

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Thyroid hormones have profound and widespread physiological effects, including regulation of metabolism and thermogenesis. Thyroid hormones also play key roles in growth and development which are phylogenetically diverse [1]. For example, over 100 years ago, it was shown that extracts of mammalian thyroid induce metamorphosis in tadpoles [2], whereas thyroidectomy prevents metamorphosis and arrests development [3]. Not long afterward, thyroidectomy was shown to disrupt the seasonality of breeding in ducks, songbirds, and sheep, and it is now known that thyroid hormone action on the mediobasal hypothalamus is essential to seasonal reproductive cycling across vertebrate species [4,5]. Key roles of thyroid hormones in human growth and development are also well established, as demonstrated by disorders such as congenital hypothyroidism and thyroid hormone resistance alpha [6,7]. The purpose of this review is to provide an overview of pituitary-thyroid axis function across the human lifespan.
Go to : 
PITUITARY-THYROID AXIS SETPOINTS
In a classic study by Andersen et al. [8], it was shown that in healthy individuals studied repeatedly over a year, circulating levels of thyrotropin (TSH) and thyroxine (T4) are tightly regulated, with much less variation observed within a given individual across time than between individuals. The physiological ranges for each individual (reflecting biological variation) are therefore narrower than the population-derived laboratory reference ranges. This gives rise to the key concept that each individual has set points (individual means) for TSH and T4, and what is normal for one individual may not be normal for another, even within conventional definitions of euthyroidism [9]. Notably, circulating TSH exists in several different isoforms with varying degrees of glycosylation, sialylation, and sulfonation which affect tissue availability and bioactivity [10,11]; this is not reflected in immunoreactive TSH concentrations determined by routine laboratory assays.
Setpoints for hypothalamo-pituitary-thyroid (HPT) axis function are probably established in utero. Evidence for this comes from individuals with congenital hypothyroidism in whom, despite early detection and treatment of hypothyroidism, the relationship between TSH and free T4 may be altered through childhood and adult life. In one study, at any given TSH concentration, children with congenital hypothyroidism had higher free T4 concentrations than children with autoimmune hypothyroidism [12]. Similarly, when individuals with congenital hypothyroidism were studied as adults, higher levothyroxine dosage and higher free T4 concentrations were required to achieve the same TSH concentration compared with patients with hypothyroidism from total thyroidectomy [13].
Pituitary-thyroid axis setpoints are to a large extent heritable traits. Estimates of heritability vary [14-18], but are up to 60% to 70% for each of TSH, free T4, and free triiodothyronine (T3) (Table 1). In recent years, there have been substantial advances in understanding the genetic architecture of pituitary-thyroid axis function [19]. This started with candidate gene studies, then genome wide association studies, initially using gene chips with relatively sparse coverage of the human genome in small populations, and more recently in large populations with more densely spaced genetic markers and high levels of imputation. This approach has been highly successful in identifying common genetic variants associated with thyroid function. In the most recent meta-analysis of 32 cohorts with a total sample size of more than 70,000 participants, 42 independent genetic loci were found to be associated with TSH, and 21% with free T4 [20]. Even with this large-scale effort, however, only 33% of genetic variance in TSH and 21% of variance in free T4 were accounted for. The “missing heritability” of thyroid function probably arises from multiple (as yet unidentified) genetic variants, each with small effects, requiring even larger studies for detection [21]. Rare variants (defined as a minor allele frequency <1%) are also likely to be important; although individually rare, these are collectively quite frequent and may have greater individual effect sizes than do common variants. In a whole genome sequencing study, several rare variants were identified as associated with thyroid function [22], and larger studies are required to build on this.
