

 PDF
PDF Citation
Citation Print
Print



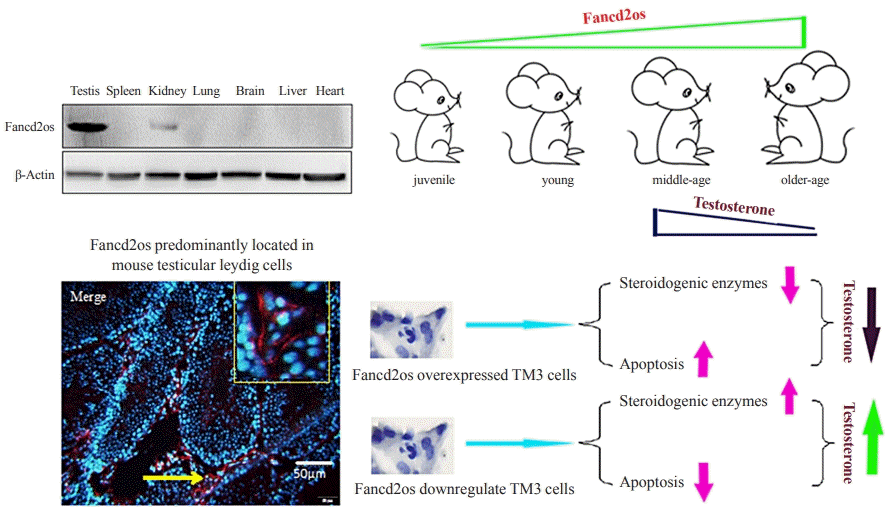
INTRODUCTION
It is well known that testicular Leydig cells, which are distributed in the interstitial compartment of the testes and neighboring testicular seminiferous tubules, are the main source of testosterone biosynthesis and play an essential role in the initiation of spermatogenesis and maintenance of male reproductive function [1]. Hence, the balance between proliferation and cell death of Leydig cells is pivotal for testosterone production [2]. Apoptosis is a physiological process of cell death that is instrumental in maintaining various biological functions. Leydig cell dysfunction disturbs testicular development and can even lead to infertility. It is well documented that numerous factors, including age, exercise, nutrients, body weight, and gene expression patterns, can affect Leydig cell development and testosterone synthesis [3,4]. A longitudinal study on age-related changes in serum testosterone levels documented that the decline in serum testosterone levels was correlated with the incidence of hypogonadism in men as they aged [5]. These results partially clarify the decline of spermatogenesis in old men [6], but the precise underlying mechanisms remain to be elucidated.
Under physiological conditions, testosterone levels are strictly regulated by the hypothalamic-pituitary-testicular axis [7]. In this axis, the steroidogenic acute regulatory protein (StAR), a nuclear-encoded mitochondrial protein, facilitates the translocation of cholesterol from the mitochondrial outer membrane to the inner membrane to serve as the substrate for testosterone synthesis [8,9]. Therefore, StAR-mediated transportation of cholesterol is viewed as the rate-limiting step in testosterone synthesis, with StAR being the rate-limiting protein. Additionally, P450 cholesterol side-chain cleavage (P450scc) and 3β-hydroxysteroid dehydrogenase (3β-HSD) play equally vital roles in testosterone biosynthesis; the former transforms cholesterol into pregnenolone, while the latter converts pregnenolone to testosterone in the smooth endoplasmic reticulum [3]. The synergy between StAR, P450scc, and 3β-HSD ensures testosterone biosynthesis in Leydig cells [10].
Testosterone production depends not only on the activity of Leydig cells, but also on their number [11]. A reduced number of Leydig cells has been noticed in men as they age [12], which is mainly due to excessive apoptosis of Leydig cells [13]. Ultraviolet (UV) exposure has been documented to induce apoptosis in a variety of cells. Apoptosis in cells starts with cell shrinkage and membrane budding, followed by dense chromatin condensation and fragmentation [14]. Activation of caspase is a crucial biochemical event in UV-induced apoptosis [15]. Some proteins such as Bcl-2 and Bcl-xL could inhibit apoptosis induced by various factors, including UV irradiation, in a variety of cells [16].
Fancd2 opposite-strand (Fancd2os), also known as C3orf24, is a predominantly expressed gene in murine testes that encodes a protein with 178 amino acids [17]. However, the distribution of the Fancd2os protein and its spatiotemporal expression patterns have not been thoroughly investigated. Moreover, the function of the Fancd2os protein and its underlying mechanism have not been reported to date. Herein, it was demonstrated that Fancd2os was highly expressed in mouse testes, especially in testicular Leydig cells. In addition, the results established that Fancd2os can regulate testosterone synthesis via both steroidogenic enzymes and the apoptotic pathway. The aim of this study was to investigate the expression patterns of Fancd2os and elucidate its functions in testosterone production, providing novel insights for understanding the mechanism of the age-related decline in serum testosterone.
Go to : 
METHODS
Animals and treatment
Different aged adult BALB/c male mice (purchased from the Laboratory Animal of Shanxi Medical University) were used to examine the spatial expression pattern of Fancd2os. Blood samples were harvested from the abdominal aorta, and testicular tissue samples were collected to assess the spatial expression pattern of Fancd2os via real-time polymerase chain reaction (RT-PCR) and Western blot analysis after the mice were anesthetized with sodium pentobarbital (75 mg/kg body weight intraperitoneally).
Finally, immunofluorescence (IF), immunohistochemistry (IHC), and terminal deoxynucleotidyl transferase-mediated dUTP-biotin nick end labeling (TUNEL) staining of testicular paraffin sections were performed. All animal experiments were approved by the Animal Ethics Committee of Shanxi Medical University (permit no. SYDL -2017-16) and complied with the National Institutes of Health guide for the care and use of laboratory animals (NIH Publications No. 8023, revised 1978).
Cell culture and treatment
GC-1 spg and GC-2 spd cells were purchased from the American Type Culture Collection (ATCC, Manassas, VA, USA) and cultured in Dulbecco’s Modified Eagle’s Medium (DMEM, Gibco, Thermo Fisher, Waltham, MA, USA) with 10% fetal bovine serum (FBS, Gibco, Thermo Fisher). TM3 and TM4 cells were obtained from the National Infrastructure of Cell Line Resource (Beijing, China) and grown in a mixture of DMEM, Ham’s F12 medium (Gibco, Thermo Fisher), 5% horse serum (Sigma, St. Louis, MO, USA), and 5% FBS at 37°C under a 5% CO2 atmosphere. Protein extraction was conducted to prepare for Western blot analysis, RNA extraction was achieved using TRIzol for the RT-PCR assay.
Construction of TM3 stable cells with fancd2os overexpression or gene silencing
For the fancd2os overexpression assay, lentivirus expression particles (Lv5(EGla-GFP/Puro)-fancd2os, titer: 1×109) were constructed and prepared by GenePharma Co. Ltd. (Suzhou, China). TM3 cells were seeded into 6-well plates in an antibiotic-free medium and infected with an Lv5(EGla-GFP/Puro)-fancd2os (100 multiplicity of infection [MOI]) according to the manufacturer’s instructions. Forty-eight hours later, the cells were screened with 1 μg/mL puromycin and referred to as Fancd2os/TM3. TM3 cells infected with Lv5(EGla-GFP/Puro) lentivirus were used as the negative control and named vector/TM3.
