

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Diabetes mellitus is a metabolic disorder that is associated with an increased risk of several comorbidities, including cardiovascular disease (CVD). Atherosclerotic CVD, such as myocardial infarction or stroke, is the main cause of death (accounting for almost 80% of the death) in patients with type 2 diabetes [1]. Atherosclerosis is accompanied by vascular endothelial dysfunction, leading to hardening of the arteries and arterial occlusion, which is a fundamental process underlying CVD. The development of atherosclerosis is complex and influenced by glucose and lipid metabolism, inflammation, and oxidative stress [2,3].
The glucagon-like peptide 1 (GLP-1) receptor has been a treatment target for diabetes. GLP-1 analogues improve insulin sensitivity in the liver and adipose tissue, reducing macrophage infiltration and inhibiting inflammation [4–7]. GLP-1 has multiple biological functions related to the cardiovascular (CV) system [8]. The CV safety and benefits of GLP-1 analogues were reported in recent CV outcome trials [9–13]. However, these drugs, particularly at high-dose, exhibit side effects, such as nausea, vomiting, and other types of gastrointestinal discomfort [14].
The fibroblast growth factor 21 (FGF21), which is a member of the FGF superfamily, is expressed in several tissues, including the liver [15]. FGF21 has emerged as an important regulator of glucose and lipid metabolism [16,17]. Recent studies reported that FGF21 decreased hepatic steatosis, blood glucose, insulin, glucagon, and body weight, and improved insulin sensitivity in rodent models [18,19]. Treatment with FGF21 decreased the development and progression of atherosclerosis in an animal model [20]. Furthermore, FGF21 therapy decreased the concentrations of low-density lipoprotein cholesterol and alleviated insulin resistance in animal models of obesity [18,21]. However, the FGF21 protein has unfavorable pharmacokinetic properties, such as susceptibility to in vivo proteolytic degradation and a short half-life (<2 hours) [22].
To avoid the side effects of liraglutide and to overcome the shortcomings of FGF21, a low-dose combination therapy with a GLP-1 analogue and FGF21 might be a preferred strategy to prevent the cardiometabolic risk associated with diabetes, by possibly lowering glucose levels and reducing body fat and CV risk. Thus, we aimed to estimate the ability of a low-dose combination of liraglutide, which is a GLP-1 analogue, and FGF21 to mitigate the vascular complications of diabetes. Thus, the effect of the GLP-1 analogue/FGF21 combination therapy on vascular cell proliferation, inflammation, and atherosclerosis was explored in a high-fat diet-induced obese mouse model and in vascular smooth muscle cells (VSMCs). Finally, the mechanisms underlying the effects of the combination therapy were investigated using molecular experiments.
METHODS
Animals
Six-to-eight-week-old C57BL/6J male mice were obtained from Jackson Laboratories (Bar Harbor, ME, USA). All mice received a Western diet (40% fat and 43% carbohydrate) for 30 weeks and an atherogenic diet (4% cholesterol, 1% cholic acid, and 0.5% 2-thiouracil) for another 10 weeks, and were divided into four treatment groups: control (saline); liraglutide (0.3 mg/kg/day), as a GLP-1 analogue; FGF21 (LY 2405319; donated by Eli Lilly and Company, Indianapolis, IN, USA; 5 mg/kg/day); and low-dose combination treatment with liraglutide (0.1 mg/kg/day) and FGF21 (2.5 mg/kg/day) (n=6/group) for 6 weeks. The investigational products were administered intraperitoneally. Mice were housed in a temperature-controlled room and maintained under a 12/12-hour light/dark cycle. Mice were sacrificed at 55 weeks of age.
This study was approved by the Institutional Animal Care Committee, Seoul National University Bundang Hospital (SNUBH) (IACUC No. BA1710-233/083-01). The animal experiments were performed in compliance with the Guide for Experimental Animal Research of the Laboratory for Experimental Animal Research, Clinical Research Institute, SNUBH, South Korea, and conformed to the provisions of the Declaration of Helsinki (2013).
Atherosclerosis in the aorta
To measure the area of atherosclerotic lesions, whole aortas prepared using the en face method were stained with Oil-Red-O solution [23]. After perfusion fixation of the euthanized mice, the whole aorta was dissected out, opened longitudinally from the heart to the bifurcation of the iliac arteries, and stained with Oil-Red-O. Each specimen was evaluated in a blinded fashion. Images of the aorta were analyzed using Image-Pro Plus software (Media Cybernetics, Silver Spring, MD, USA), and results are presented as the area of atheromatous plaque lesion calculated as a percentage of the whole aortic area.
Weight, glucose, and biochemical parameters
During the treatment period, body weight was measured twice a week. Food and water intake were also measured every week throughout the treatment period. Fasting blood glucose concentrations were measured twice a week using a glucometer (Accu-Chek Inform II, Roche, Mannheim, Germany). Blood samples were collected from mice after 8 hours of fasting at the end of the treatment period. The levels of insulin, glucagon, leptin, ghrelin, and resistin were measured using a Multiplex Assay Kit (MMHMAG-44K, Lot. 3200934, Millipore, Billerica, MA, USA). Adiponectin concentrations were measured using enzyme-linked immunosorbent assay kits (MRP300, R&D system, Minneapolis, MN, USA) according to the manufacturer’s instructions [24].
