotes osseointegration. In pathological conditions such as osteoporosis, taurine supplementation increases bone mass, while in osteoarthritis, rheumatoid arthritis, and some bone tumors there are alterations in the metabolism of taurine (created with BioRender.com). TauT, taurine transporter.
|
The consumption of taurine by preosteoblasts or osteoblasts must play an important role in the differentiation or protective effect; thus, the transport of taurine has attracted attention. In 2006, TAUT mRNA and protein expression was detected in a murine osteoblast cell line (MC3T3-E1) and primary human osteoblasts [76]. Two years later, taurine uptake via TauT was found to be dependent on the presence of extracellular calcium, which was decreased by L-type calcium channel blockers and oxidative agents, such as H2O2, in MC3T3-E1 cells [77]. Although the participation of taurine in osteoblast differentiation and maintenance is evident, more studies are necessary to understand the detailed mechanism by which these events occur.
Taurine and osteoclasts
As cells complementary to osteoblasts, osteoclasts are multinuclear giant cells that can degrade bone by secreting H+, Cl−, cathepsin K, and matrix metalloproteinases (MMPs). Osteoclasts are derived from myeloid lineage cells, mainly in response to receptor activator of NF-κB ligand (RANKL) and macrophage colony-stimulating factor [78]. RANKL is expressed by osteoblasts and osteocytes and regulates osteoclast differentiation, bone resorption, and osteoprotegerin (OPG), which is also secreted by osteoblasts. OPG is a negative regulator of osteoclastogenesis by binding with RANKL and impeding receptor interaction, ultimately playing a vital role in bone homeostasis (Fig. 4) [79]. RANKL-RANK generates a downstream signaling cascade of adaptors/kinases, activating transcription factors that induce the expression of osteoclastogenic markers, such as tartrate-resistant acid phosphatase (TRAP) and cathepsin K [78].
Like osteoblasts, the effect of taurine on osteoclasts has not been explored in detail, and the most remarkable effect is its inhibition of osteoclastogenesis. Several studies in the first decade of the millennium supported this notion; however, crucial details were not obtained until 2010. Taurine was found to inhibit osteoclastogenesis of bone marrow cells co-cultured with osteoblasts without the influence of OPG and RANKL, whereas TAUT small interfering RNA reversed this effect. This finding indicates that the inhibition of osteoclastogenesis by taurine is dependent on its transporter [76]. Similarly, as ROS are crucial in osteoclast differentiation [80], taurine inhibits LPS-induced osteoclastogenesis in RAW264.7 cells, preventing ROS generation by inhibiting COX-2 expression and JNK phosphorylation [81]. A recent study reported evidence supporting the inhibition of osteoclastogenesis by taurine, indicating that TauCl also impeded RANKL-induced osteoclastic differentiation of RAW 264.7, modifying the mRNA of markers, such as TRAP, cathepsin K, and calcitonin receptor [82,83].
Taurine and osteocytes
Osteocytes are the most abundant cells in the bone and are embedded in a lacuna-canalicular network inside the mineralized matrix, which can sense mechanical forces and induce signals to osteoblasts and osteoclasts to initiate remodeling [84]. As mechanosensitive cells, osteocytes are equipped with integrin complexes, such as β1 integrin, and ion channels, such as transient receptor potential vanilloid-type 4 (Trpv4), which serve as their main mechanosensory system for physical stimuli. In addition to assisting with the mechanical adaptation of the bone, osteocytes are a significant source of RANKL, controlling osteoclast differentiation in a positive direction and decreasing the expression level of OPG, or vice versa. Stimulatory factors from osteocytes, such as lipids (PGE2), growth factors (e.g., IGF-1), glycoproteins (e.g., Wnt), free radicals (e.g., NO), and nucleotides (e.g., ATP), have substantial effects on osteoblastogenesis and matrix formation [85].
