

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
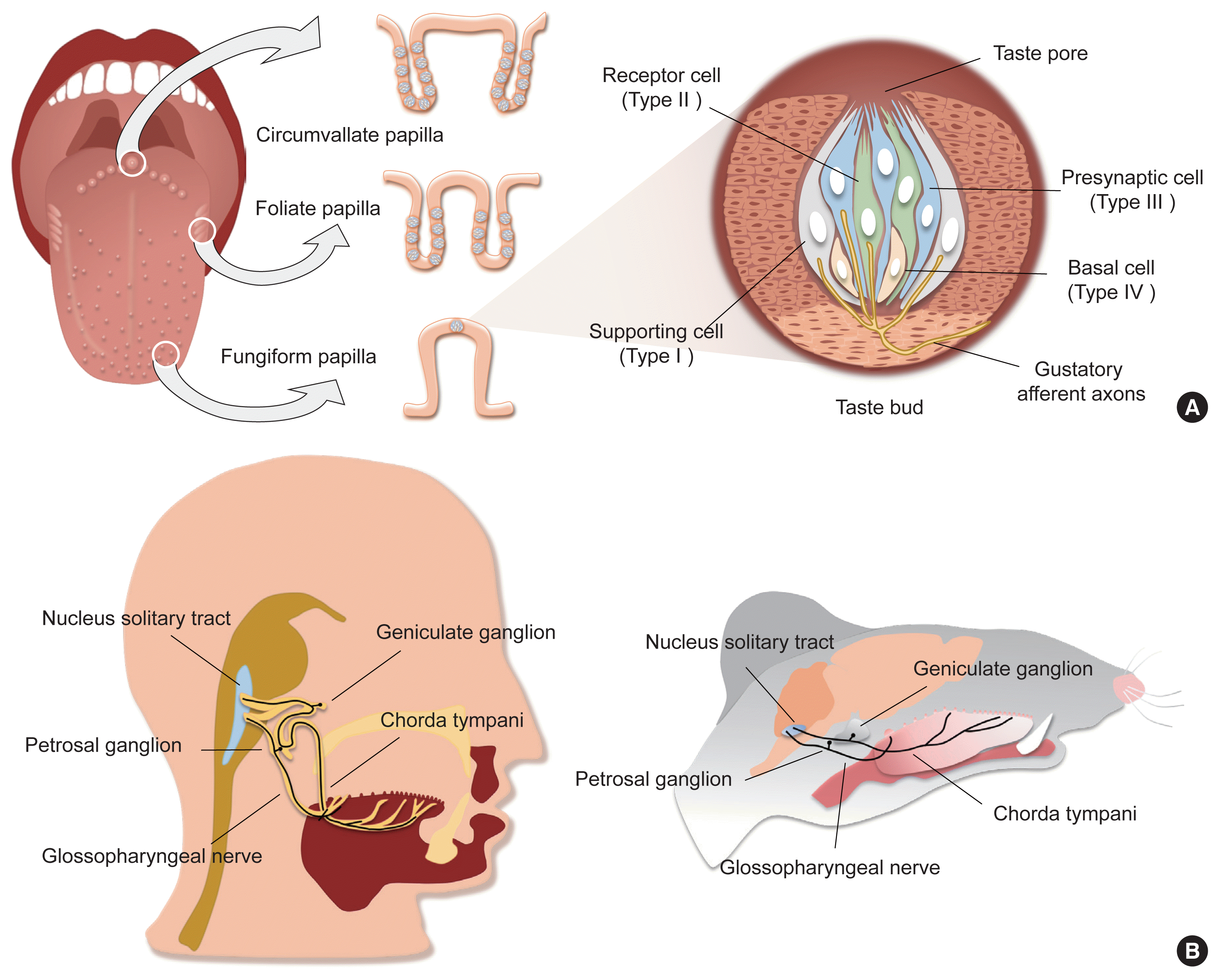
Taste is one of the specialized chemical senses that detects exogenous chemicals, and arises from contact of non-volatile chemicals dissolved in the saliva and mucus in the oral cavity with taste buds, the primary taste organs. Taste buds are located on the fungiform papilla (FuP), foliate papilla (FoP), and circumvallate papilla (CVP), but not on the filiform papilla, among the four types of papillary structures on the surface of the tongue (Fig. 1A). In addition, they are abundantly embedded in the soft palatal mucosa as well as the pharynx, and to a lesser extent the larynx, the epiglottis, and the bronchus. FuP-located taste buds are innervated by the chorda tympani nerve, a branch of the facial nerve, whereas FoP- and CVP-located taste buds are innervated by the glossopharyngeal nerve. The chorda tympani and glossopharyngeal nerve, whose neuronal cell bodies are located at the geniculate and petrosal ganglion, respectively, project their axons to the nucleus solitary tract, relaying taste information into the central nervous system (Fig. 1B).