Cell culture
Rat aortic smooth muscle cells were purchased from the American Type Culture Collection (ATCC, Manassas, VA, USA). Cells were maintained in Dulbecco’s Modified Eagle’s Medium (DMEM) with 10% fetal bovine serum (HyClone, Logan, UT, USA) in a humidified atmosphere of 5% CO2 and 95% air at 37°C. The medium was replaced every 2 days. Prior to the experiments, the cells were plated in 96-well plates at a density of approximately 1.5×104 cells per well for 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium (MTS) assay, or in 6-well plates at 5×106 cells per well for all other assays. For the experiments, the cells were incubated with the agents for 24 hours at 37°C. In each experiment, treatments were performed in triplicate.
Cell viability assay
Cells were cultured to 80% to 90% confluence and then starved in serum-free DMEM for 24 hours. The medium was replaced with fresh medium containing different concentrations of liraglutide and FGF21 in the presence of the platelet-derived growth factor (PDGF; 10 ng/mL). PDGF is a growth factor that is commonly used to induce the proliferation of vascular cells [25]. Cell viability was analyzed using the MTS cell proliferation assay (CellTiter 96 AQueous Cell Proliferation Assay Kit, Promega Corp., Madison, WI, USA), according to the manufacturer’s protocol. Briefly, the cells were cultured for 24 hours and 20 μL/well of the MTS solution was added to the samples in 100 μL of culture medium. The cells were subsequently incubated at 37°C for 4 hours and the absorbance was measured at 490 nm using a microplate reader.
Wound-healing migration assay
Cells were grown to 90% confluence and then subjected to scratching using a sterile pipette tip. The medium was replaced with fresh medium containing different concentrations of liraglutide and FGF21 in the presence of PDGF (10 ng/mL). The scratch wound was allowed to heal for 24 hours in the presence of the indicated chemicals. A phase-contrast microscope (Optika, Ponteranica, Italy) captured images of each sample at 0 and 24 hours, and the ability of VSMCs to migrate was evaluated by measuring the area of the scratch wound at both time points, using Image J software version 1.29x (National Institutes of Health, Bethesda, MD, USA).
Annexin V staining and flow cytometry
To determine the rate of apoptosis, the cells were incubated in culture medium containing liraglutide and FGF21 for 24 hours and stained with Annexin V/fluorescein isothiocyanate (FITC), according to the manufacturer’s protocol (Molecular Probes, Thermo Fisher Scientific, Inc., Waltham, MA, USA). Approximately 1×105 cells were harvested and washed with phosphate-buffered saline. Cells were then resuspended in 100 μL of Annexin V binding buffer (10 mM HEPES, 140 mM NaCl, and 2.5 mM CaCl2; pH 7.4), incubated with 5 μL of Annexin V/FITC for 15 minutes at room temperature, and counterstained with propidium iodide (PI; final concentration, 1 μg/mL) for 10 minutes at room temperature. After the incu-bation period, the cells were diluted with 190 μL of Annexin V binding buffer and analyzed by flow cytometry using a Becton Dickinson FACScan flow cytometer with the Cell Quest 3.1 software (BD Biosciences, Franklin Lakes, NJ, USA).
Reverse transcription quantitative polymerase chain reaction analysis
Total RNA was extracted from the cells according to the Promega Total RNA Isolation System manual. Reverse transcription quantitative polymerase chain reaction was performed on an ABI Prism 7500 Sequence Detection System (Applied Biosystems Inc., Foster City, CA, USA), according to the manufacturer’s instructions. SYBR Green (Toyobo Corp., Osaka, Japan) was used as a double stranded DNA-specific fluorescent dye. The specific primers used here were as follows: uncoupling protein-1 (UCP1) forward, 5′–ATCGGCCTCTACGACAACGTCAA–3′ and reverse, 5′–ACCAGTTCTGTGCAGTTGACCAG–3′; Akt1 forward, 5′–ACTCATTCCAGACCCACGAC–3′ and reverse, 5′–ATACACATCCTGCCACACGA–3′; and β actin forward, F 5′–CGTTGACATCCGTAAAGACCTC–3′ and reverse, 5′–TAGGAGCCAGGGCAGTAATCT–3′.
Western blot analysis
Cells (1×106/mL) were resuspended in a lysis buffer (Cell Signaling Technology Inc., Danvers, MA, USA) containing a protease inhibitor cocktail (cOmplete Mini Protease Inhibitor Tablet, Roche Diagnostics, Rotkreuz, Switzerland). The protein concentrations were measured in the supernatant using a Pierce BCA Protein Assay Kit (cat. no. 23225, Thermo Fisher Scientific Inc.). A total of 40 μg of protein was loaded per gel lane, separated via 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), and transferred onto nitrocellulose membranes. The membranes were blocked for 2 hours at room temperature with 5% skimmed milk in Tris-buffered saline with Tween 20 (TBST; 20 mM Tris, 500 nM NaCl, and 0.1% Tween 20), followed by incubation with primary antibodies against cleaved caspase-3, matric mentalloproteinase-9 (MMP-9), intercellular adhesion molecule-1 (ICAM-1), monocyte chemoattractant protein-1 (MCP-1), phosphorylated Akt (p-Akt), phosphorylated extracellular signal-regulated kinase (p-ERK), phosphorylated p38 (p-p38), and phosphorylate JNK (p-JNK) (all 1:1,000; Cell Signaling Technology, Boston, MA, USA), and β-actin (1:5,000; Santa Cruz Biotechnology, Santa Cruz, CA, USA) overnight at 4°C. After three washes with TBST, the membranes were incubated with secondary horseradish peroxidase conjugated anti immunoglobulin G antibodies (1:5,000; Invitrogen; Thermo Fisher Scientific Inc.) for 2 hours at room temperature and visualized using a Pierce enhanced chemiluminescence substrate (Thermo Fisher Scientific Inc.). Densitometric quantification of the protein bands was achieved using the ImageJ software version 1.29x (National Institutes of Health).