In contrast to osteoblasts and osteoclasts, the relationship between taurine and osteocytes had not been described until 2 years ago [34], and more findings are yet to be obtained. Interestingly, primary osteocytes and IDGSW3 cells can synthesize taurine as a self-protective mechanism, encoding enzymes that convert cysteine into cysteine sulfinate and hypotaurine [34]. In that study, the researchers also found that taurine supplementation protected IDG-SW3 and MLO-Y4 cells from death triggered by H2O2 treatment (Fig. 4). Additionally, the gene of the taurine uptake transporter, SLC6A6, was highly expressed in IDG-SW3 cells under exogenous taurine supplementation, and sclerostin, a well-known negative regulator of the Wnt signaling pathway, was inhibited by taurine.
Taurine and bone healing
Bone healing is the process of tissue reconstitution in the presence of fractures or bone defects. The major metabolic phases of fracture healing include inflammation, endochondral ossification, and coupled remodeling [86]. The role of taurine and its derivatives in bone healing has been highlighted. The effects of oral administration of taurine on rabbits with osteotomy in the early stages of bone healing included the formation of a callus rich in osseous components compared with that of control rats, which produced a callus rich in fibrous components [87]. In another model of glucocorticoid-induced rabbit osteonecrosis, taurine administration normalized the expression of mitochondrial transcription factor A and the complex responsible for the synthesis of ATP; thus, mitochondrial function prevented the development of osteonecrosis [88]. Additionally, tauroursodeoxycholic acid, a common bile acid, induces new blood vessel formation and enhances bone formation after implantation, as revealed by a three-dimensional micro-computed tomographic analysis of rabbit calvarial defects [89].
Finally, taurine incorporation into biomaterials to improve the osseointegration of bioactive scaffolds for bone defect regeneration has been explored. The three-dimensional combination of polylactic acid, polycaprolactone, gelatin nanofibers, and taurine was found to be suitable for cell growth and proliferation. Furthermore, bone defects treated with this scaffold resulted in higher new bone formation than that observed in the control group, illustrating the potential use of taurine in biomaterials [90].
Go to :
TAURINE AND BONE DISORDERS
Taurine and osteoporosis
Osteoporosis is a chronic condition in which bone mass declines. According to the World Health Organization, adults whose bone mineral density (BMD) or BMC values are at least 2.5 standard deviations less than the mean BMD for normal adults have osteoporosis [91]. Clinically, osteoporosis is characterized by back pain caused by vertebral deterioration, fractures, and weight loss. Osteoporosis-related fractures in the USA are associated with an estimated direct cost of $17 billion per year, with an expected increase to $25.3 billion by 2025 [92].
The relationship between taurine and osteoporosis has been studied using different approaches; most notably, metabolomic studies have shown low taurine levels in the blood of patients with osteoporosis. Previous studies have shown that postmenopausal Brazilian women with osteopenia or osteoporosis had reduced taurine levels in plasma compared to healthy volunteers [93]. The same results were obtained for Caucasian pre-menopausal women in the USA [94], Japanese women with low estradiol and BMD levels [95] and older Chinese adults with low BMD and fracture [96]. In addition to its serum level, taurine levels in urine are lower in menopausal women than in normal women [97]; thus, low taurine levels in the serum and urine highlight the potential of this amino acid as a biomarker of osteoporosis and bone loss. This result may be related to the high plasma homocysteine (Hcy) levels measured in patients with osteoporosis and low BMD, as recently highlighted elsewhere [98]. Increased Hcy levels result in the dysregulation of the transsulfuration pathway, as L-cysteine is synthesized from Hcy and taurine is synthesized from L-cysteine. In osteoporosis, a decrease in taurine synthesis causes the dysregulation of calcium and vitamin D, which are essential elements for bone growth and resorption [99].
The advantage of restoring plasma taurine levels in osteoporosis models has been analyzed; various treatments, including the administration of bone collagen peptides from Bos grunniens [98] alendronate sodium [99] and natural extracts [100], increase the concentrations of serum taurine and normalizes bone turnover markers and biomechanical parameters. Taurine supplementation alleviates osteoporosis by increasing 1,25(OH)2D3, IGF-1, transforming growth factor (TGF)-β, BMC, and BMD levels in ovariectomized rats [101]. It also increases bone mass and improves resistance to force by promoting, serum osteocalcin, and ALP activity in postmenopausal rats [102].
