

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
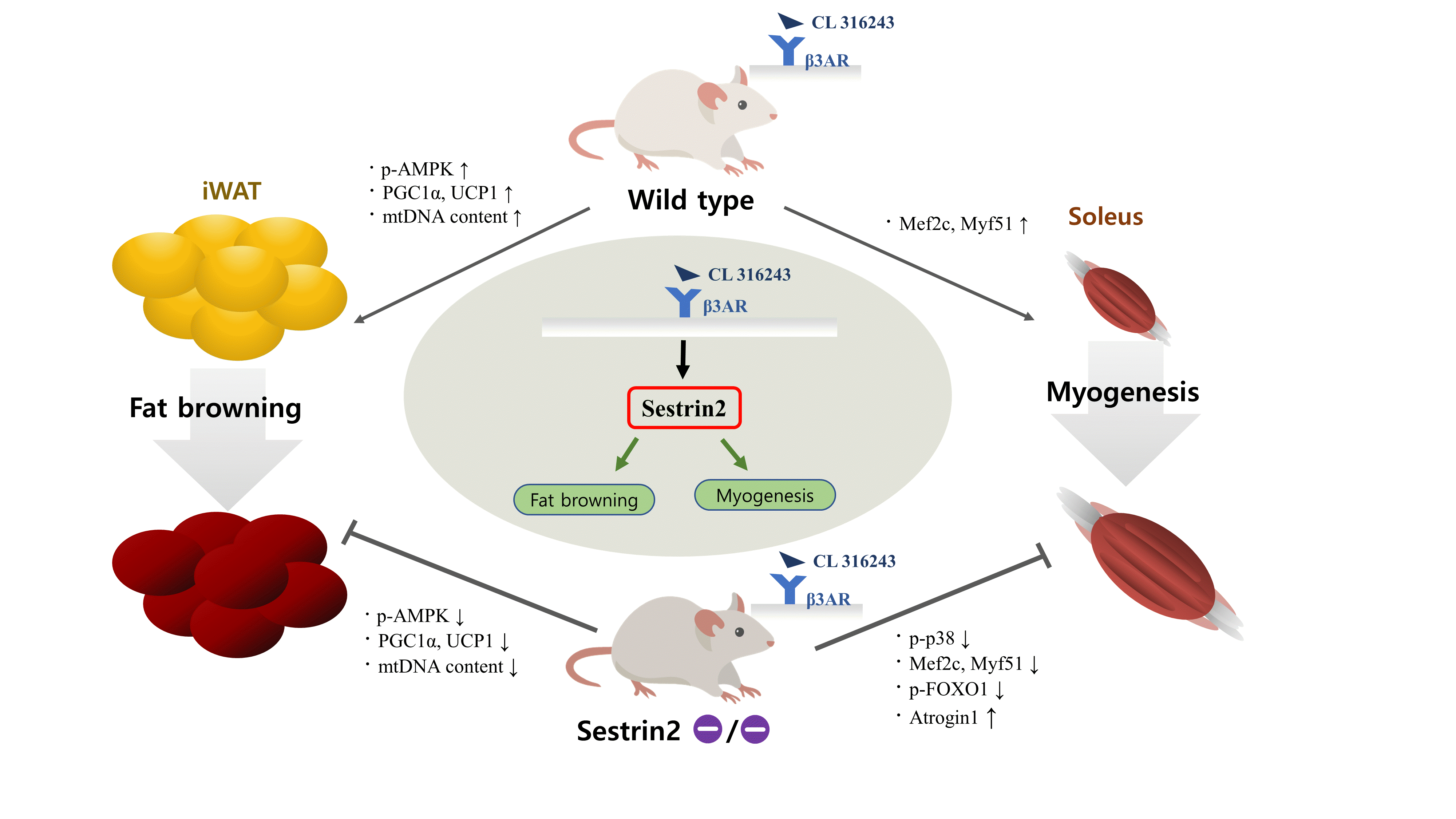
Excessive fat deposition and loss of muscle are referred to as sarcopenic obesity (SO) [1]. SO is closely associated with metabolic diseases, physical disability, and cardiovascular mortality in the elderly. Although physical rehabilitation is recommended for SO treatment, fear of falling and anorexia limit these activities in elderly patients [2]; therefore, pharmacological treatments for SO are needed.