Statistical analyses
Quantitative data are presented as mean±standard deviation for animal studies and mean±standard error for cell studies. Differences between mean values were compared statistically by two-tailed Student’s t test or one-way analysis of variance followed by Tukey’s post hoc comparison. Statistical analyses were performed using the SPSS Windows software version 22.0 (IBM Corp., Armonk, NY, USA). Significance was set at P<0.05. All experiments were performed at least three times.
RESULTS
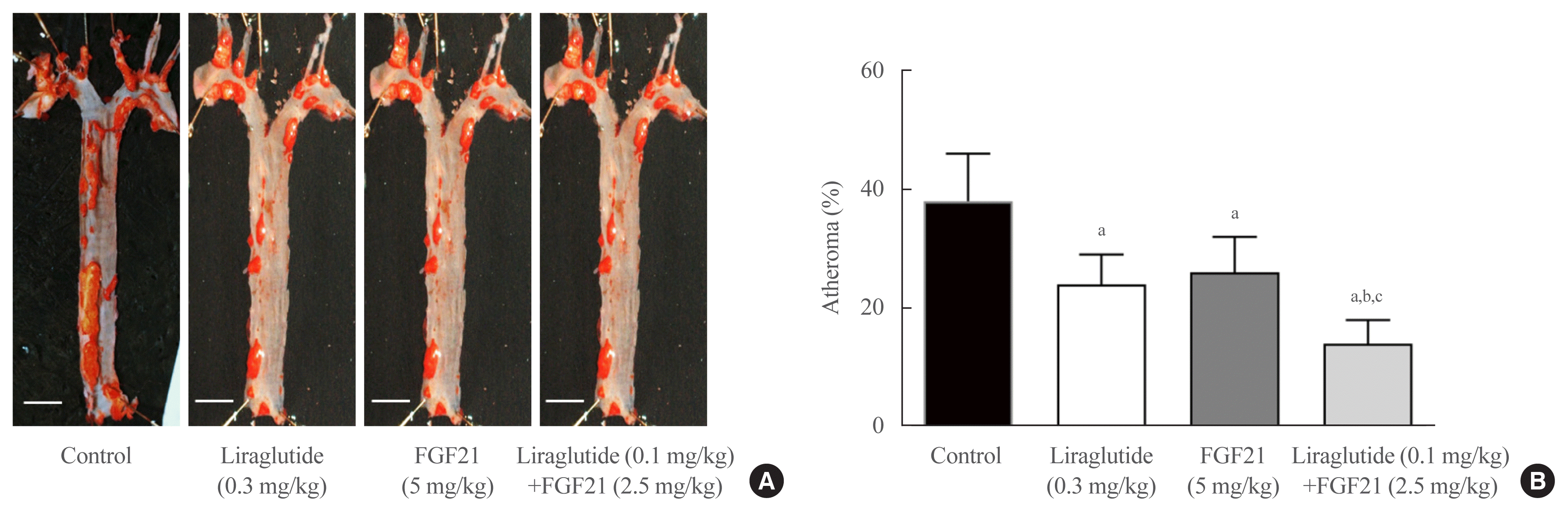
Effects of a 6-week treatment with liraglutide, FGF21, or their combination on atheroma in the aorta
Plaque accumulation in the aorta of C57BL/6J male mice is shown in Fig. 1. Quantification showed that the plaque areas were significantly smaller in the groups treated with liraglutide, with FGF21, or their combination than in controls (all P<0.05). Moreover, the low-dose combination therapy of liraglutide (0.1 mg/kg) and FGF21 (2.5 mg/kg) showed significantly lower atheromatous plaque formation in the aorta than either liraglutide alone or FGF21 alone (both P<0.05) (Fig. 1).