A taste bud is composed of 50 to 100 epithelial cell aggregates arranged into a shape of an onion bulb, and is surrounded by lingual keratinocytes, which are morphologically distinct (Fig. 1A). Early ultrastructural analyses distinguished intragemmal—within any budlike or bulblike body—cells into four types of taste bud cells according to their morphology [1,2]. This classification is widely used today as it accurately reflects the gene expression patterns and physiological characteristics of each type of taste bud cell. Type I cells, the most abundant cell type (accounting for half of the total intragemmal cell population), have irregular and indented margins and wrap other types of cells at the periphery of taste buds. As they are reminiscent of glial cells surrounding neurons in the brain, they are referred to as “glia-like cells.” Type II cells, which show a spindle shape with a large, ovoid nucleus, are known as “receptor cells” due to their cellular responses to sweet, umami, or bitter stimuli via G-protein coupled receptor (GPCR) pathways. Type III cells, which accumulate synaptic vesicles toward nearby innervating neurons, are referred to as “presynaptic cells.” At the base of taste buds, type IV cells arise from perigemmal stem/progenitor cells, and differentiate into other types of intragemmal cells.
In this review, we introduce recent advances in understanding peripheral taste decoding, including (1) the discovery of non-canonical chemical synapses in type II cells, (2) the unexpected participation of type II cells in sensing appetitive salty taste, (3) the molecular mechanism of sour taste and its limitations, and (4) the dedicated responses and molecular heterogeneity of primary gustatory neurons.
DISCOVERY OF NON-CANONICAL CHEMICAL SYNAPSES IN TYPE II CELLS
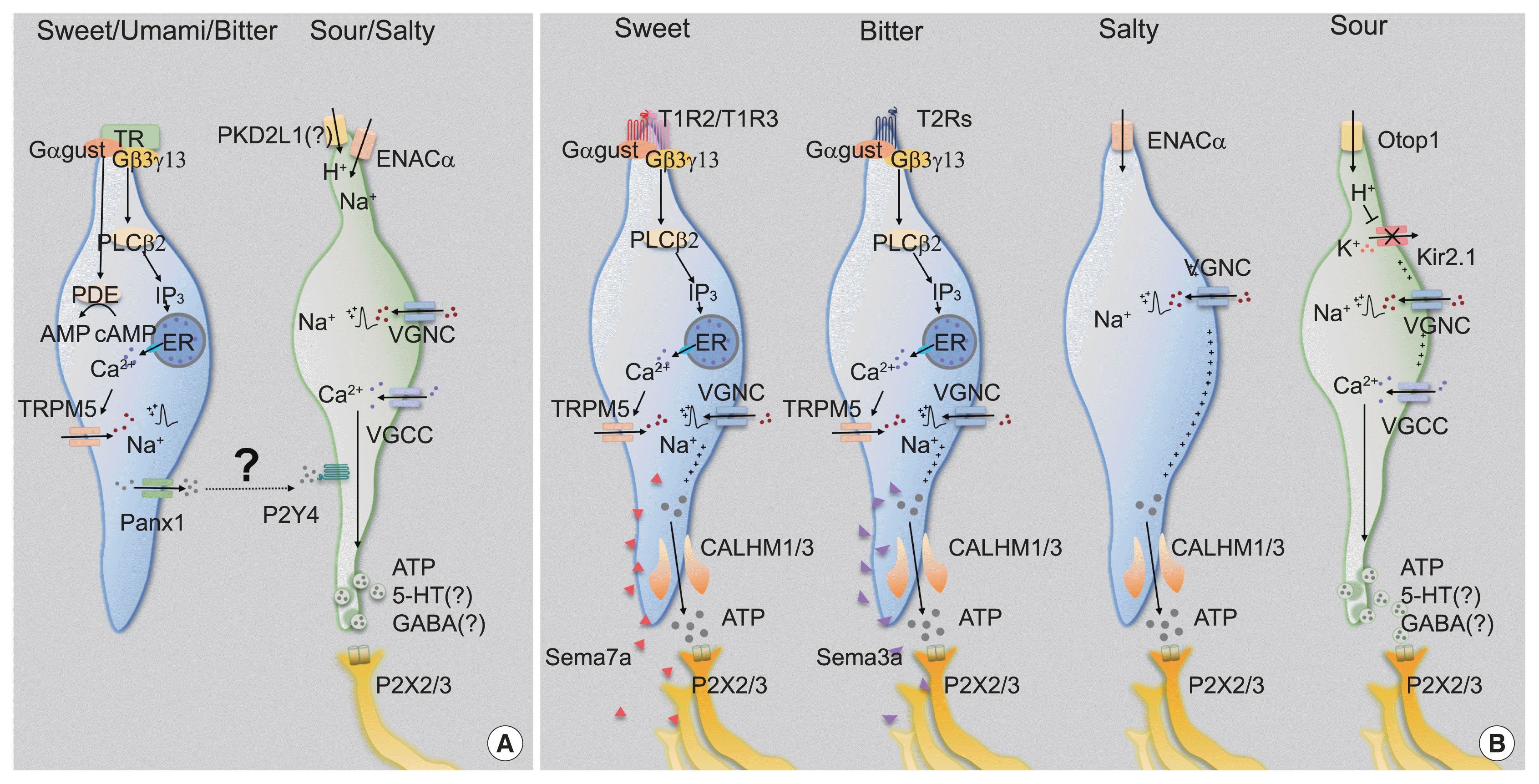