White adipose tissue (WAT) and brown adipose tissue (BAT) are the major types of body fat. The cells in WAT contain unilocular large lipid droplets (LDs) and store excess energy in the form of triglycerides, whereas BAT cells contain multiple small LDs and are involved with energy expenditure [3]. Studies have reported that a subset of adipocytes in WAT can be changed into brown-like adipocytes (called beige adipocytes) through the browning process. Browning, which is induced by β3-adrenergic receptors (β3AR) and cold exposure, has anti-obesity effects [3]. Administration of a β3AR agonist, CL 316,243 (CL), increased multilocular LD formation and energy expenditure in WAT and improved insulin sensitivity in an adenosine monophosphate-activated protein kinase (AMPK)-dependent manner [4]. AMPK, a downstream molecule of β3AR, promotes the browning process. In humans, β3AR activation improves muscle oxidative capacity, pancreatic beta-cell function, and insulin sensitivity [5]. A detailed understanding of β3AR signaling is essential for the treatment of SO.
Sestrin2, an intracellular AMPK activator, has anti-inflammatory effects, and insulin resistance and endoplasmic reticulum dysfunction can occur with sestrin2-deficiency [6,7]. Sestrin2 also regulates energy expenditure and body composition; therefore, sestrin2-knockout (KO) mice showed increased LD size and reduced mitochondria biogenesis in BAT [8]. In addition, sestrin2 blocks immobilization-mediated muscle atrophy via autophagy [9]. Despite its various metabolic effects, the role of sestrin2 in changes in body composition remains unclear. In this experiment, we hypothesized that sestrin2 would act as a mediator in the β3AR-AMPK pathway and sought to clarify the roles of sestrin2 in adipose and muscle tissue under CL treatment in sestrin2-KO mice.
METHODS
Animals
Sestrin2-KO mice with a C57BL/6 background were provided by Dr. M. Karin (University of California, San Diego, USA) and Dr. J.H. Lee (University of Michigan, USA) [6,10]. Wild-type (WT) and sestrin2-KO mice were obtained from a heterozygous cross. Three-month-old male WT mice were randomly divided into vehicle-injected WT mice (WT-Veh, n=6) and CL (Cayman Chemical, Ann Arbor, MI, USA)-injected WT mice (WT-CL, n=6). Three-month-old male sestrin2-KO mice received CL injections (KO-CL, n=6).
Detailed methods and materials are provided in the Supplemental Methods. This study was approved by the Institutional Animal Care and Use Committee (IACUC) of Korea University (Seoul, Korea) (KOREA-2017-0153).
RESULTS
Changes to the β3AR-mediated browning process in the iWAT of sestrin2-KO mice
When we injected WT and sestrin2-KO mice with CL, the rectal temperature increased in the WT mice, and this effect was attenuated by sestin2 deficiency (Fig. 1A). CL treatment reduced the percentage of epididymal white adipose tissue (eWAT) weight to total body weight; however, inguinal white adipose tissue (iWAT) weight was not affected (Fig. 1B). Western blots revealed increases in phosphorylated AMPK, peroxisome proliferator-activated receptor gamma coactivator-1α (PGC1α), uncoupling protein-1 (UCP-1), and sestrin2 levels after CL treatment in iWAT, while these effects were blocked by sestin2 deficiency (Fig. 1C). Through quantitative real-time polymerase chain reaction, we identified that brown fat markers Ucp1, elongation of very long chain fatty acids protein 3 (Elovl3), and cell death activator CIDE-A (Cidea) (Fig. 1D), the mitochondrial biogenesis marker mitochondrial transcription factor A (Tfam) (Fig. 1E), and mitochondrial DNA content (Fig. 1F) all increased after CL treatment in iWAT from WT mice, but these effects were not observed in iWAT from sestrin2-KO mice. As a result, the reduction of the LD diameter observed in iWAT induced by CL was greater in WT mice than in sestrin2-KO mice (Fig. 1G). Unlike in iWAT, phosphorylated AMPK, brown fat markers, mitochondrial biogenesis markers, and mitochondrial DNA content were not affected by CL treatment in eWAT (Fig. 1C-F). However, the LD diameter significantly decreased after CL treatment in eWAT from both WT and sestrin2-KO mice (Fig. 1G). These results suggest that the β3AR-mediated browning process may be regulated by sestrin2, especially in iWAT.