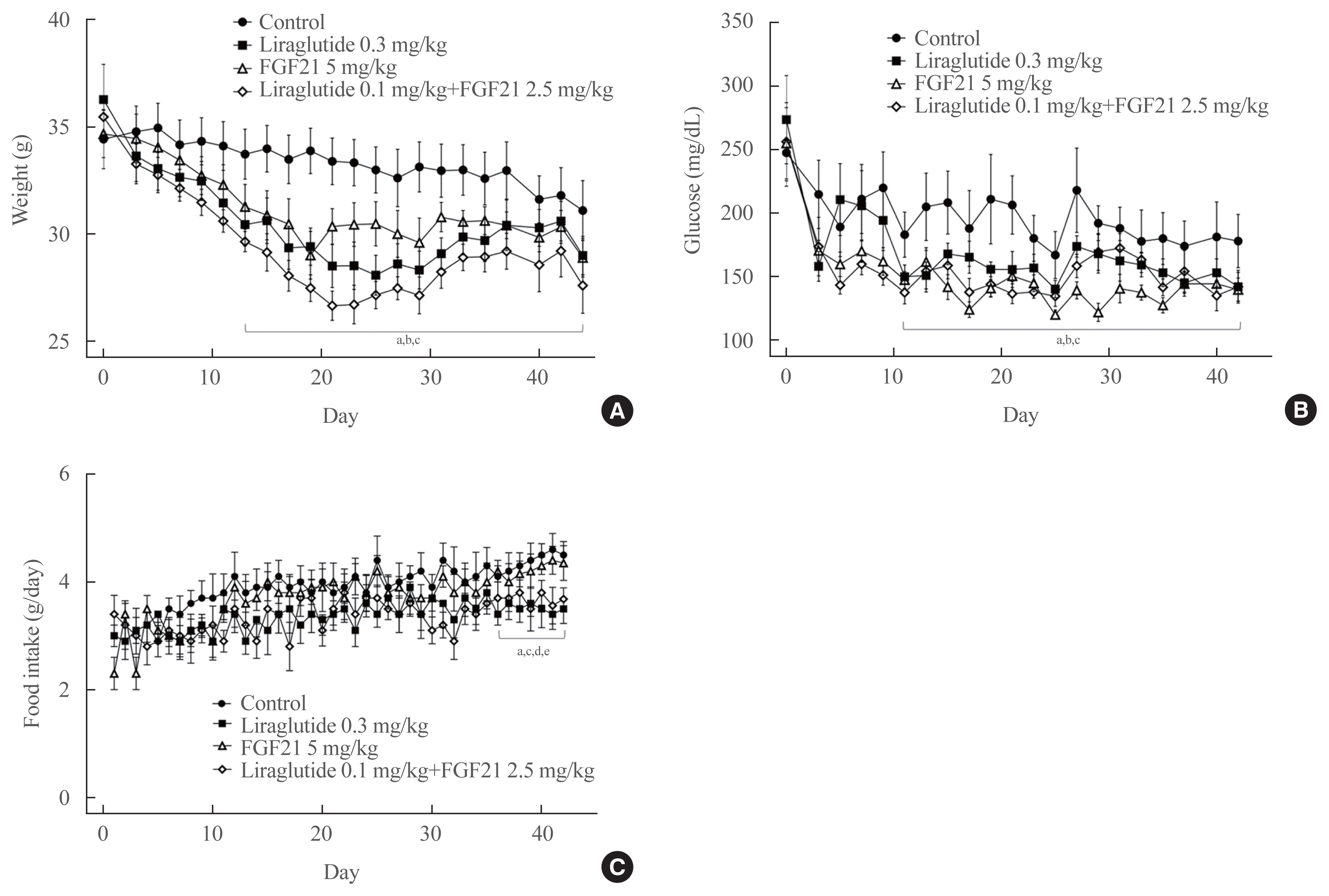
Effects of a 6-week treatment with liraglutide, FGF21, or their combination on body weight, food intake, and blood glucose concentrations
Among the animals that were fed a high-fat and atherogenic diet, weight was lower in the liraglutide, FGF21, and low-dose combination of liraglutide and FGF21 groups compared with the control group, starting at 13 days after treatment and with the exception of days 40 and 42 (Fig. 2A). Random glucose concentrations were lower in the liraglutide, FGF21, and low-dose combination of liraglutide and FGF21 groups compared with the control group, starting at 11 days after treatment and with the exception of days 23, 31, and 35 (Fig. 2B). Similarly, food intake was lower in the liraglutide alone and low-dose combination of liraglutide and FGF21 groups than in the FGF21 or control groups starting at 36 days after treatment (Fig. 2C).
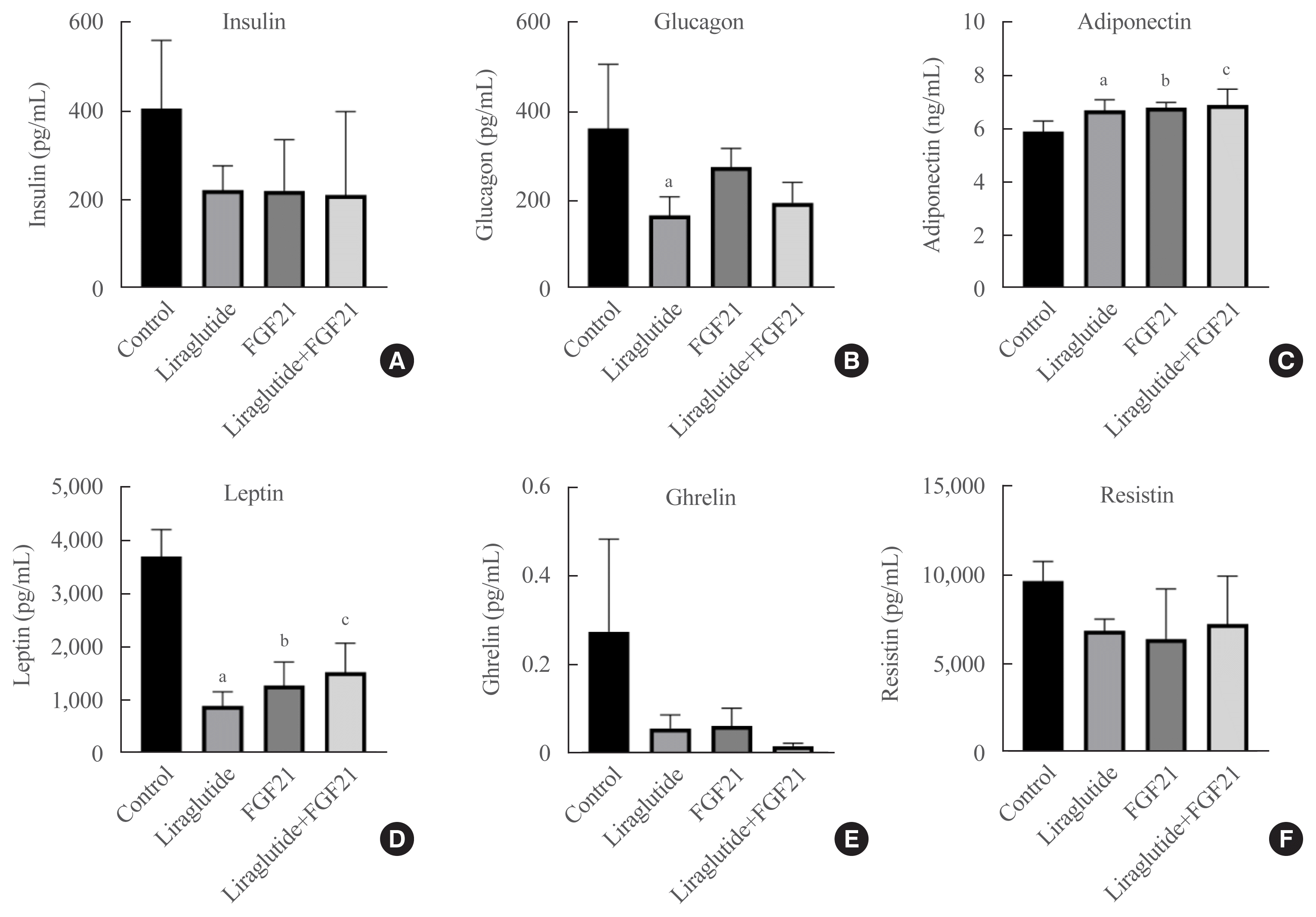
Effect of the liraglutide and FGF21 combination treatment on biochemical parameters