For the fancd2os knockdown assay, lentivirus RNA interference particles (LV3-fancd2os-Mus-253, sequence: 5´-GCTGCGGCACTACACCTAC-3´; LV3-fancd2os-Mus-364, sequence: 5´-GCTTGTCCTAGACAGACCACT-3´; LV3-fancd2os-Mus-430, sequence: 5´-GCTCCGAGCCGTGAGTAACAA-3´, titer: 7×108) were constructed and prepared by GenePharma Co. Ltd. TM3 cells were seeded into 6-well plates in an antibiotic-free medium and infected with the lentivirus particles mentioned above (100 MOI) according to the manufacturer’s instructions. Forty-eight hours later, the cells were screened with 1 μg/mL puromycin. TM3 cells infected with LV3-NC (5´-TTCTCCGA-ACGTGTCACGT-3´) served as the negative control.
Total RNA extraction and RT-PCR
Total RNA from mouse tissues and GC-1, GC-2, TM3, TM4, or TM3 stable cells were isolated using the TRIzol reagent (Invitrogen, Waltham, MA, USA), and RT-PCR was performed using the UltraSYBR two-step RT-qPCR kit (CWBIO, Taizhou, China) according to the manufacturer’s protocol. Relative Fancd2os mRNA levels were normalized against the housekeeping gene β-actin. The 2−ΔΔCT method was used to calculate the relative levels of gene expression. The primers for PCR were as follows. A forward primer, 5´-TTCGGTAGGGTCATCACAG-3´, and a reverse primer, 5´-AGTCGCATAGAGCAAGAGG-3´, were used to amplify fancd2os. A forward primer, 5´-AACTTGTGGACCGCATGGAGGC-3´ and a reverse primer, 5´-ACAGCGCACGCTCACGAAGT-3´ were used to amplify StAR. A forward primer, 5´-AAGACTTTGGTACAGGTGGCT-3´ and a reverse primer, 5´-AGGATGAGGCTGAACTTGGTC-3´ were used to amplify P450scc. A forward primer, 5´-CTCTACTGGCTTGCCTTCCTG-3´ and a reverse primer, 5´-TGCTGTTCCACTATTGTCCCT-3´ were used to amplify 3β-HSD. A forward primer, 5´-CTATTGGCAACGAGCGGT-3´ and a reverse primer, 5´-GGTCTTTACGGATGTCAACG-3´ were used to amplify β-actin.
Immunohistochemistry and immunofluorescence
Mouse testicular tissues were isolated, fixed with 4% paraformaldehyde, embedded in paraffin, and then sectioned at 4 μm. For IHC staining, the slides were warmed at 60°C for 1 hour, deparaffinized in xylene, and rehydrated. The slides were incubated in 3% H2O2 for 10 minutes to block endogenous peroxidase activity, followed by 3% bovine serum albumin (BSA) for 1 hour. The slices were then incubated with a 1:100 diluted Fancd2os polyclonal antibody (No.102379, antigen used and epitope recognized: N-19, Santa Cruz Biotechnology, Dallas, TX, USA) at 4°C overnight. Detection was performed using the SABC-AP Kit (BOSTER Biological Technology, Wuhan, China) according to the manufacturer’s protocol. Next, the sections were counterstained with hematoxylin and viewed under a CKX51 light microscope (Olympus, Tokyo, Japan). For tissue IF staining, 4-μm slices were sectioned and performed according to the published methods [18]. For cellular IF staining, fancd2os/TM3 or vector/TM3 cells were crawled on the slides, fixed in 4% methanol-free formaldehyde and phosphate-buffered saline (PBS) for 15 minutes, followed by washing twice with PBS. Slides were blocked with 5% BSA in PBS for 2 hours at 37°C. Blocked samples were incubated with Fancd2os antibody (1:100) in PBS containing 2% BSA overnight at 4°C; the next day, the slices were washed with PBS three times for 10 minutes each time and then incubated with the Alexa Fluor 546-conjugated goat anti-rabbit antibody (Life Technologies, Thermo Fisher, Waltham, MA, USA) for 1 hour at 37°C in a dark box, followed by washing three times with PBS for 10 minutes each time. Finally, the samples were stained with 4′,6-diamidino-2-phenylindole (DAPI, Sigma) as the nuclear counterstain. Fluorescence signals were analyzed using an FV1000 confocal laser scanning microscope (Olympus). Finally, five fields were randomly selected from each slice, and at least 100 cells were counted in each field. All experiments were repeated in triplicate for each group.
Testosterone concentration assay
Blood samples were placed in 1.5-mL microcentrifuge tubes at 4°C overnight and subsequently centrifuged at 2,000 rpm for 20 minutes, and the supernatant was collected. The testosterone concentration of both serum and cell supernatants was measured using a testosterone ELISA Kit (Elabscience, Houston, TX, USA) according to the manufacturer’s protocol. Each experiment was independently repeated three times, and in each of these three biological repetitions, three samples were prepared.
Detection of apoptosis
TM3, vector/TM3, and fancd2os/TM3 cells were seeded in 6-well plates at a density of 2×105 cells/well and treated with 20 J/m2 UV light for 6 hours. Next, the cells were digested by 0.25% trypsin and centrifuged at 2,000 rpm for 5 minutes. The acquired cells were stained with annexin V-fluorescein isothiocyanate (FITC) to quantify the percentage of cells undergoing apoptosis, whereas the necrotic cells were counterstained with propidium iodide (PI) as previously described [19]. Apoptosis was analyzed using flow cytometry (BD Biosciences, Franklin Lakes, NJ, USA).
To detect the apoptosis of mouse testicular Leydig cells, paraffin-embedded sections were cut into 4-μm slices and deparaffinized, and a TUNEL assay was performed according to the manufacturer’s instructions using a TUNEL assay kit (Beyotime, Shanghai, China). Murine testicular Leydig cells, including TUNEL-positive cells, were counted under a CKX51 light microscope, and the apoptotic rate was calculated as the ratio of TUNEL-positive Leydig cells to total Leydig cells. Five fields were randomly chosen from each slice, and at least 100 cells were counted in each field. All the experiments were repeated in triplicate for each group.
Western blots
Mouse tissues were ground in liquid nitrogen, and then total protein was extracted using sodium dodecyl sulfate (SDS) lysis buffer (50 mM Tris–HCl [pH 6.8], 10% glycerol, and 2% SDS). Samples were separated in a 12% polyacrylamide gel and then transferred to polyvinylidene difluoride membranes (Millipore, Burlington, MA, USA). The membranes were blocked in 5% skimmed milk for 1 hour at room temperature and subsequently probed at 4°C overnight with the indicated primary antibodies. The next day, the membranes were incubated with horseradish peroxidase-conjugated secondary antibodies (1:5,000) and examined using enhanced chemiluminescence (Transgen, Beijing, China). The following primary antibodies were used (1:1,000 unless otherwise indicated): fancd2os (No.102379, Santa Cruz Biotechnology), cleaved-caspase 3 (No.9664, CST, Danvers, MA, USA), Bax (No.14796, CST), Bcl-2 (No.3498, CST), Bcl-xL (No.2764, CST), and β-actin (No.3700, 1:3,000, CST).