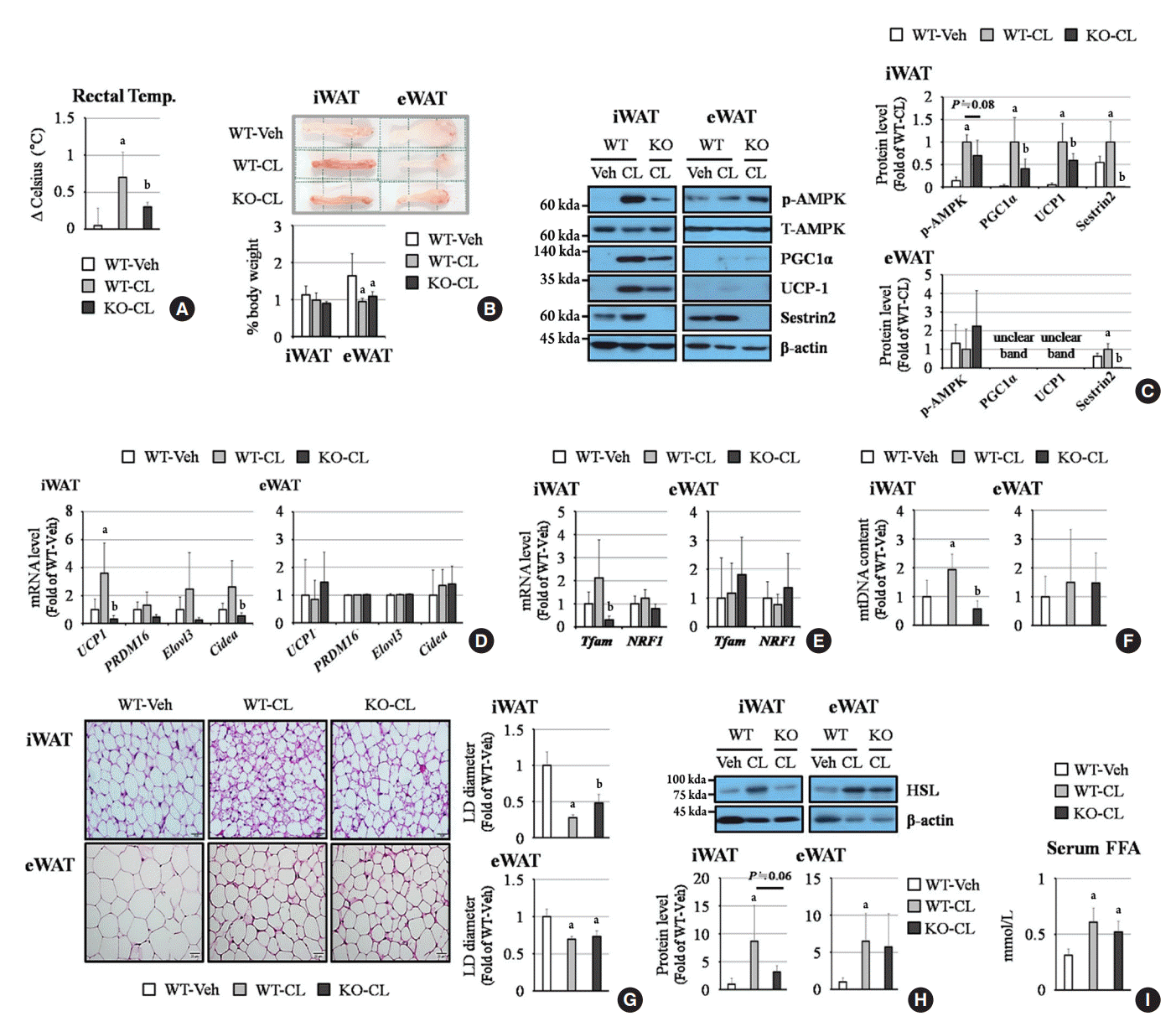
Fig. 1.