Liraglutide, FGF21, and their low-dose combination regimen decreased insulin levels, albeit without statistical significance. Glucagon levels were significantly suppressed only by the liraglutide treatment (Fig. 3). All three treatments increased the levels of circulating adiponectin in these high-fat diet-fed mice. Furthermore, leptin levels were significantly lower in all treatment groups. Finally, the levels of both ghrelin and resistin exhibited decreasing patterns, albeit without statistical significance (Fig. 3).
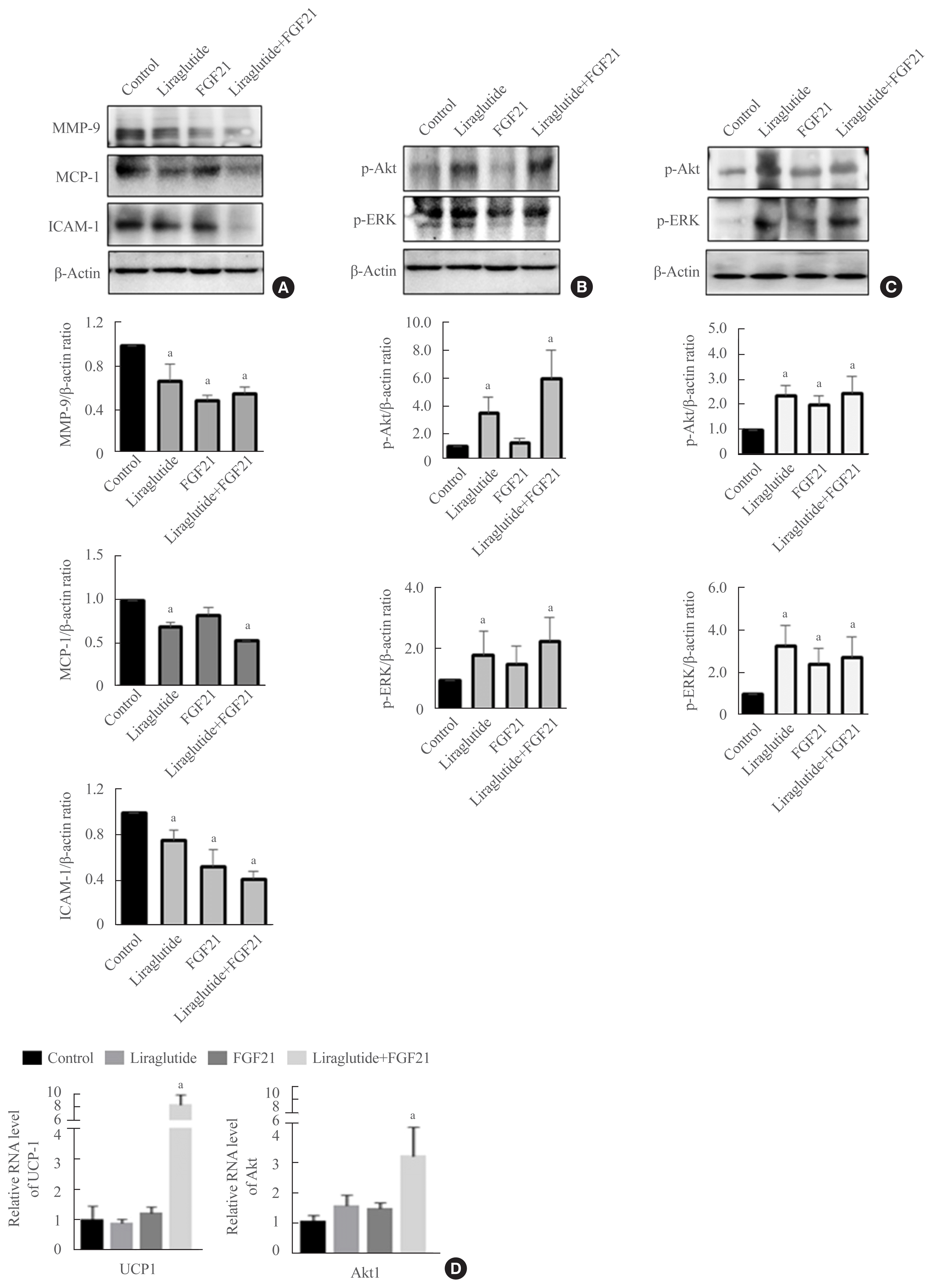
Hepatic expression of MMP-9, MCP-1, ICAM-1, UCP1, and Akt1
Both liraglutide alone and its combination with FGF21 downregulated MMP-9, MCP-1, and ICAM-1 in the mouse liver compared with the control group (Fig. 4A). To investigate whether UCP1 and Akt1 were involved in the regulation of glucose metabolism in the combination treatment, the hepatic expression levels of UCP1 and Akt1 were tested. As shown in Fig. 4D, the mRNA expression of UCP1 and Akt1 was significantly increased in animals treated with a low-dose combination of liraglutide and FGF21 compared with the controls or either liraglutide-alone- or FGF21-alone-treated mice. Therefore, the liraglutide and FGF21 combination treatment exerted a synergistic effect on the expression of these molecules.