Statistical analysis
The data were expressed as mean±standard deviation from at least three independent experiments. The Student’s t test was used for comparisons involving two groups, and one-way analysis of variance was used for multi-group comparisons. Pearson coefficients were used to analyze correlations between the expression of Fancd2os and apoptosis in mouse testicular Leydig cells. An r≥0.5 was considered to indicate a strong correlation, while an r<0.5 was considered to reflect a poor correlation [20]. The threshold for statistical significance was set at P<0.05. All statistical analyses were performed using Prism version 6.0 (GraphPad Software, La Jolla, CA, USA).
Go to :
RESULTS
Fancd2os is highly expressed in the testes and is primarily located in murine Leydig cells
Mouse Fancd2os is located on chromosome 6, and its gene number in GeneBank is NM_027633.3. This gene contains four exons and encodes a protein with 178 amino acids. As displayed in Fig. 1A, there are evolutionarily conserved variations in the Fancd2os protein among different genera, with 96.87% similarity in the amino acid sequence. To explore the expression patterns of Fancd2os, mature adult male mouse (6 months old) [21] tissues were collected, and Fancd2os expression was examined using RT-PCR and Western blots. These experiments showed that adult mouse testes exhibited a higher expression level of Fancd2os than other tissues (Fig. 1B, C). Moreover, testes from juvenile mice (1 month old), young mice (3 months old), middle-aged mice (12 months old), and aged mice (24 months old) [21] were analyzed to assess the temporal expression of Fancd2os. As illustrated in Fig. 1D, E, Fancd2os expression showed a dynamic variation throughout the development of the mice. The expression of Fancd2os mRNA was low in young mice, rose with age, and then plateaued during the old period. Furthermore, fancd2os protein levels exhibited similar expression patterns.
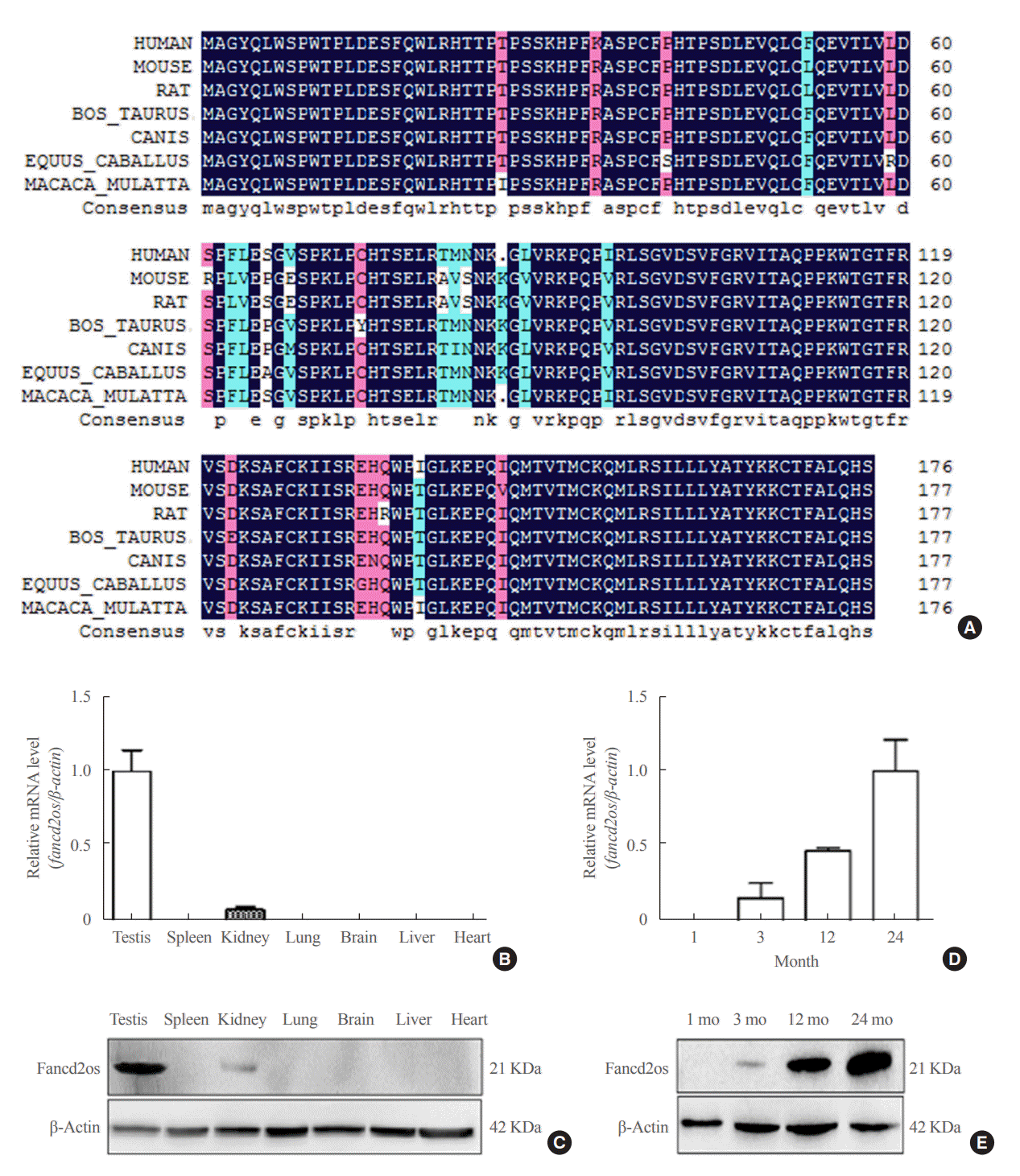 | Fig. 1.The expression of Fancd2 opposite-strand (Fancd2os) in a panel of mouse tissues and its temporal expression in developing mouse testes. (A) Alignment of the amino acid sequence of Fancd2os in mice (NP_081909.2), humans (NP_001158311.1), rats (NP_001102721.1), Macaca mulatta (NP_001181271.1), Bos taurus (NP_001070589.1), Equus caballus (XP_023475748.1), and Canis lupus (XP_
005632228.1). The alignment was performed by DNAMAN (Lynnon). Homology levels are highlighted in different colors: black, 100%; pink, 75%; blue, 50%. (B, C) The tissue distribution of Fancd2os mRNA in adult mice was analyzed by (B) quantitative real-time polymerase chain reaction (RT-PCR) and (C) Western blots. (D, E) The mRNA and protein levels of Fancd2os in the testes from juvenile, young, middle-aged, and aged mice were detected by quantitative RT-PCR and Western blots. β-Actin served as a loading control. Each assay was repeated three times with similar results. Each number represents the age of the mice.
|
In order to explore the function of Fancd2os, the distribution of Fancd2os in mature adult murine testes was investigated using IF staining. As outlined in Fig. 2A, Fancd2os-positive cells were distributed in the germinal cells as well as testicular Leydig cells. Conversely, the Fancd2os protein was identified in the cytoplasm. Afterward, the expression of Fancd2os in GC-1 spg cells (a mouse spermatogonia cell line), GC-2 spd cells (a mouse spermatocyte cell line), TM3 cells (a mouse Leydig cell line), and TM4 cells (a mouse Sertoli cell line) was explored using quantitative RT-PCR (qRT-PCR) and Western blots. The results indicated that Fancd2os was mainly expressed in TM3 and TM4 cells (Fig. 2B, C). Next, the location of Fancd2os was determined via a cellular IF assay using Fancd2os antibody in the TM3 cells that were infected with Lv5(EGla-GFP/Puro)-fancd2os to stably express Fancd2os. The results also corroborated that Fancd2os was distributed in the cytoplasm (Fig. 2D).