Sestrin2 deficiency reduces CL 316,243 (CL)-mediated fat browning in inguinal white adipose tissue (iWAT), but not in epididymal white adipose tissue (eWAT). (A) Rectal temperature changes during 1 hour under vehicle (Veh) or CL treatment. (B) Fat images and weight (per body weight). The side length of a dotted square is 1.2 cm. (C) Western blots of phosphorylated adenosine monophosphate-activated protein kinase (p-AMPK), peroxisome proliferator-activated receptor gamma coactivator-1α (PGC1α), uncoupling protein-1 (UCP-1), and sestrin2. (D, E) Quantitative polymerase chain reaction (PCR) of UCP-1, PR domain containing 16 (PRMD16), elongation of very long chain fatty acids protein 3 (Elovl3), cell death activator CIDE-A (Cidea), mitochondrial transcription factor A (Tfam), and nuclear respiratory factor 1 (NRF1). (F) Ratio of mitochondrial DNA (mtDNA) to genomic DNA (gDNA). (G) Hematoxylin and eosin (H&E) staining; the lipid droplet (LD) diameter was measured from five views for each tissue using ImageJ software. The length of a scale bar is 20 μm. (H) Western blot of hormone-sensitive lipase (HSL) abundance. (I) Measurements of serum free fatty acid (FFA) levels. All graphs were obtained from six mice per group. Error bars represent mean±standard deviation. KO, knockout. aP<0.05 vs. the wild-type (WT)-Veh group; bP<0.05 vs. the WT-CL group, as analyzed with analysis of variance (ANOVA) followed by the Tukey-Kramer test.
![]()
To identify the role of sestrin2 in lipolysis, hormone-sensitive lipase (HSL) levels were determined by Western blots. HSL levels increased after CL treatment in iWAT, and this effect was attenuated by sestrin2 deficiency. In contrast, in eWAT, the CL-mediated increase in HSL expression was not attenuated by sestrin2 deficiency (Fig. 1H). Likewise, serum free fatty acid (FFA) levels were elevated after CL treatment in WT mice and not decreased in sestrin2-KO mice (Fig. 1I). These results indicate that CL-induced lipolysis occurs in a sestrin2-dependent manner in iWAT, but not in eWAT, and this increase of lipolysis in eWAT might be responsible for elevating the serum FFA concentration.
Change of β3AR-mediated myogenesis in the soleus muscle of sestrin2-KO mice
The proportion of soleus muscle mass to total body weight increased after CL treatment in WT mice, but this effect was not observed in sestrin2-deficient mice. The mass of the extensor digitorum longus (EDL), tibialis anterior, gastrocnemius, and quadriceps was not affected by CL treatment (Fig. 2A). Western blots revealed that p38 phosphorylation and sestrin2 levels, but not AMPK phosphorylation, tended to be elevated after CL treatment in the soleus, but these responses were abrogated by sestrin2 deficiency (Fig. 2B). CL-mediated increases in myogenic markers myocyte enhancer factor 2C (Mef2c) and myogenic factor 5 (Myf5) were observed in the soleus of WT mice, but not in sestrin2-KO mice. In contrast, in the EDL, the CL-induced elevation of myogenic markers was detected in both WT and sestrin2-KO mice (Fig. 2C). Under CL treatment, muscle atrophy-related events, including reduced forkhead box O1 (FOXO1) phosphorylation and increased atrogin1 expression, were observed in the soleus, but not in the EDL, from sestrin2-KO mice (Fig. 2D). These results suggest that under CL treatment, myogenic markers decrease, and muscle atrophic molecules increase in the soleus of sestrin2-KO mice.