Type II cells detect either of sweet, umami, or bitter tastants via taste receptors, the subfamily of GPCR superfamily [3–6]. Expression of each type of taste receptor differs in a taste modality-specific manner. The sweet or umami taste receptor cell (TRC) only expresses T1R3, the co-receptor for sweet and umami tastes, forming a heterodimer with T1R2 or T1R1, respectively [3,4,6]. In contrast, the single bitter TRC expresses multiple bitter taste receptors (T2Rs) among the 25 and 35 T2Rs in human and mice, respectively, but T2R seems to function alone [7]. Taste receptors initiate intracellular signal transduction by stimulating gustducin (Ggust). Ggust is a taste-specific Gα protein with amino acid sequences similar to Gi [7], which simultaneously increases intracellular Ca2+ and decreases cyclic adenosine monophosphate (cAMP) [8,9]. Using an early version of chemogenetics, a receptor activated solely by a synthetic ligand (RASSL), artificial manipulation of sweet and bitter TRCs induced attractive and aversive behaviors for spiradoline, respectively [6,10]. Thus, one could infer that intracellular cAMP reduction leads to activation in type II TRCs. Since most excitable cells are hyperpolarized by intracellular cAMP reduction, this unique characteristic of type II TRCs is remarkable. However, the mechanism through which cAMP reduction leads to cellular activity remains elusive [8].
In contrast, the role of intracellular Ca2+ signaling has been well established. The Gβ3,γ13 complex, which is coupled to Ggust, dissociates from it upon taste receptor stimulation [11]. The free Gβ3,γ13 complex activates phospholipase Cβ2 (PLCβ2), hydrolyzing phosphatidylinositol 4,5-bisphosphate into inositol trisphosphate (IP3) and diacylglycerol. IP3 opens IP3R3 on the endoplasmic reticulum membrane, increasing cytosolic Ca2+ levels [12]. As a Ca2+-activated Na+ channel, transient receptor potential M5 (TrpM5) induces depolarization [13,14], and finally generates action potentials in Type II TRCs (Fig. 2A).