Epigenetic mechanisms such as DNA methylation, phosphorylation, acetylation and histone modification regulate gene expression without altering the DNA nucleotide sequence. This provides a potential link between environmental influences, gene expression and thyroid function [23]. Although non-permanent, epigenetic modifications can be transmitted to subsequent generations, and may account for some of the missing heritability of thyroid function [24]. Our group recently published the first epigenome-wide association study of thyroid function in healthy individuals, using methylation data from leukocyte DNA from two cohorts of adolescent Australians [25]. We identified two differentially methylated positions (DMPs) associated with TSH, none with free T4 and six with free T3, including DMPs in KLF9 and DOT1L, both genes which are known to be induced by T3. Further research is required to explore the relevance of these loci to pituitary-thyroid axis physiology and thyroid hormone action. DNA methylation shows marked changes across the lifespan [23,24], and is likely to be relevant to age-related changes in HPT axis function.
Go to :
ENVIRONMENTAL INFLUENCES
There are numerous environmental influences on HPT axis function. At the population level, the most important is iodine status [26]. The clinical impact of severe iodine deficiency is well-known, but even small differences in iodine status can affect the population distribution of TSH and alter the relationship between TSH and age [27-31]. Serum TSH, and to a lesser extent free T4 and free T3, is also affected by photoperiod and temperature, exhibiting circadian, seasonal and circannual variation [32-37]. The magnitude of these physiological effects can be clinically relevant: for example, the increase in TSH during winter can cause seasonal variation in the diagnosis of subclinical hypothyroidism [38]. Tobacco smoking affects thyroid function [39], such that current smokers have lower mean TSH concentrations than people who have never smoked. After smoking cessation, TSH levels gradually normalize over a prolonged period of 10 years or more [40].
Go to :
THYROID FUNCTION IN UTERO
Thyroid embryogenesis is largely complete by 7 weeks gestation, but terminal differentiation of the gland, characterized by follicle formation and thyroid hormone synthesis, does not occur until 10 to 12 weeks [41-44]. TSH, T4, and T3 are detectable in fetal circulation from 11 to 12 weeks onwards, but physiologically relevant thyroid hormone secretion does not occur until the second trimester. It is often stated that the fetal HPT axis is functionally mature by 18 to 20 weeks gestation [45,46], but in fact there is good evidence that maturation continues through the third trimester. These data come from studies utilizing cordocentesis for fetal blood sampling in utero or cord blood samples from fetuses born prematurely or undergoing termination of pregnancy [41,47-49]. The data are not completely consistent across studies, but it appears that fetal TSH levels are approximately 4 mU/L at 12 weeks, increase to 7 mU/L at 28 weeks, then remain stable until term. During the third trimester, fetal TSH is poorly sialylated and more bioactive than in older children and adults [50]; whether this is true earlier in gestation is not known. Serum total T4 concentrations in the fetus are low at 12 weeks gestation (~20 nmol/L), increase to ~70 nmol/L by 28 weeks then increase further to ~120 nmol/L at term, similar to levels in older children. Thyroxine-binding globulin (TBG) levels are low at 12 weeks, then increase progressively until term. Free T4 concentrations are also low (~2 pmol/L) at 12 weeks, increase to 15 to 20 pmol/L at 28 weeks, then plateau until term, influenced by the increase in TBG. Fetal total T3 levels are barely detectable at 12 weeks gestation, increase to ~0.4 nmol/L by 28 weeks and increase further to ~0.9 nmol/L at term, still lower than in older children.
Prior to maturation of the fetal thyroid, transplacental passage of maternal T4 occurs and plays an important role in fetal brain development. Transplacental transfer of T4 continues until fetal free T4 concentrations approximate maternal free T4 levels at ~28 weeks gestation [51]; in fetuses with congenital hypothyroidism, transplacental passage of T4 continues until term [52].