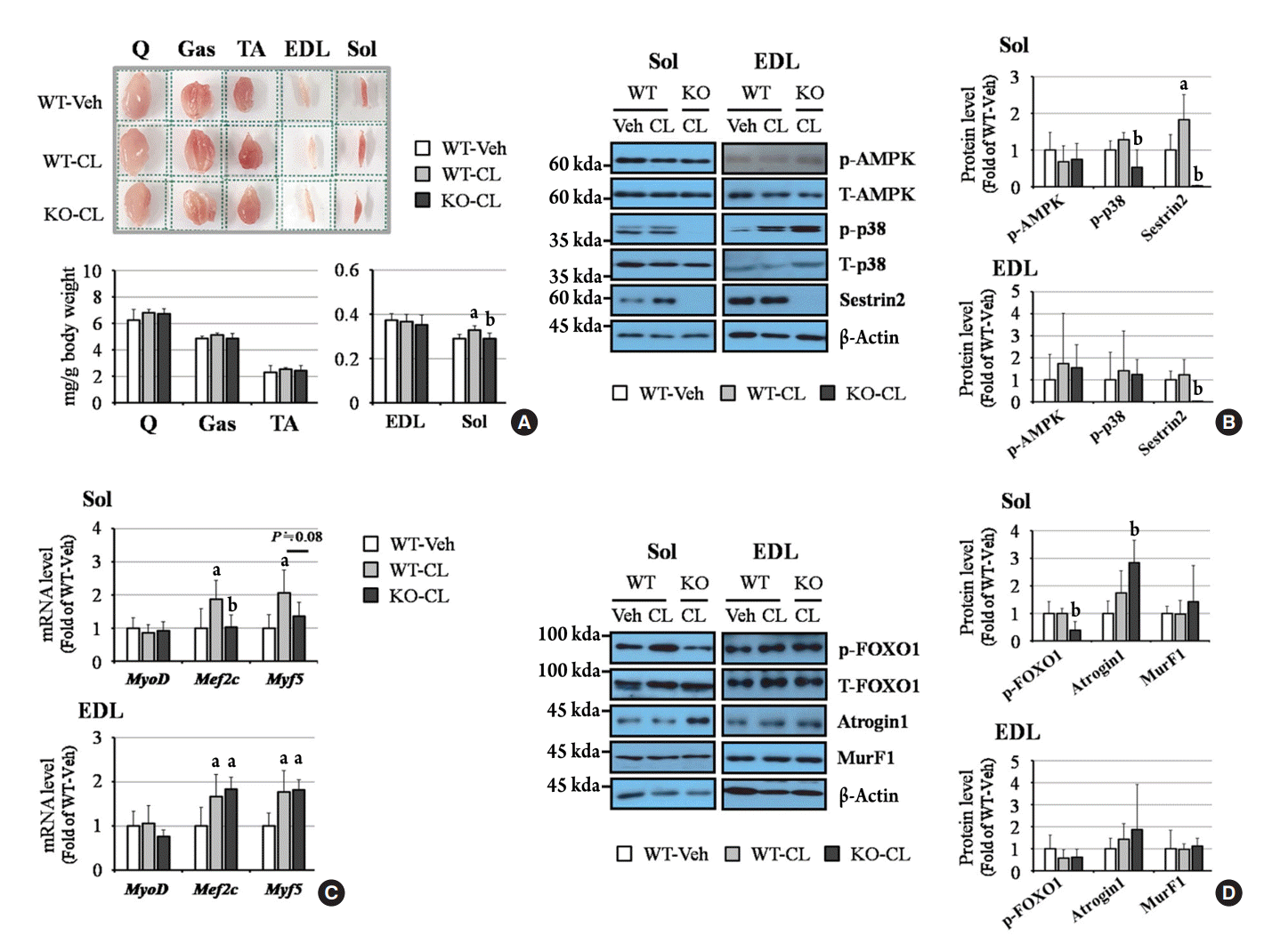
Fig. 2.
Sestrin2 deficiency regulates myogenic and atrophic-related events under CL 316,243 (CL) treatment in the soleus, but not in the extensor digitorum longus (EDL). (A) Muscle images and weight (per body weight). The side length of the dotted square is 1.2 cm. (B) Western blots of phosphorylated adenosine monophosphate-activated protein kinase (p-AMPK), p38, and sestrin2. (C) Quantitative polymerase chain reaction (PCR) of myoblast determination protein 1 (MyoD), myocyte enhancer factor 2C (Mef2c), and myogenic factor 5 (Myf5). (D) Western blot of phosphorylated forkhead box O1 (FOXO1), atrogin1, and muscle ring-finger protein-1 (MurF1). All graphs were obtained from six mice per group. Error bars represent mean±standard deviation. Q, quadriceps; TA, tibialis anterior; Gas, gastrocnemius; Sol, soleus. aP<0.05 vs. the wild-type (WT)-vehicle (Veh) group; bP<0.05 vs. the WT-CL group, as analyzed with analysis of variance (ANOVA) followed by the Tukey-Kramer test.