There has been a consensus that type II cells function as the cellular sensors for tastants, but also a long debate regarding how sensory information is transmitted to primary gustatory neurons. The absence of intracellular synaptic structures such as synaptic vesicles in ultra-high resolution transmission electron micrography images [2,15] and synaptic molecules in immunohistochemistry [15] had been a strong evidence that type II cells transmit the sensory information they generate by releasing synaptic vesicles of type III cells via cell-cell communication systems such as gap junctions [16]. Nonetheless, this proposal presented an insufficient explanation for the crosstalk-less transmission of each sensation modality.
Among the several neurotransmitters released from taste buds [17,18], adenosine triphosphate (ATP) has been highlighted as the genuine neurotransmitter of type II cells because genetic ablation of extracellular purinergic receptor X2 (P2X2) and P2X3 simultaneously results in depleted nerve responses and behaviors to most modalities of taste stimuli [19]. This means that a specific neurotransmitter does not transmit each specific modal taste, implying the existence of a direct connection between taste cells and gustatory neurons.
The discovery of calcium homeostasis modulator 1/3 (CALHM1/3) has been the master key that solves the questions raised above. First, all type II cells express CALHM1/3 [20,21]. Second, genetic depletion of CALHM1 impairs nerve responses and preference behaviors toward sweet, bitter, and umami tastants. Finally, CALHM1/3 directly effluxes ATP upon taste stimulation [20,21]. Therefore, CALHM1/3 provides the mechanism of gustatory information transmission without synaptic vesicles, refuting the cell-cell communication model and ultimately suggesting a novel type of purinergic synapses without exocytosis (Fig. 2B).
RESPONSIBILITY OF TYPE II CELLS IN FuP FOR APPETITIVE SALTY TASTE
Usually, a low concentration of Na+ results in an appetitive salty taste, whereas a high concentration manifests as an aversive salty taste. Although the salt appetite induced by Na+ depletion makes this simple classification unclear [22], amiloride serves as a solid branching point because it blocks gustatory responses to NaCl under 100 mM, but not above 100 mM. Thus, the former is regarded as amiloride-sensitive salty taste, and the latter as amiloride-insensitive [23,24]. As amiloride is an inhibitor of epithelial sodium channel (ENAC), ENAC has been believed to mediate appetitive salty taste. An ENAC heterotrimeric complex composed of ENACα, β, and γ functions as a selective Na+ channel that passively permeates extracellular Na+ in the nephrons [25]. In CVP, ENACα is expressed in type III cells [23]. Moreover, conditional ablation of Scnn1a, an ENACα-encoding gene, in all mature taste cells resulted in diminished nerve responses and preferences for a low concentration of Na+ in an amiloride-dependent manner [23]. Therefore, ENACα has been suggested to be required for appetitive salty taste detection (Fig. 2A).
However, multiple groups have raised questions about the role played by ENAC. The generation of fluorescent tag-inserted knock-in mice for Scnn1β and Scnn1γ revealed that neither is co-expressed with ENACα in taste buds, thereby implying that appetitive salty taste perception is instead initiated by an unknown mechanism, not by the heterotrimeric ENAC complex [26]. Moreover, an independent electrophysiological approach revealed that an amiloride-sensitive current was not recorded in ENACα-expressing cells in CVP, but in FuP [27]. Surprisingly, the cells expressing ENACα in FuP were positive for CALHM1/3, the novel marker of type II cells. CALHM3 knockout (KO) mice lost amiloride-sensitive nerve responses and attractive behavior for salt, indicating that appetitive salty taste is mediated by type II cells and refuting the previous hypothesis of engagement of type III cells (Fig. 2B) [27]. However, others reported that CALHM1 KO mice, which had lost an additional component for purinergic transmission, had a normal ability to sense low concentrations of salt [28]. For the moment, more comprehensive evidence is needed to establish the paradigm of appetitive salty taste.