Go to :
EARLY LIFE AND CHILDHOOD
Immediately after birth, there is a surge in TSH secretion which peaks at around 30 minutes postnatally, with levels as high as 60 to 80 mU/L. This is followed by an increase in T4 which in turn peaks during the first day of life, when free T4 levels which can be twice as high as in older children or adults [49,53]. TSH and free T4 levels fall during the first month of life, but can still be above levels seen in adults [49,54,55]. Circulating T3 concentrations are low at birth, increase during the first week of life, and remain elevated across the first neonatal month. Studies of babies who were warmed or cooled in the first few hours and days of life suggest that high TSH levels during the first month of life are a response to the temperature drop between the intrauterine and extrauterine environments, although this may not be true of the early TSH surge immediately after birth, which appears to be temperature-independent [53]. In premature infants, the neonatal TSH surge in the first day of life is attenuated compared with term infants; the T4 surge is also attenuated in infants born at 28 to 34 weeks gestation and is absent in severely premature babies born at 23 to 27 weeks [49,56]. This, along with other factors, predisposes to hypothyroxinemia in premature infants [44], and provides further evidence that the fetal pituitary-thyroid axis is not fully mature until late gestation.
After the first month of life, TSH levels remain somewhat higher than in adults, gradually falling and plateauing at 6 to 9 months of age, then remaining largely stable through the rest of childhood, accompanied by a gradual fall in free T4 concentrations across this period [54]. The predominant TSH isoforms in children up to 18 months of age have low sialylation and N-glycosylation, resulting in high bioactivity, after which TSH glycobiology becomes similar to that seen in adults [57]. Circulating T3 levels peak at 4 to 5 months of age, then gradually decline. At all ages, mean TSH levels are slightly higher in boys than girls.
Reference ranges for TSH, free T4, and free T3 in children are typically wider than in adults, but are not well-standardized or harmonized. In a recent review, Onsesveren et al. [58] reported considerable variation: for example, for children aged 7 days to 3 months, reference interval lower limits for TSH ranged from 0.16 to 1.80 mU/L and upper limits from 4.38 to 12.56 mU/L; for 5 to 10 years of age the lower limits were 0.48 to 1.30 mU/L and upper limits 3.36 to 5.66 mU/L. Likely contributors to this variation include differences between studies in assay methods and statistical analysis, as well as iodine status, ethnicity, and anthropometric factors.
Children who develop hypothyroidism require higher doses of levothyroxine per kilogram (kg) of body weight than do adults, approximating 10 to 15 µg/kg/day in neonates, 4 to 6 µg/kg/day from age 1 to 5 years, and 2 to 4 µg/kg/day in older children and adolescents, whereas adults typically require 1.6 to 2.0 µg/kg/day [59]. This indicates the high level of metabolic and secretory activity of the thyroid during childhood.
Go to :
ADOLESCENCE
Adolescence is an important time of growth and development in which the thyroid plays a key role. We recently published the first detailed longitudinal analysis of thyroid function in an adolescent cohort [60], finding complex, sexually dimorphic patterns of thyroid function tests (Fig. 1). TSH was consistently higher in males than females with stable values from age 12 to 14 years, then an increase from age 14 to 16 in both sexes. In girls, free T4 increased from age 12 to 14 while remaining unchanged in boys, then increased from age 14 to 16 in both sexes. Free T3 was higher at all time points in boys than girls, and trajectories showed a marked sex difference. In girls, there was minimal change in free T3 from age 12 to 14, then a sharp decline to age 16, whereas in boys free T3 increased from age 12 to 14 then declined by age 16. These data are broadly consistent with limited data from other longitudinal studies [61,62] and cross-sectional studies [63]. The physiological basis for these changes is not fully understood. The sex differences (particularly for free T3) are partly explained by the earlier age of puberty in girls than boys, and the reciprocal changes in free T4 and free T3 concentrations suggest altered activity of one or more of the iodothyronine deiodinases, probably as a result of increased secretion of sex hormones, growth hormone/insulin growth factor 1 and other factors during puberty.