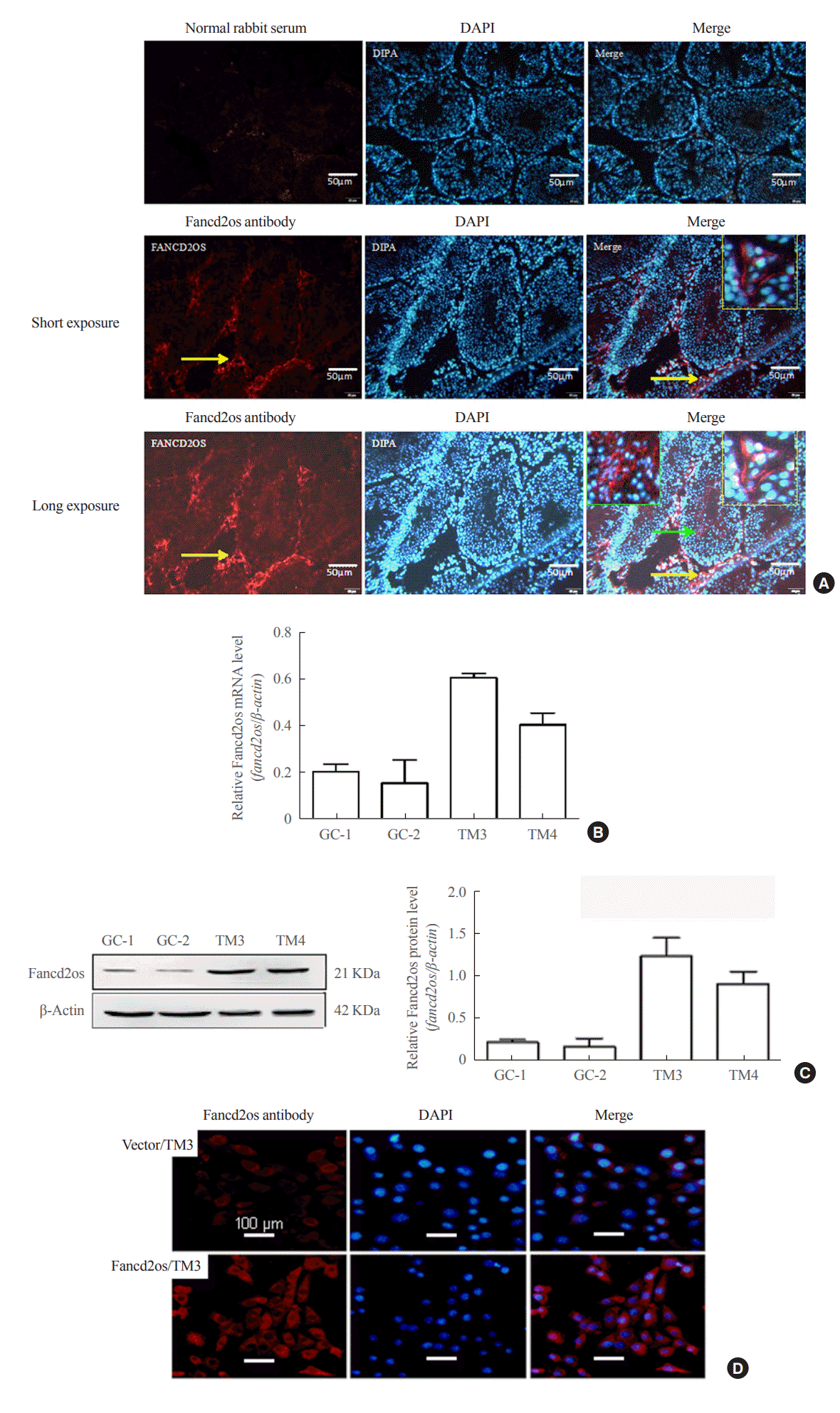 | Fig. 2.Fancd2 opposite-strand (Fancd2os) is primarily distributed in mouse testicular Leydig cells and localized in the cytoplasm. (A) Immunofluorescence analysis was performed with an anti-Fancd2os polyclonal antibody (bottom panel) to detect the localization of Fancd2os in adult mouse testes. A negative control was performed using rabbit immunoglobulin G (upper panel). The yellow arrows indicate Fancd2os-positive cells, and the yellow boxes represent the magnified field of view. The green arrows indicate Fancd2os-positive cells in seminiferous tubule, and the green boxes represent the magnified field of view. Scale bars represent 50 μm. (B, C) The cellular distribution of Fancd2os mRNA and protein in four mouse testes-related cell lines were measured by (B) quantitative real-time polymerase chain reaction and (C) Western blots. β-Actin served as a loading control. Each assay was repeated three times with similar results. (D) TM3 cells that stably expressed Fancd2os were subjected to immunofluorescence analysis with an anti-Fancd2os antibody followed by incubation with Alexa Fluor 546 goat anti-mouse antibody. Fluorescence signals were analyzed using confocal laser scanning microscopy. Nuclei were stained with 4′,6-diamidino-2-phenylindole (DAPI). Scale bars represent 100 μm. DAPI, dapidiamidino-phenyl-indole.
|
Fancd2os reduces testosterone production in TM3 cells via the downregulation of steroidogenic enzymes
To evaluate the putative functions of Fancd2os, the TM3 cell line, in which Fancd2os was highly expressed, was utilized as a cell model. Stable Fancd2os/TM3 and vector/TM3 cells were prepared by infecting them with lentivirus expression particles, selected by puromycin, and confirmed by qRT-PCR and Western blots (Fig. 3A, B). Simultaneously, stable Fancd2os knockdown TM3 cells were constructed by infecting them with lentivirus knockdown particles, selected by puromycin and confirmed by qRT-PCR and Western blots (Fig. 3C, D). The results confirmed that TM3 cells with Fancd2os stable overexpression or knockdown were established.