Effect of liraglutide and FGF21 combination on the p-Akt and ERK pathways in the liver and visceral fat
To identify the signaling pathways involved in the effects of the liraglutide and FGF21 combination treatment, we examined the phosphorylation of Akt and ERK by Western blotting. In the liver and visceral fat of obese mice, p-Akt and p-ERK were upregulated in the liraglutide-treated and combination groups compared with the control group (Fig. 4B, C). Moreover, in the liver, the combination treatment exerted a partial additive effect.
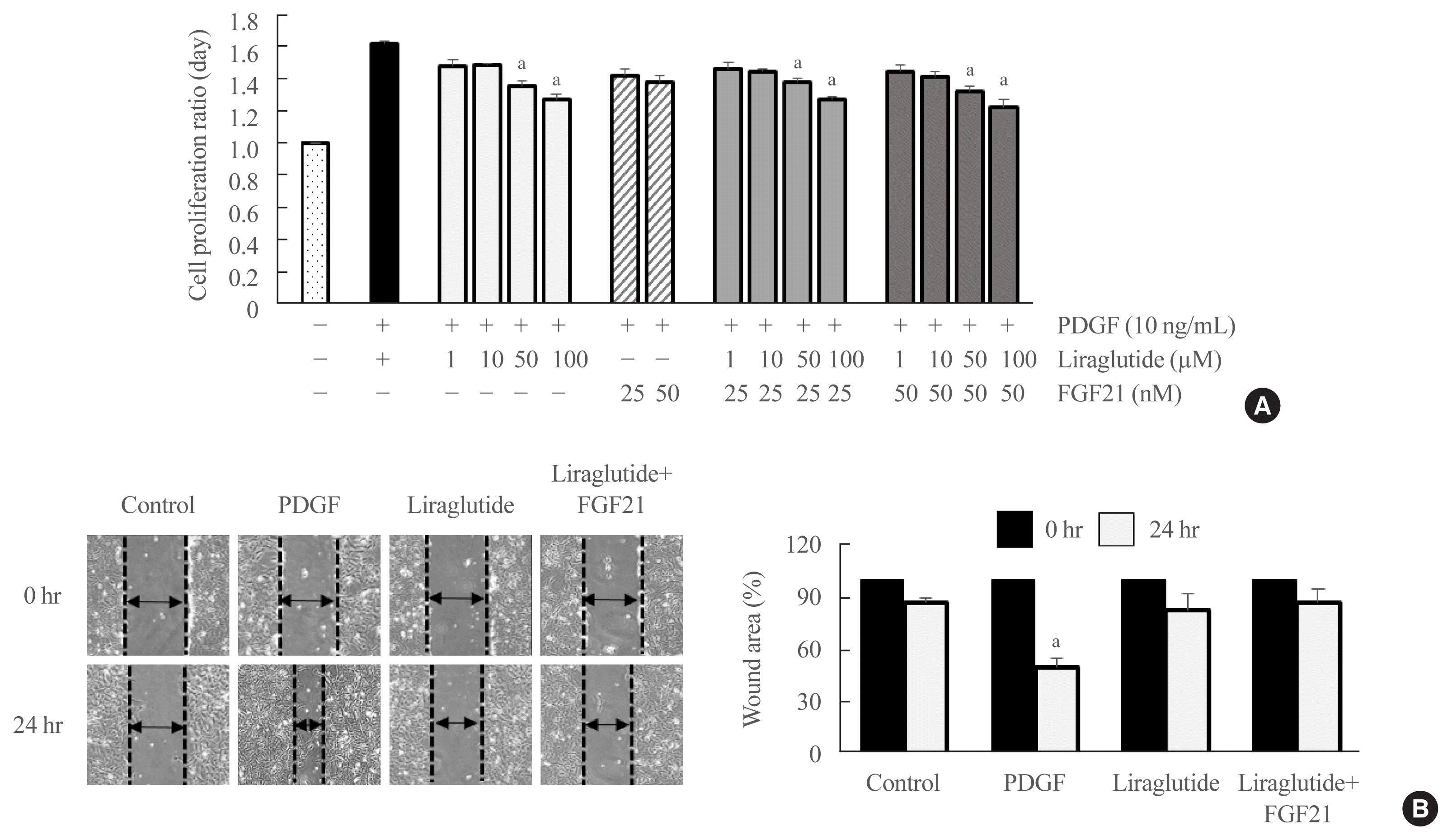
Effect of the liraglutide and FGF21 combination treatment on the proliferation of VSMCs
The PDGF-induced proliferative effects were significantly reversed by liraglutide concentrations above 50 μM and by the combined treatment with concentrations above 50 μM liraglutide and FGF21 (Fig. 5A). Subsequent experiments were performed using toxic cellular concentrations (50 μM liraglutide alone or 10 μM liraglutide combined with 50 nM FGF21). We focused on further defining the therapeutic combinational potential of liraglutide and FGF21.