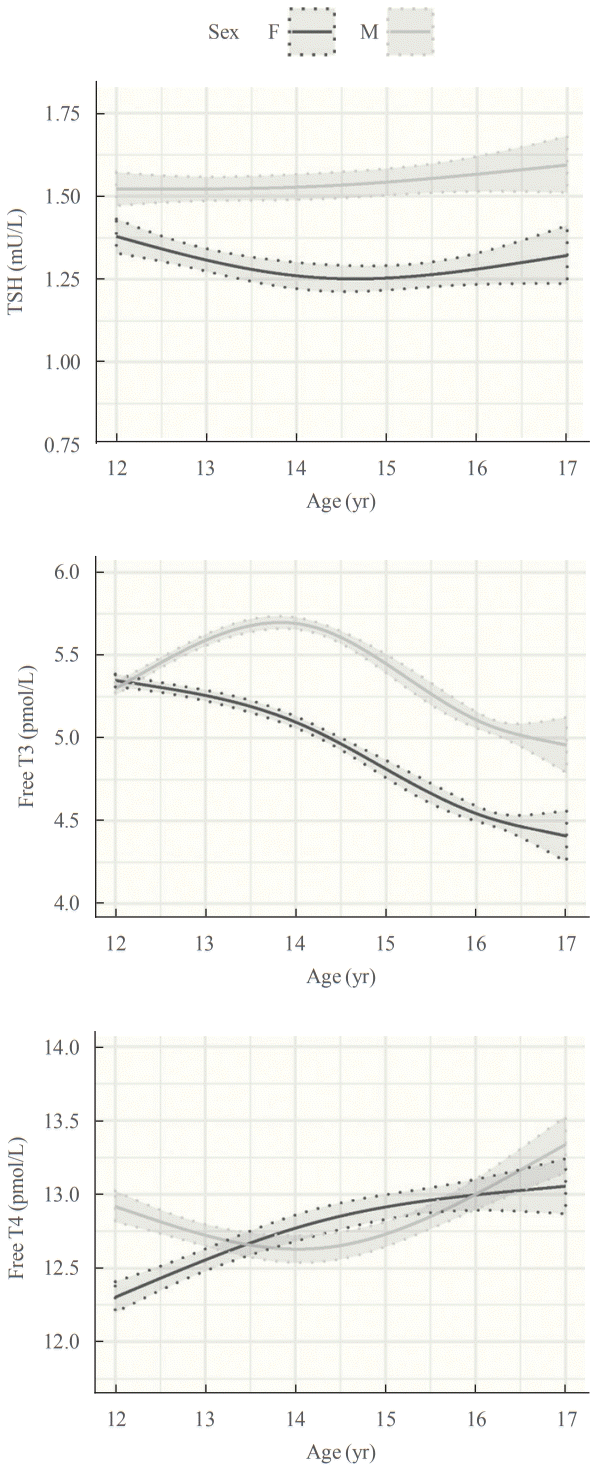 | Fig. 1.Longitudinal changes in thyrotropin (TSH), free triiodothyronine (T3), and free thyroxine (T4) in adolescent males and females. Results are visualized using generalized additive models fitted to the unadjusted data. Mean values and 95% confidence bands are illustrated by solid and dotted lines respectively. Reproduced from Campbell et al. [60], with permission from Oxford University Press.
|
These results are clinically relevant, because reference intervals for thyroid function tests clearly differ between adolescents and adults. For TSH and free T4, the differences were relatively minor, reflecting narrower distributions in teenagers than adults. For free T3, however, there is an upward shift in concentrations in adolescents than adults, most evident in 12-year-old girls and 14-year-old boys. In these age groups, use of adult reference ranges would misclassify 35% of girls and 58% of boys as having abnormal free T3 levels.