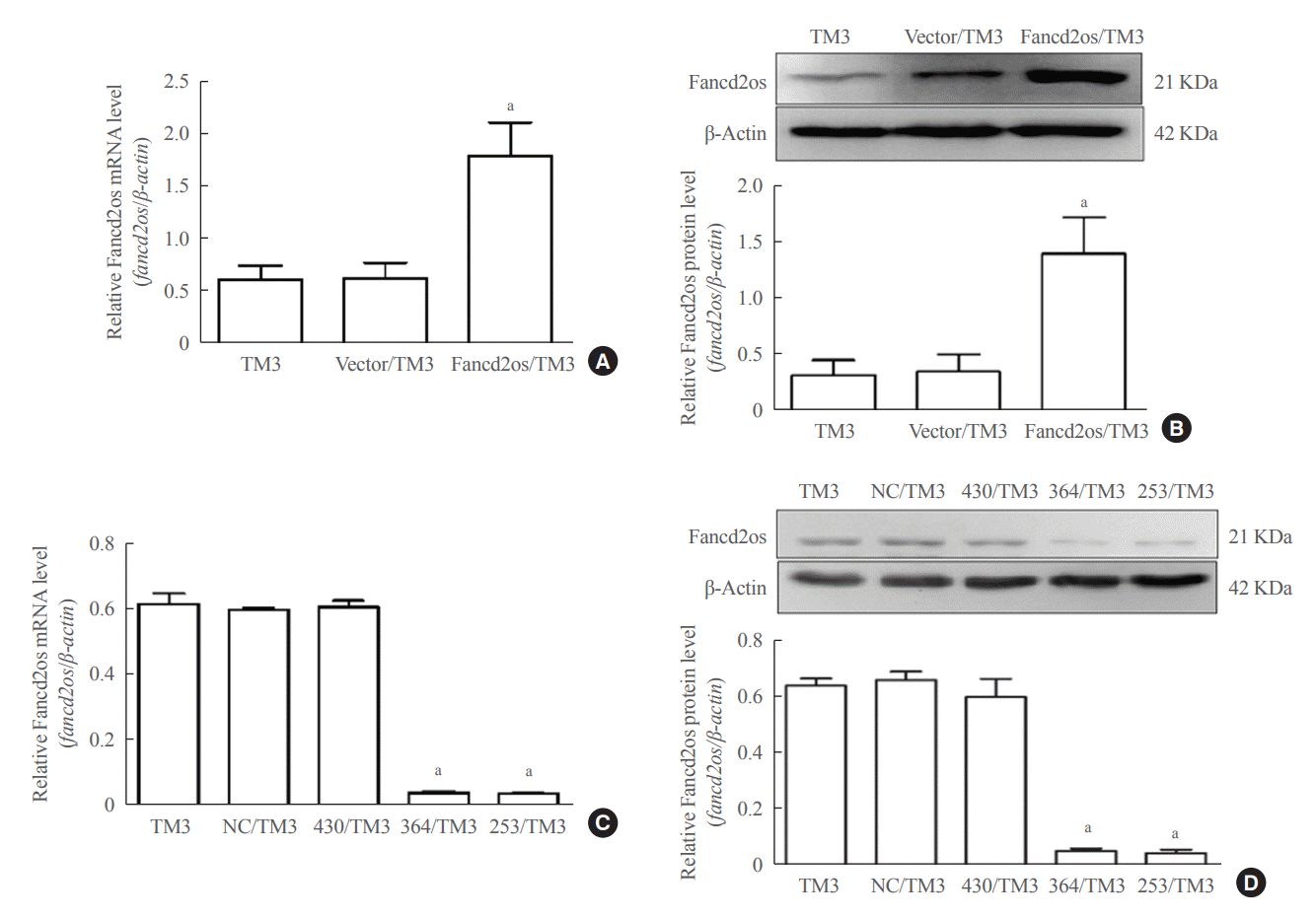 | Fig. 3.Stable Fancd2 opposite-strand (Fancd2os)-overexpressing TM3 or stable Fancd2os knockdown TM3 cells were constructed. TM3 cells were infected with Lv5(EGla-GFP/Puro)-fancd2os, LV3-fancd2os-Mus-253/364/430, or vector following puromycin screen, the stable Fancd2os/TM3 and stable Fancd2os knockdown TM3 cells were confirmed using (A, C) real-time polymerase chain reaction and (B, D) Western blots. Each bar represents the mean±standard deviation from three separate experiments. Significant difference compared to TM3, vector/TM3 or NC/TM3. aP<0.01.
|
Based on the knowledge that Leydig cells are the primary sources of testosterone production [22], testosterone levels were evaluated in Fancd2os-overexpressing TM3 cells and Fancd2os knockdown TM3 cells. As depicted in Fig. 4A, testosterone levels were lower in the Fancd2os-overexpressing TM3 cells than in TM3 or vector/TM3 cells. On the contrary, Fancd2os knockdown boosted testosterone levels in TM3 cells (Fig. 4B). These data indicate that Fancd2os reduces testosterone production.
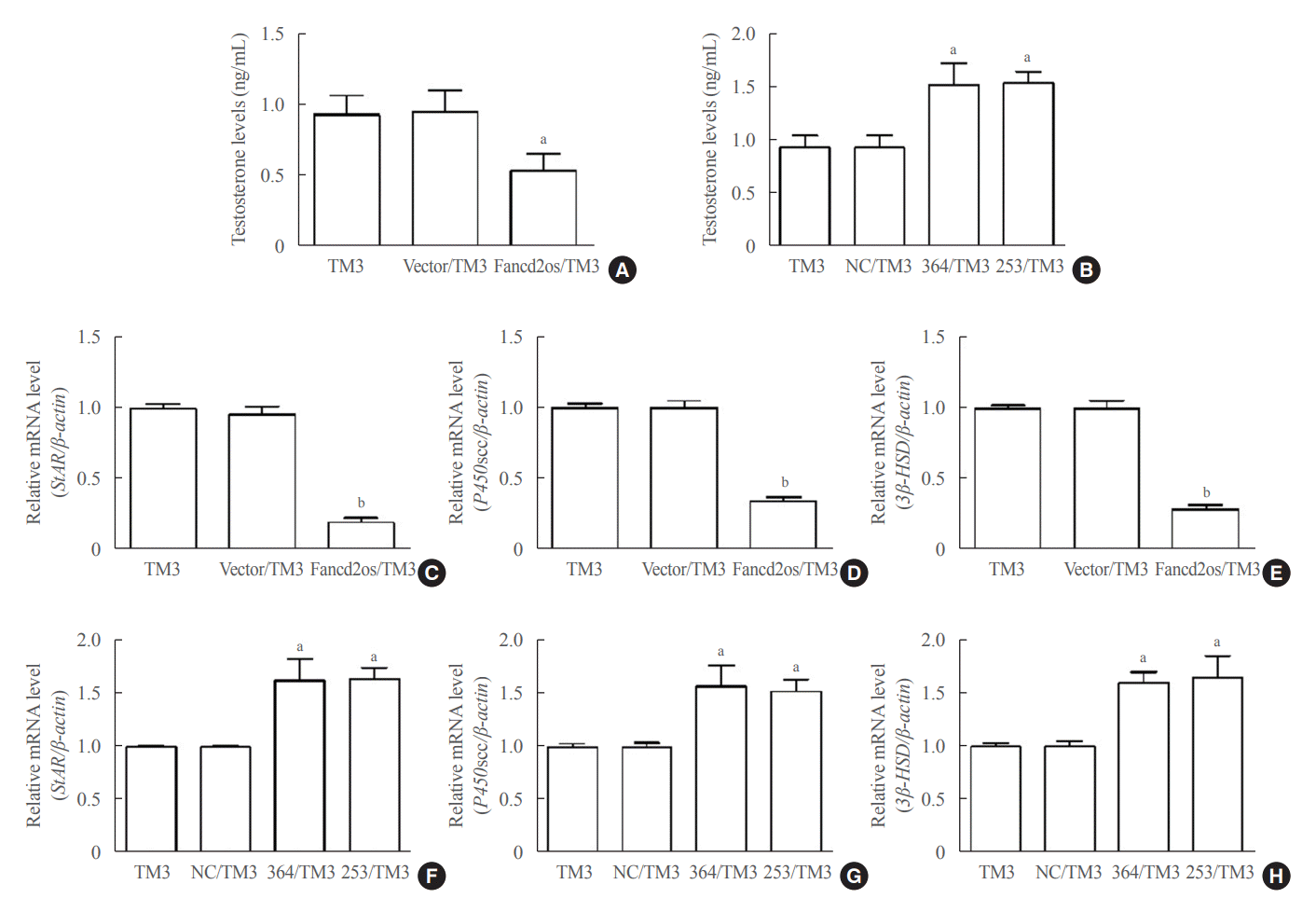 | Fig. 4.Fancd2 opposite-strand (Fancd2os) reduces testosterone production and steroidogenic enzyme expression in TM3 cells. The testosterone levels in Fancd2os-overexpressing (A) or knockdown TM3 cells (B) were detected by enzyme-linked immunosorbent assays (n=3 per group). (C, D, E) Relative quantities of mRNA expression of steroidogenic acute regulatory protein (StAR), P450 cholesterol side-chain cleavage (P450scc), and 3β-hydroxysteroid dehydrogenase (3β-HSD) in Fancd2os-overexpressing TM3 cells or Fancd2os knockdown TM3 cells (F, G, H) were determined real-time polymerase chain reaction using β-actin as a housekeeping gene. Each bar represents the mean±standard deviation from three separate experiments. Significant difference compared to TM3, vector/TM3 or NC/TM3. aP<0.05; bP<0.01.