Effect of the liraglutide and FGF21 combination treatment on the PDGF-induced migration of VSMCs
A wound-healing migration assay was performed to determine the effects of liraglutide alone or the liraglutide and FGF21 combination treatment on PDGF-induced VSMC migration. VSMC motility was significantly increased after a 24-hour treatment with PDGF compared with both the liraglutide-alone and the combination groups (Fig. 5B).
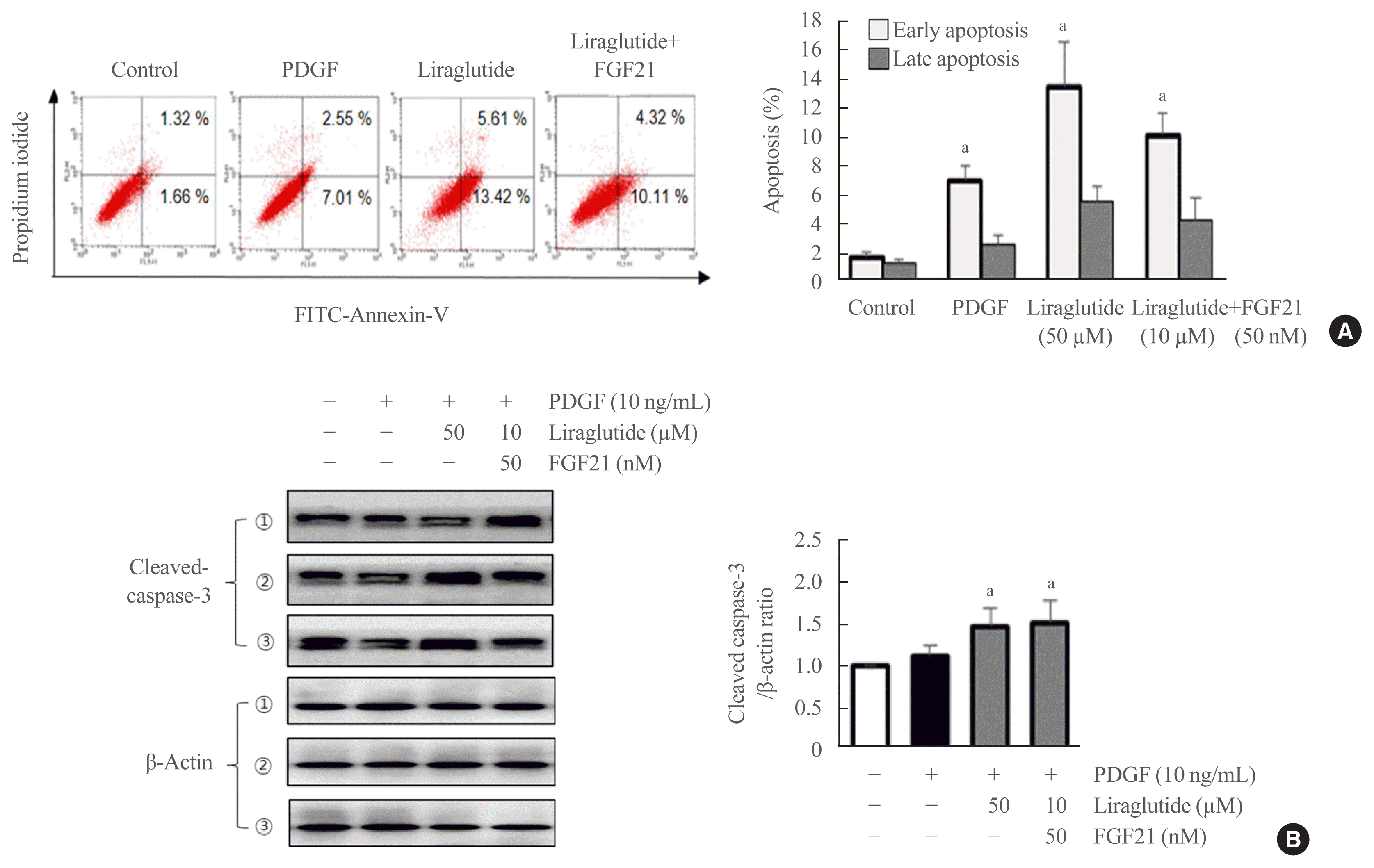
Effect of the liraglutide and FGF21 combination therapy on VSMC apoptosis
To investigate the possible anti-apoptotic effects of the liraglutide and FGF21 combination therapy, a flow cytometry assay was performed using VSMCs. After 24 hours of treatment with the drugs, approximately 19.03% and 14.43% of apoptotic cells were detected in the liraglutide-alone and the combination groups, respectively. In addition, early apoptosis was dramatically increased in cells treated with liraglutide alone or the liraglutide and FGF21 combination compared with PDGF-stimulated cells. Therefore, the proportion of apoptotic cells was higher upon treatment with liraglutide alone and the liraglutide and FGF21 combination compared with PDGF treatment (Fig. 6A). The activation of caspase-3 is marked by the cleavage of procaspase-3 and is the executioner of apoptosis. We examined the levels of cleaved caspase-3 by Western blotting. Apoptosis was effectively induced in VSMCs treated with liraglutide and the liraglutide and FGF21 combination compared with PDGF-stimulated cells (Fig. 6B).