Go to :
THYROID FUNCTION IN ADULT LIFE
In healthy, older adults without thyroid disease, it was previously thought there was an age-related reduction in TSH levels [64,65], but this was based largely on studies from iodine-deficient populations after exclusion of individuals with subclinical hypothyroidism, which biased the results. In studies of iodinesufficient populations, including the National Health and Nutrition Examination Survey in the United States (NHANES III), it was shown convincingly that in healthy individuals with no evidence of thyroid disease, there is in fact a progressive increase in TSH concentrations with increasing age which is apparent from the third decade of life [66,67]. A limitation of NHANES III was the cross-sectional design, but an age-related increase in TSH secretion was subsequently confirmed in two longitudinal studies. In a reference group of Busselton Health Study participants with no evidence of thyroid disease studied on two occasions, 13 years apart (mean age at baseline 45.5 years), there was a significant increase in mean TSH concentrations with no change in mean free T4 levels [68]. In a second longitudinal study of older people (mean age 72 at baseline), also studied on two occasions, 13 years apart, there was an identical increase in mean TSH in a disease-free subcohort, accompanied by a small increase in mean free T4 [69]. These results were not replicated in a third study (mean age at baseline 65 years), in which mean TSH did not change significantly over time [70], probably because of the shorter duration of follow-up (6.5 years).
Further insight into the age-related increase in TSH comes from examining the relationship between TSH and free T4. A key feature of the TSH-free T4 relationship is that small changes in circulating free T4 concentrations result in relatively large changes in TSH. This accounts for the high community prevalence of subclinical thyroid disease, since TSH is frequently out of range accompanied by normal free T4 rather than the other way around, and underpins the “TSH-first” approach in screening for thyroid disease [71,72]. It was previously thought that there was a simple inverse log-linear relationship between TSH and free T4 [73-75], but it is now known that the relationship between log TSH and free T4 is complex and non-linear [76,77]. In an analysis of over 120,000 individuals from a laboratory database [72], we reported that the TSH-free T4 relationship differed across age groups, such that at any given free T4 concentration within the reference range, older people had higher TSH concentrations than younger people (Fig. 2), suggesting an age-related alteration in TSH setpoint.
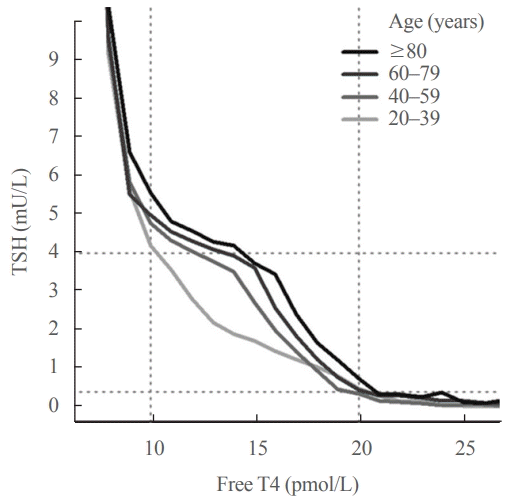 | Fig. 2.Thyrotropin (TSH) and free thyroxine (T4) relationship analyzed by age in 120,403 patients who were not taking thyroxine treatment. The median TSH for each free T4 integer value (in pmol/ L) was calculated, then plotted as 20-year age bands in adults. The youngest age band (1 to 19 years) is not depicted because there were insufficient data to calculate median TSH for each free T4 value reliably; for clarity, interquartile ranges are not shown. Dotted horizontal and vertical lines mark the TSH reference range (0.4 to 4.0 mU/L) and free T4 reference range (10 to 20 pmol/L), respectively. Reproduced from Hadlow et al. [72], with permission from Oxford University Press.
|
The mechanism for the age-related in TSH increase with aging is uncertain, and there are several possibilities. Firstly, it is possible (but largely untested) that TSH isoforms change with aging, resulting in reduced TSH bioactivity in older adults [11,78]. Secondly, there could be attenuation of the negative feedback effect of free T4 on TSH secretion, for example by reduced type 2 iodothyronine deiodinase activity in the pituitary. Thirdly, there could be a reduction in thyrocyte responsiveness to TSH, requiring higher TSH concentrations to maintain the same free T4 level. More research is required to address this question.