|
Mouse TM3 Leydig cells produce male hormones by increasing the levels of steroidogenic enzymes related to testosterone production in the testes [23]. During testosterone biosynthesis, StAR protein, P450scc, and 3β-HSD have been regarded as three pivotal rate-limiting molecules [24]. Therefore, the levels of StAR, P450scc, and 3β-HSD were assessed in Fancd2os-overexpressing TM3 cells and Fancd2os knockdown TM3 cells. We found that the StAR, P450scc, and 3β-HSD levels were lower in Fancd2os-overexpressing TM3 cells than in TM3 or vector/TM3 cells (Fig. 4C-E). Contrarily, Fancd2os knockdown enhanced the StAR, P450scc, and 3β-HSD levels in TM3 cells (Fig. 4F-H). These data indicate that Fancd2os diminishes StAR, P450scc, and 3β-HSD production.
Fancd2os decreases testosterone levels in TM3 cells by promoting cellular apoptosis
Apoptosis of Leydig cells severely affects testosterone biosynthesis and secretion [25]. To further explore the underlying mechanism of Fancd2os in reducing testosterone levels, we next determined whether apoptosis was involved in the modulatory role of Fancd2os on testosterone levels using Fancd2os-overexpressing or knockdown TM3 cells. In the apoptotic assay, UV radiation was used to induce apoptosis, since it can evoke apoptosis in various cells, including germ cells [26]. As outlined in Fig. 5A, B, flow cytometry determined that overexpression of Fancd2os promoted the apoptosis of TM3 cells (annexin V-positive, PI-positive) treated with UV stimulation compared with controls. In contrast, the knockdown of Fancd2os resulted in the inhibition of cellular apoptosis in TM3 cells. Further Western blot analysis of total proteins isolated from Fancd2os-overexpressing TM3 cells exposed to UV revealed an enhanced level of cleaved caspase-3 and a decreased level of Bcl-2, which are hallmarks of apoptosis (Fig. 5C) [27]. Interestingly, the level of cleaved caspase-3 decreased, while that of Bcl-2 increased, in Fancd2os knockdown TM3 cells stimulated with UV (Fig. 5D). These results indicate that Fancd2os inhibits testosterone synthesis in TM3 cells by stimulating cellular apoptosis.
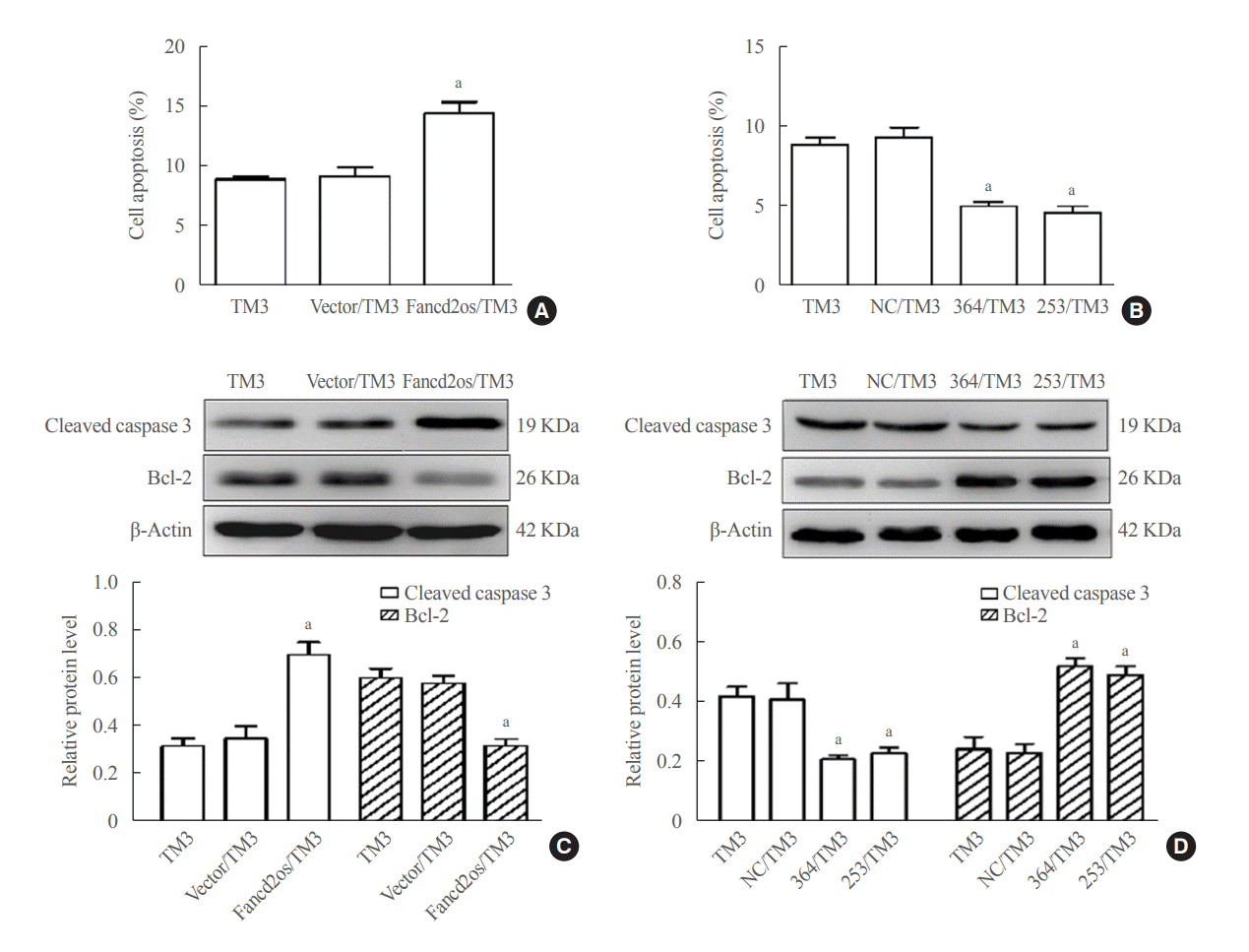 | Fig. 5.Fancd2 opposite-strand (Fancd2os) evokes cellular apoptosis and then attenuates testosterone levels in TM3 cells. Fancd2os-overexpressing (A, C) or knockdown TM3 cells (B, D) were stimulated with 20 J/m2 ultraviolet (UV) for 6 hours, and then double-stained with annexin V-fluorescein isothiocyanate (FITC) and propidium iodide for analyzing apoptosis by flow cytometry assay (A, B), and apoptosis related proteins were determined using Western blots (C, D). Each bar represents the mean±standard deviation from three separate experiments. Significant difference compared to TM3, vector/TM3, or NC/TM3. aP<0.05.