Cations other than Na+ are also detected as taste. One of the bitter receptors, taste 2 receptor member 7 has been reported to detect several metal ions, although in vivo evidence is still required [29,30]. Moreover, tongue slice imaging was presented to support that type II cells respond to Cl−, representing amiloride-insensitive salty taste, but this possibility requires further investigation [31].
SOUR SENSING MECHANISM OF TYPE III CELLS
A series of genetic experiments revealed that type III cells are required for sensing sour taste [32]. Polycyctic kidney disease 2-like 1 (PKD2L1), a specific molecular marker of type III cells, shows a distinctive expression pattern in double immunohistochemistry against type II cell markers [33]. PKD2L1-TeNT mice, which express tetanus toxin (TeNT) to silence synaptic transmission of type III cells, showed diminished nerve responses to sour stimuli [34]. Therefore, one might expect that type III cells promote aversive behaviors because sour stimuli evoke aversive behaviors and activate type III cells. However, it is interesting to note that the behavior of PKD2L1-TeNT mice in response to sour stimuli had not been reported until recently.
The Oka group recently reported the unexpected results that PKD2L1-TeNT mice displayed a normal level of aversive behavior against sour stimuli [35]. Rather, optogenetic activation of type III cells does not induce aversive, but attractive behaviors. Although the attractive behavior induced by optogenetics is suspected by other group, it is still agreed that artificial activation of type III cells does not induce aversion by itself [36]. This means that type III cell activity is unnecessary and insufficient to drive sour-induced aversive behaviors, despite type III cellular responses. It has subsequently been shown that trigeminal chemesthesis is additionally required to generate sour-induced aversion [37].
How do type III cells respond to acids? Initially, PKD2L1, the well-known marker of type III cells, had received attention as an acid sensor (Fig. 2A) [33], but this possibility was quickly refuted because PKD2L1 KO mice responded to acidic stimuli normally [38]. A recent transcriptomic comparison between type II and type III cells indicated that otopetrin 1 (Otop1) is the genuine sour sensor [39]. Otop1 gates protons from outside into the cytosol, mediates intracellular acidification to close Kir2.1 [40], and ultimately depolarizes type III cells (Fig. 2B) [41]. Although Otop1 KO mice lack both the electrophysiologic nerve responses and the ganglionic Ca2+ responses to sour stimuli, they still show normal acid-induced aversive behaviors, confirming that the sour taste itself is insufficient to generate aversion [37,41].
Type III cells also respond to carbonated water, pure water, and high salts [24,35,42]. The detection of both carbonated and pure water is an intrinsically identical phenomenon involving reduction of the pH of solvents. CO2 from carbonated water is dissolved into the solvent by carbonic anhydrase 4 expressed in type III cells, generating protons [42]. The ingestion of pure water removes bicarbonate from the saliva, transiently reducing pH [35]. A further investigation is required to establish the responses of type III cells to high salts [24].
Taken together, it is currently accepted that type III cells mainly respond to stimuli that change the pH or ionic balance of the oral cavity, but it is still debated whether the activity of type III cells promotes preference or avoidance by itself. This may stem from the possible heterogeneity of type III cells, but their scarcity has made it hard to unveil the truth.
DEDICATED RESPONSES OF THE GENICULATE GANGLION
Early single-fiber nerve recordings as well as recent in vivo Ca2+ imaging techniques have shown the dedicated responses of primary gustatory neurons to each taste modality [43–45], which implies the existence of selective connections between taste-responsive cells in taste buds and primary gustatory neurons. Therefore, taste-responsive cells for each modality require molecular machinery for exclusive synapses to the corresponding neurons.
Recently, the Zuker group demonstrated that several semaphorins (SEMAs), as a subcategory of cell adhesion molecules, are responsible for the precise innervation of gustatory neurons into taste buds in a modality-specific manner (Fig. 2B) [46]. SEMA expression differs depending on cellular modality type, with SEMA7A found in sweet taste cells and SEMA3A in bitter taste cells. Genetic depletion of each SEMA disrupts both taste-guided behaviors and the modality-specific responses of the ganglion. Moreover, ectopic expression of human SEMA7A in sour cells transmitted sour signals into a sweet-dedicated ganglion, confirming the role of the SEMA system in wiring epithelial cells to neurons. Given that SEMAs are secreted from taste bud cells, specific binding partners should be expressed in each type of neuron. These findings imply that primary gustatory neurons show considerable molecular heterogeneity.