Go to :
IMPLICATIONS OF AGE-RELATED TSH INCREASE
The age-related increase in TSH, occurring without an accompanying fall in free T4 has important clinical implications. Firstly, it raises the question of whether age-related reference ranges for TSH should be applied to older adults. Following the publication of NHANES III data discussed above, this was advocated [79], but has not been widely adopted. One reason for this may be that in the early 2000s, there was robust debate about the TSH reference range in general, and in particular whether the TSH reference upper limit should be reduced from approximately 4–4.5 to 2.5 or 3.0 mU/L [80,81]. This debate is now largely settled, in favor of a TSH upper limit remaining at 4.0 to 4.5 mU/L [29,82], but may have overshadowed the important question of age-related ranges.
Table 2 shows examples of age-related reference ranges from selected studies in the literature, using young adults aged under 30 years as the comparator group [31,68,83-88]. A consistent feature is that the upper limit for TSH (defined as the 97.5th centile) is substantially higher in older age groups, being 0.5 to 2.7 mU/L greater than for young adults, whereas the lower limit of the reference range is not affected by age. The absolute values for TSH reference range limits differ between studies, reflecting methodological differences (e.g., definition of diseasefree groups, availability of thyroid antibody measurements) as well as preanalytic and analytic factors. These include environmental factors (especially iodine status and smoking, and to a lesser extent sex, season and temperature), ethnicity (which is known to impact TSH reference ranges [67,79,83]) and analytic factors, as TSH assays are poorly harmonized, with inter-assay differences of up to 1 mU/L at high-normal TSH concentrations [84,89]. Ideally, therefore, TSH reference ranges should be generated by laboratories from local population-based data where possible.
Table 2.
Age-Related Reference Ranges for Thyrotropin in Older People, with Young Adults as the Comparator Group
| Study | Location | Number | Assay |
TSH, mU/L |
|||
|---|---|---|---|---|---|---|---|
| Age 20–30 years | Age 60–70 years | Age 70–80 years | Age 80–90 years | ||||
| Boucai et al. (2011) [83] | USA | 13,296 | Nichols | 0.40–3.60 | 0.46–4.70 | 0.47–5.60 | 0.44–6.30 |
| Kahapola-Arachchige et al. (2012) [84] | Australia | 148,938 | Siemens Centaur | 0.49–3.67 | 0.52–4.32 | 0.48–4.52 | 0.47–4.9 |
| Bremner et al. (2012) [68] | Australia | 1,751 | Immulite | 0.51–3.54 | 3.48–4.70 | 0.52–5.28a | - |
| Vadiveloo et al. (2013) [85] | Scotland | 153,127 | Roche | 0.52–4.15 | 0.48–4.59 | 0.40–4.96 | 0.36–5.49 |
| Farrell et al. (2017) [86] | Australia | 604,194 | Siemens Centaur | 0.53–3.86 | 0.52–4.43 | 0.55–4.66 | 0.52–4.89 |
| Park et al. (2018) [31] | Korea | 5,987 | Roche | 0.67–6.05 | 0.56–7.77 | 0.42–6.68a | - |
| Mokhtar (2020) [87]b | Algeria | 8,838 | Abbott Architect | 0.46–3.90 | 0.42–5.10 | 0.36–5.30a | - |
| Raverot et al. (2020) [88] | France | 156,025 | Abbott Architect | 0.31–4.37 | 0.24–4.72 | 0.24–4.88 | 0.25–4.92 |
Data are from selected studies of populations thought to be free of thyroid disease (disease-free or reference populations) [31,68,83-88]. In some studies, sex-specific reference intervals were provided for men and women which have been combined to a single range for the purposes of this table. Young adults are shown as age 20 to 30 years, but in some studies data are for age 18 to 30 years.