|
Higher levels of Fancd2os are correlated with apoptosis and lower testosterone production in mouse testicular Leydig cells
It is widely accepted that testosterone production decreases with age in men [28]; thus, serum testosterone levels in mice of different ages were investigated. As shown in Fig. 6A, the testosterone serum level increased with age from the juvenile period to middle age, yet its level attenuated from middle-aged to older mice. To explore the correlation between lower serum testosterone levels and higher levels of Fancd2os and apoptosis in mouse Leydig cells, the levels of Fancd2os in murine testes obtained from different aged mice were measured using an IHC assay. As shown in Fig. 6B, Fancd2os was mainly located in mouse testicular Leydig cells, and its levels rose with age. These results are consistent with our previous findings (Figs. 1E, 2A). Additionally, the TUNEL assay demonstrated that apoptosis of mouse Leydig cells gradually increased with age, accompanied by ascending Fancd2os levels (Fig. 6C). To further evaluate the regulatory function of Fancd2os in the apoptosis of Leydig cells, correlations between the expression of Fancd2os and apoptosis in mouse testicular Leydig cells were appraised using Pearson correlation coefficients. As delineated in Fig. 6D, a strong positive correlation was established between the expression of Fancd2os and TUNEL-positive staining (r=0.9609, P=0.0195) (Fig. 6D). Taken together, the data indicate that the increase of Fancd2os promotes apoptosis of Leydig cells, thereby contributing to the decline in testosterone levels. The above results also demonstrate that elevated Fancd2os levels in mouse Leydig cells suppress the testosterone level with age by triggering the apoptosis of Leydig cells.
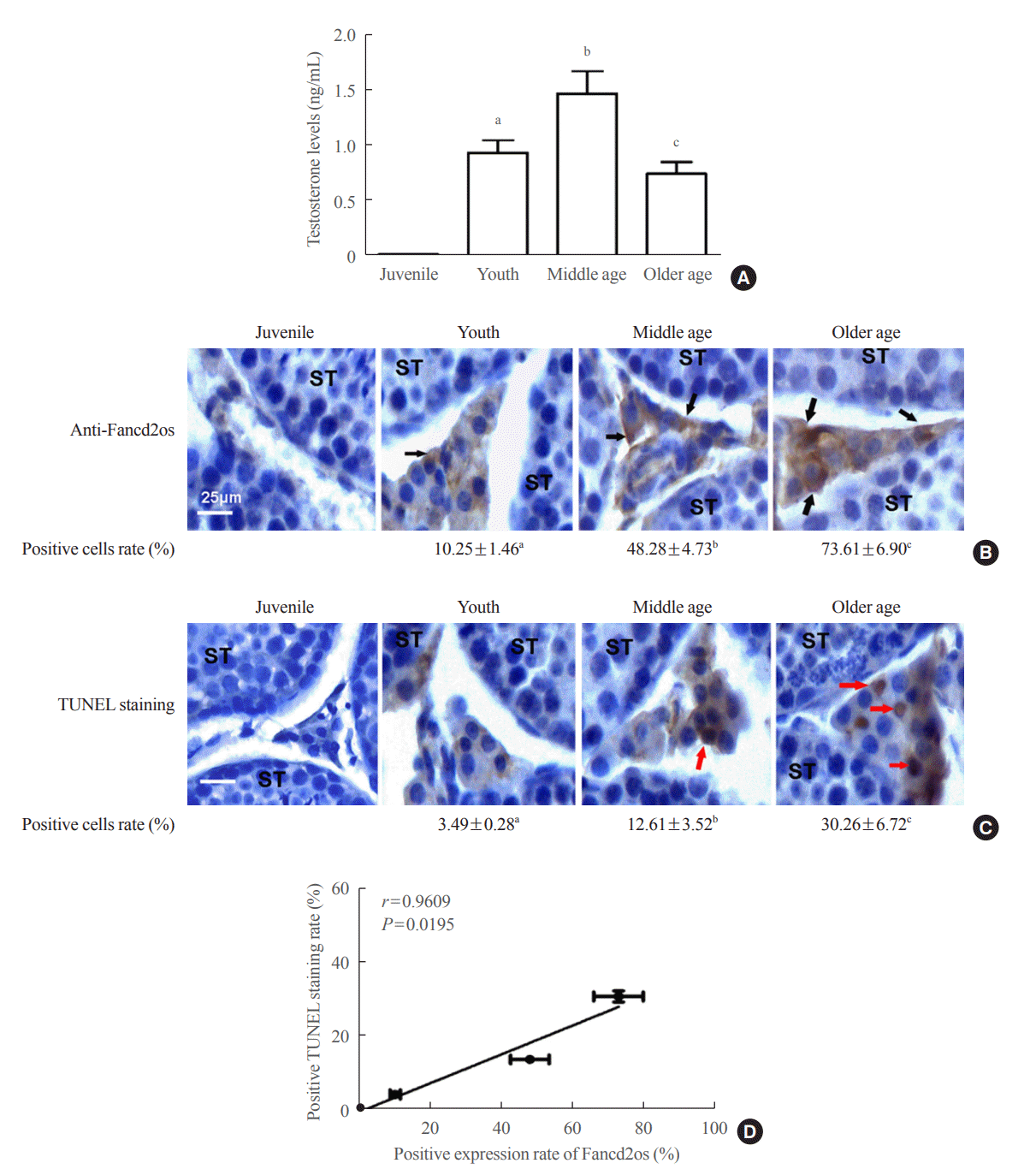 | Fig. 6.Higher Fancd2 opposite-strand (Fancd2os) levels in older mouse Leydig cells result in cellular apoptosis and lower serum testosterone production. (A) The serum testosterone levels from mice of different ages were measured using enzyme-linked immunosorbent assays. (B, C) The testis tissues from different aged mice were sliced and then Fancd2os protein expression and apoptosis were analyzed using immunochemistry and the terminal deoxynucleotidyl transferase-mediated dUTP-biotin nick end labeling (TUNEL) assay, respectively. ST represents seminiferous tubule. Black arrows and red arrows indicate Fancd2os-positive cells and TUNEL-positive cells, respectively. Scale bar represents 25 μm. The results are given as the mean±standard deviation (n=3). (D) The correlations between Fancd2os expression (B) and the TUNEL-positive staining rate (C) were analyzed using Pearson correlation coefficients. An r≥0.5 was considered to indicate a strong correlation, and P<0.05 was considered statistically significant. aP<0.05 compared with juvenile mice; bP<0.05 compared with young mice; cP<0.05 compared with middle-aged mice.