Single-cell RNA sequencing (scRNAseq) enabled classification of entire population of neurons of the geniculate ganglion based on transcriptomic profiles [37,47]. Gustatory neurons are not only molecularly isolated from the somatosensory components of the auriculotemporal nerve [47], but are also divided into five classes corresponding to each taste modality [37]. Indeed, the Zuker group identified specific markers for each group: Spondin1 for sweet; Cdh4 for umami; Cdh13 for bitter; Penk for sour; and Egr2 for salty taste. They created KO mice and transgenic mice carrying Cre for each marker, observing behavioral defects and selective responses of the labeled ganglia, respectively [37]. Therefore, it was confirmed that the relay of information for each taste modality is genetically defined at the entrance of the neural network.
HORMONAL EFFECTS ON TASTE PERCEPTION
Hormones are usually defined as chemical signals secreted into the bloodstream which act on distant tissues for their specific regulation. In addition to the primary ‘endocrine’ organs, taste buds also had been reported to express metabolic hormones such as vasoactive intestinal peptide and serotonin [48,49]. In the early 21st century, more evidence had accumulated that taste cells express several metabolic hormones and their relevant receptors. Accordingly, the concept has emerged that hormones can act on taste cells and, as a result, can affect taste perception in autocrine and/or paracrine manner. Several types of metabolic hormones expressed in taste cells had been studied, including neuropeptide Y (NPY) and cholecystokinin (CCK) [50,51]. Similar to the diametrical roles of anorexigenic CCK and orexigenic NPY in energy metabolism [52], they also have opposite effects on potassium current in taste cells [53]. Resulting amplification of bitter taste perception against sweet taste perception are assumed not to pass over bitter-tasting toxins, which can be an example of regulation of taste decoding in a coordinated manner.
In the past decade, more hormones and/or their receptors have been discovered in taste cells, and their roles in taste decoding by modulating intracellular signaling have been delineated. Glucagon enhances sweet taste, which is similar to glucagon-like peptide-1, another cleavage product generated from a proglucagon peptide [54,55]. Ghrelin reduces salty and sour taste responsivity [56]. Angiotensin inhibits salty taste and enhances sweet taste sensitivities [57] while leptin selectively suppresses neural and behavioral responses to sweet taste [58]. Various research groups have tried to define the expression patterns and roles in taste perception of the hormones, which are well described in other review articles [59,60].
Conversely, regulation of synthesis and secretion of hormones from taste buds has been unclear yet. Considering that nutrients and energy status regulate synthesis and secretion of metabolic hormones in major endocrine organs, it is plausible that each tastant also might facilitate hormonal expression and/or secretion in taste cells. Furthermore, the taste buds derived-hormones might affect systemic metabolism. Cephalic phases insulin responses might be the good example to support to this [61,62]. However, the extent of taste bud derived-hormones affecting at systemic levels should be re-examined. Taste buds are too tiny and limited in their number to study with conventional biochemical research methods, thus novel tools including organoids would serve as the alternatives in this field.
CONCLUSIONS
Although the discovery of CALHM1/3, Scnn1a, Otop1, and SEMA broadened the current understanding of peripheral taste decoding, several questions still remain. Are there any unidentified taste modalities like fat and starch? How many types of intragemmal cells can be classified according to the transcriptomic profile? What is the molecular mechanism of the differentiation of taste cells that respond to specific taste modalities? What is the effect of each taste stimulus on regulating the expression of each hormone in taste cells? Do knowledges of taste decoding obtained from animal experiments have human relevance? High-throughput scRNAseq for taste buds would be expected to resolve transcriptomic heterogeneity issues, illuminating unexpected and novel molecular pathways, and eventually helping to regulate feeding behaviors and metabolism in humans.
 XML Download
XML Download