![]()
DISCUSSION
The β3AR-AMPK axis is a well-established signaling pathway for fat browning. AMPK deficiency was associated with decreased energy expenditure in CL-injected mice [11]. Given that sestrin2 is an endogenous AMPK regulator, we investigated whether sestrin2 affects β3AR-mediated fat browning. Our results showed that, in iWAT, β3AR-mediated AMPK phosphorylation, increase of brown fat markers, and mitochondrial biogenesis were blocked by sestrin2 deficiency. Furthermore, sestrin2 was linked to increased myogenic gene expression and decreased muscle atrophic proteins in the soleus of CL-treated mice.
UCP-1, which is expressed in brown fat but rarely detected in white fat [12], is located in the inner membrane of mitochondria and converts the proton gradient generated from oxidative phosphorylation to heat [13]. Therefore, increasing UCP-1 expression is considered to be an important strategy for promoting fat browning and energy expenditure. We showed that the abundance of UCP-1 mRNA and protein in iWAT was increased dramatically by CL treatment via a sestrin2-dependent manner in iWAT, but not in eWAT (Fig. 1C, D). Generally, the reactivity to CL depends on the type of fat. iWAT expresses higher levels of browning genes and mitochondrial activities than eWAT. Zuriaga et al. [14] and Gonzalez-Hurtado et al. [15] reported that CL treatment or cold stimulation induced the expression of UCP-1 and mitochondrial genes in iWAT, but not in eWAT. Our results also showed that, under CL treatment, the sestrin2-mediated reduction of LD size was related to UCP-1 expression and mitochondrial biogenesis in iWAT. However, the LD diameter of eWAT decreased after CL treatment, without beneficial sestrin2-mediated effects (Fig. 1C-G). Considering the elevation of HSL, a lipolysis marker, and serum FFA by CL treatment (Fig. 1H, I), we think that the triglycerides accumulated in eWAT were converted to FFAs and secreted into the blood, which led to LD size reduction in eWAT.
Although accelerated muscle atrophy and accompanying metabolic disturbances occur in the elderly [16], there is no definite treatment for SO. Sestrin2 could be a candidate for inhibiting SO since Segales et al. [9] reported that sestrin2 reduced FOXO1-atrogin1 signaling and restored muscle fiber size in hind limb-immobilized mice. Sestrin2 also improves insulin sensitivity [17], and its levels can be elevated by physical exercise in mice [18]. In our results, sestrin2 affected the CL-mediated induction of myogenic genes, Mef2c and Myf5, as well as attenuation of the FOXO1-atrogin1 pathway in the soleus, but not in the EDL (Fig. 2C, D). The soleus, which is composed mainly of type-1 and type-2a muscle fibers, has more mitochondria than the EDL, a type-2b and 2x fiber-dominant muscle [19]. Therefore, we infer that sestrin2 plays an important role in mitochondria-rich muscles such as the soleus. It is well-known that beneficial β2AR-mediated effects are mainly observed in type-2 fibers. Treatment with clenbuterol, a β2AR agonist, significantly increased the cross-sectional area of type-2 fibers in a β-arrestin-dependent manner and resulted in increases of the soleus and EDL mass. However, the beneficial effects mediated by the β2AR-β-arrestin axis were not detected in type-1 fibers [20]. Under β3AR activation, myogenic molecules increased in a sestrin2-dependent manner in the soleus, but in a sestrin2-independent manner in the EDL (Fig. 2C). These results show that βARs-mediated intracellular signaling is muscle type-dependent.
In conclusion, the present study found that sestrin2 elevated fat browning and myogenic molecules and reduced muscle atrophy-related molecules under CL treatment, suggesting that the β3AR-sestrin2 axis might be a good therapeutic target for the modulation of healthy body composition.
 XML Download
XML Download