![]()
Application of age-related reference ranges for TSH would reduce the number of older people regarded as abnormal on the basis of a mildly elevated TSH. This would be desirable, as the natural history of TSH elevations in older people appears favorable. TSH can increase as a result of nonthyroidal systemic illness, transient thyroiditis or simply biological variation, and often returns to the reference range on repeat testing [90]. For that reason, it is recommended that subclinical hypothyroidism be confirmed by repeat testing before levothyroxine treatment is started, but in clinical practice, patients are frequently started on treatment after a single measurement of TSH [91], and many continue treatment lifelong which may be unnecessary [92] and, if not adequately monitored, potentially harmful [93,94].
Even when confirmed on repeat testing, mild subclinical hypothyroidism in older people appears to have no health impact. A recent combined analysis of data from participants aged ≥80 years or more in two randomized controlled trials reported no benefit of levothyroxine treatment for subclinical hypothyroidism [95]. This is consistent with the full results of one of the included trials, which found no symptomatic treatment benefit in people aged ≥65 years or more [96]. Observational studies suggest that mild TSH elevation in older people does not predict adverse health outcomes. In fact, higher TSH is associated with greater life expectancy, including extreme longevity [69,78,97-101]. Although subclinical hypothyroidism is a risk factor for cardiovascular disease, a meta-analysis of individual participant data from 11 cohort studies found that this only applied to TSH levels ≥7.0 mU/L, with no increased risk associated with TSH 4.5 to 7.0 mU/L [102]. More recently, a study level meta-analysis of 35 cohort studies found that subclinical hypothyroidism was a predictor of mortality and cardiovascular outcomes only in cohorts which were younger (median age <65 years) and/or had high predicted cardiovascular risk at baseline [103]. In an informative community-based cohort study, stratified analysis showed that TSH in the highest quartile of subclinical hypothyroidism (TSH >6.57 mU/L) was a predictor of cardiovascular mortality and events (whereas TSH 4.0 to 6.57 mU/L was not), and that within this subgroup, participants aged ≥65 years were not at increased risk unless they had pre-existing cardiovascular risk factors [104]. Finally, in a large observational study from the United Kingdom, levothyroxine treatment of people with TSH between 5 and 10 mU/L was associated with reduced cardiovascular risk and overall mortality in patients aged 40 to 70 years (compared with people left untreated), but not in those aged over 70 years [105].
These data provide support a hypothesis advanced some years ago that subclinical hypothyroidism is a cardiovascular risk factor from middle age up to 65 or 70 years of age, but not in older people, whereas in the very elderly (over 85 years) it may be associated with prolonged survival [106,107]. In light of this, several clinical guidelines now recommend against routine levothyroxine treatment in older people with mild subclinical hypothyroidism [108-110]. In a recent editorial, Cappola [111] suggested that the upper limit of the TSH reference range be extended to 7 mU/L for people over the age of 80. This would probably reduce unnecessary levothyroxine prescribing for older people, but this dichotomous approach is less physiological and intellectually less appealing than applying graduated or continuous age-related reference ranges to all adults.
Go to :
CONCLUSIONS
This review highlights the considerable changes which occur physiologically in HPT axis function across the lifespan. Setpoints for thyroid function are to a large degree heritable and probably established in utero, but much of the underlying genetic architecture remains undescribed, and an understanding of epigenetic influences is only beginning to emerge. There are complex, dynamic changes in thyroid function tests across childhood and adolescence and also in adulthood, such that in older people TSH increases with aging without an accompanying fall in free T4. Age-related reference ranges for thyroid function tests should be routinely used for children and adolescents. There is a strong case for implementing age-related reference ranges for TSH in adults to prevent inappropriate diagnosis of subclinical hypothyroidism in older people and to discourage unnecessary levothyroxine prescribing.
Go to :
 XML Download
XML Download