|
Go to :
DISCUSSION
Testosterone, the principal male sex hormone, is produced chiefly in testicular Leydig cells (95% of testosterone is synthesized in Leydig cells) and then released into the blood [18,29]. It plays a pivotal role in spermatogenesis and maintaining secondary sexual functions in males [30]. Low testosterone levels are often associated with delayed puberty, impotence, infertility, or decreased spermatogenesis [31]. It is well documented that testosterone biosynthesis begins in utero, rises sharply in puberty, and then decreases with age in men [32,33]. This age-dependent decline in testosterone is also observed in older male rodents such as rats and mice (>20 months of age) in conjunction with a descent in fertility [34]. However, the mechanism underlying the attenuation in testosterone levels with age has not been fully investigated.
This study identified Fancd2os as an evolutionary conserved protein highly expressed in murine testes by characterizing its expression patterns. Our findings exhibited that Fancd2os was more abundant in adult mouse testes than in other tissues, as well as in older mouse testes than in testes samples from mice of other ages. Moreover, the distribution of Fancd2os in mouse testes was examined, and we found that Fancd2os was predominantly located in the cytoplasm of Leydig cells. Additionally, the expression pattern and distribution of Fancd2os were also validated using four cell lines originating from mouse testes. These results imply that Fancd2os may function as a modulator in testosterone production. Then, TM3, an established mouse Leydig cell line in which Fancd2os is highly expressed (Fig. 2), was used to prepare Fancd2os overexpression or knockdown cellular models. Consistent with our conjecture, Fancd2os overexpression caused a decline in testosterone levels, while its knockdown enhanced testosterone production (Fig. 4).
Under physiological conditions, steroidogenesis is modulated by the binding of luteinizing hormone (LH) to its receptor (LHR) in Leydig cells, which activates the adenylyl cyclase and cyclic adenosine monophosphate-protein kinase A (cAMP-PKA) signaling pathways, leading to the translocation of cholesterol from the cytoplasm to the mitochondria, mediated by StAR. Cholesterol is subsequently converted to pregnenolone by P450scc. In this process, the translocation of cholesterol mediated by StAR is a rate-limiting step; thus, StAR is regarded as a rate-limiting protein in testosterone synthesis [35]. Based on the fact that Fancd2os can mediate testosterone production, the correlation between Fancd2os and the levels of key enzymes related to steroidogenesis was investigated. Our work determined that the overexpression of Fancd2os could attenuate the transcription levels of StAR, P450scc, and 3β-HSD. Conversely, Fancd2os knockdown exerted opposite effects on these enzymes.
Moreover, CYP17A1 is also involved in the synthesis of testosterone. Pregnenolone is further converted by 3β-HSD into progesterone, which is then converted to androstenedione by CYP17A1. Androstenedione is finally converted into testosterone by 17β-HSD [36]. Whether Fancd2os regulates the synthesis of testosterone via CYP17A1 was not examined in the present study. Further research should be undertaken to investigate the expression pattern of CYP17A1 in Fancd2os-overexpressing or Fancd2os-knockdown TM3 cells. The underlying mechanism of Fancd2os’s function in governing the expression of steroid hormone synthetase should also be explored in our next study.
Furthermore, the number and activity of Leydig cells influence steroidogenesis [11]. The former is maintained at equilibrium owing to the balance between proliferation and apoptosis [37]. Leydig cells have only a marginal turnover rate and show low mitotic activity in the testes of adult mammals [12]. Therefore, apoptosis seems crucial for the development, maturation, and death of Leydig cells. Excessive apoptosis may cause a decreased number of Leydig cells and then bring about a decline in testosterone production [13]. Several factors affect the apoptosis of Leydig cells, such as developmental stage, glucocorticoids, certain hormones (including follicle-stimulating hormone, LH, and human chorionic gonadotropin), and testosterone [38]. Furthermore, various proteins are involved in the regulation of cellular apoptosis, such as stem cell factor (SCF)/c-kit, Bcl-2, and Bcl-xL, which inhibit apoptosis, whereas caspase-3, Fas, Bax, and clusterin stimulate apoptosis [39]. Therefore, the role of Fancd2os in the regulation of apoptosis in Leydig cells was assessed. Of note, clinical treatment with radiotherapy in adult men always leads to impaired fertility caused by germ cell apoptosis [40,41]. Herein, UV radiation was selected to induce apoptosis of Fancd2os-overexpressing or knockdown TM3 cells, considering that UV is a common apoptotic stimulator [42,43]. Caspases and the Bcl-2 family are crucial mediators of apoptosis. UV actives caspase-3 by cleaving its precursor form to its active subunits [15]. Overexpression of Bcl-2 prevented UV-induced apoptosis [13]. To determine the mechanism of Fancd2os in the regulation of apoptosis, caspase-3 and Bcl-2 expression were analyzed in Leydig cells with Fancd2os overexpression or knockdown. Fancd2os-overexpressing TM3 cells exposed to UV revealed higher cleaved caspase-3 expression, reduced Bcl-2 expression, and an increased level of apoptosis, while suppression of Fancd2os expression had contrary effects. However, the underlying mechanism of Fancd2os’s function on apoptosis of Leydig cells remains elusive, and further studies in Fancd2os knockout mice are warranted, as a potentially better way to provide comprehensive and rigorous data.
To confirm our in vitro results, serum testosterone levels from mice of different ages were measured. Our data showed that the serum testosterone levels declined with advancing age, although they increased from juvenile to middle age. Furthermore, the expression pattern of Fancd2os and the apoptosis of Leydig cells in murine testes at different ages were assessed. Fancd2os levels gradually rose with age, and the apoptosis of Leydig cells also gradually increased in mouse testes. Although our in vitro assays revealed that overexpressed Fancd2os in TM3 cells promoted cellular apoptosis and decreased testosterone levels, in vivo assays demonstrated that Fancd2os expression and apoptosis both increased with age, while testosterone increased during middle age (when apoptosis and Fancd2os expression increased) and then declined in old age. Although increased Fancd2os expression in cells led to decreased testosterone levels, that was not the case for middle-aged mice, indicating that the situation in vivo is more complex. The data presented herein only suggest the possibility that aging, Fancd2os, apoptosis, and testosterone production might be linked. Further studies including mechanistic experiments are required to confirm these associations.
Collectively, we identified a highly expressed testicular protein, Fancd2os, which is primarily distributed in Leydig cells and has an age-dependent expression pattern. In vivo and in vitro experiments verified that Fancd2os modulates testosterone biosynthesis by regulating either the transcription levels of StAR, P450scc, and 3β-HSD or the apoptosis of Leydig cells. These findings contribute to our understanding of the mechanism of age-related testosterone decline.
Go to :
 XML Download
XML Download