Taurine and bone tumors
Bone tumors are rare neoplasms that encompass a broad group of benign and malignant entities that affect the bone, cartilage, and connective tissues [103]. Sarcomas of bone present an annual incidence of 0.2% in adults and account for 5% of childhood malignancies; as they are benign tumors, they are asymptomatic and are only detected as incidental findings. The three most frequent bone malignancies are osteosarcoma and Ewing’s sarcoma in adolescents, and chondrosarcoma in adults. Although standard treatment includes surgery, chemotherapy, and radiation, surgical resection does not ensure complete removal of the tumor, and recurrence and metastasis can result in a high mortality rate. Therefore, a better understanding of the biology of bone sarcomas is necessary to improve their outcomes, identify and exploit unique characteristics, and propose new treatment targets. Many metabolomic studies have been conducted to search for metabolic biomarkers associated with tumor prediction and diagnosis; however, there are limited data to provide a comprehensive metabolic picture of bone tumors. Nonetheless, taurine has appeared in some of these studies as a possible metabolic marker of bone sarcomas. For example, a serum untargeted metabolomics analysis of 65 patients diagnosed with osteosarcoma revealed higher levels of ascorbic acid, niacinamide, and taurine than in healthy controls [104]. A similar study revealed differences in the synthesis and degradation of ketone bodies and taurine metabolism compared to normal serum [105]. Furthermore, the response of osteosarcoma cells to some treatments highlights taurine as a participant, with the intracellular metabolic profile of MG-63 human OS cells induced by the chemotherapy agent cisplatin and decreased taurine levels, which reflect the activation of defense mechanisms against oxidative stress [106]. Similarly, treatment of Saos-2 osteosarcoma cells with nanographene oxide sheets reduced the levels of several amino acids, such as taurine, which is also related to oxidative stress.
Finally, the action of taurine or its derivatives is important in bone tumor treatment. For example, taurolidine is cytotoxic to osteosarcoma cells and supports the effect of doxorubicin and carboplatin [107]. In addition, taurine chloramine exerts cytotoxic effects against osteosarcoma cell lines (HOS, SAOS-2, and MG-63) by inducing the loss of mitochondrial membrane potential, activating the intrinsic mitochondrial apoptosis pathway [108].
Go to :
TAURINE AND CARTILAGE
The cartilage is a flexible connective tissue in the skeletal system, which maintains fluid joint motion by coating the surfaces between the bones. Cartilage is a semi-rigid, avascular, and aneural tissue, composed of chondrocytes embedded in an extracellular matrix of collagen fibers, proteoglycans, and other noncollagenous proteins. The proliferation and differentiation of marrow-derived mesenchymal stem cells into chondrocytes—also known as chondrogenesis—are complex processes that involve positive regulators, such as FGF, TGF-β/BMP, Wnt/B-catenin, Hedgehog, Notch, and vascular endothelial growth factor pathways. Furthermore, pro-chondrogenic transcription factors, such as SOX9, RUNX, and trico-rhino phalangeal syndrome type 1 (Trps1), also control chondrogenesis [109].
Taurine and chondrocytes
Taurine participates not only in the maintenance of the bone, but also in cartilage maintenance, and could effectively promote chondrogenic differentiation of human umbilical cord-derived mesenchymal stem cells, enhancing the synthesis of glycosaminoglycans and upregulating the expression of collagen II, aggrecan, and SOX9 chondrocyte mRNA [110]. Similar results were found for human articular chondrocytes treated with taurine, such as the promotion of proliferation, maintenance of the phenotype-enhanced accumulation of glycosaminoglycans and collagens, and increased expression level of SOX9 (Fig. 4) [111]. Similarly, osteoblasts express the TAUT for its uptake under certain conditions; chondrocytes also express this carrier, which is upregulated in hypertonic conditions to protect the cell in hyperosmolarity environments [112].
Taurine and osteoarthritis
Cartilage problems include tears and injuries caused by sports accidents, genetic factors, and various types of arthritis. Within the last group, osteoarthritis (OA) is the most common form, defined as a degenerative cartilage disorder that affects diarthrodial joints. The estimated prevalence of OA is 303.1 69 million worldwide among adults aged ≥65 years, representing the primary cause of disability in the elderly population [113].
Metabolism is considerably altered in OA. Normal chondrocytes utilize glucose via glycolysis, and oxidative phosphorylation maintains mitochondrial function under optimal conditions. Normal energy homeostasis is mainly supported by the AMP activated protein kinase–NAD dependent protein deacetylase sirtuin 1– peroxisome proliferator activated receptor γ co‑activator 1α (AMPK–SIRT1–PGC1α) pathway; however, chondrocytes in OA switch from oxidative phosphorylation to glycolysis, impairing mitochondrial metabolism and accumulating ROS, thereby inhibiting AMPK signaling [114]. Furthermore, accumulating evidence shows that other metabolic pathways, such as taurine metabolism, are deregulated in addition to glycolysis. Particularly, metabolomic analysis revealed that in serum samples of OA patients, taurine is significantly lower than that in healthy controls, with a sensitivity and specificity ≥80% [115]; the same tendency was observed in the subchondral bone [116] and synovial fluid [117]. Overall, both studies suggested that taurine is relevant to sclerosis. Complementarily, targeted metabolomics also revealed higher levels of taurine, such as phenylalanine and proline, in osteophyte cartilage samples than in control samples, which were related to collagen dissolution or the destruction of boundary layers involved in osteophyte formation [118].
In addition to taurine’s participation as a crucial element in OA, various studies have highlighted its beneficial effects as a possible treatment. For example, daily taurine intake induces high SOD activity in the blood of patients with knee OA [119], whereas taurine tail vein injection relieved OA symptoms in a rat surgery model in a dose- and time-dependent manner, alleviating allodynia and knee swelling inhibition. A histopathological analysis revealed the suppression of matrix loss and cartilage degeneration and the downregulation of matrix metalloproteinase-3 (MMP-3) and CCAAT-enhancer-binding protein homologous protein (CHOP) expression [120]. Finally, in vitro studies have revealed that taurine exerted an antioxidant effect by alleviating H2O2-induced endoplasmic reticulum stress and subsequently inhibiting patient-derived chondrocyte apoptosis in different grades of OA [121].
Taurine and rheumatoid arthritis
Besides OA, rheumatoid arthritis (RA), another common chronic joint disorder, is an autoimmune inflammatory condition that involves degradation of the extracellular matrix of cartilage. The global estimated prevalence of RA is 0.46% [122]. Although there is no cure for RA, early treatment and supportive treatments can reduce the risk of joint damage and offer better living conditions. In the search for new treatments, several studies have evaluated natural products or existing drugs, and metabolomic approaches have analyzed their effectiveness. Among these, taurine has been identified as a biomarker with the potential to monitor the efficacy of pharmacotherapy. Chinese medicine formulas can regulate taurine and hypotaurine metabolism in the urine samples of the RA rat model [123-125]. Furthermore, the serum metabolic profiles of patients diagnosed with RA showed alterations in taurine levels in response to rituximab [126] and methotrexate treatment [127].
The use of taurine as a therapeutic candidate in RA has been suggested because of its antioxidant and anti-inflammatory properties; this subject was reviewed in detail recently using animal and in vitro research [128]. In brief, these studies suggest the benefits of taurine and its derivatives in managing RA in humans. Fig. 4 summarizes the potential roles of taurine in bone and cartilage under normal and pathological conditions.
Go to :
CONCLUSION
Overall, the present data indicate that taurine plays an important role in bone development and function. We emphasized the ability of taurine to control osteoblastogenesis, osteoclastogenesis, and maintenance of chondrocyte phenotype. Indeed, taurine is emerging as a key player in bone disorders and a promising therapeutic approach for diseases such as osteoporosis and OA. However, more research and clinical studies are needed to elucidate the details of the taurine-bone relationship and the therapeutic efficacy of taurine.
Go to :
 XML Download
XML Download