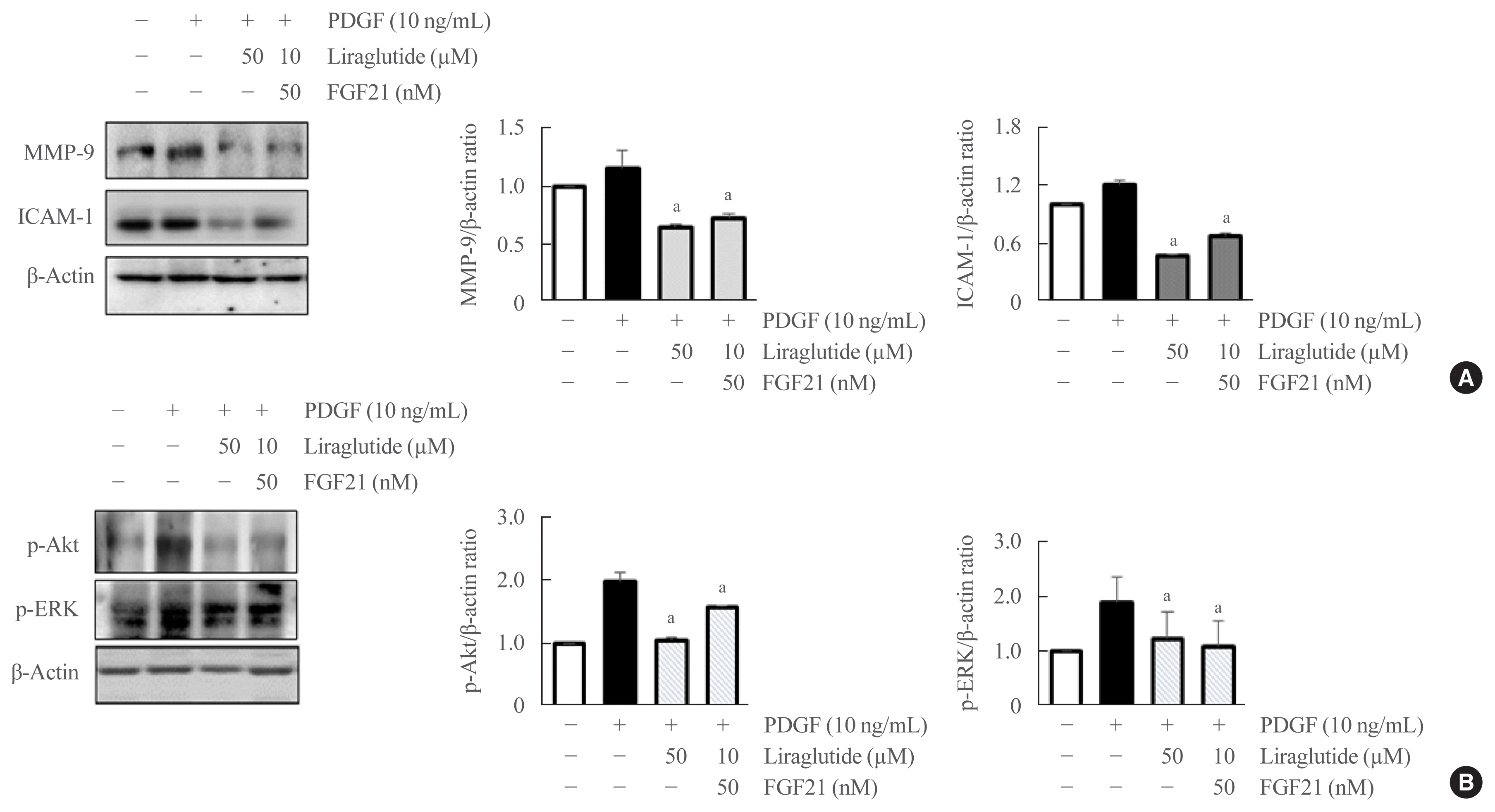
Effect of liraglutide and FGF21 combination on the expression of MMP-9, ICAM-1, p-Akt, and ERK1/2 in VSMCs
The expression of MMP-9 and ICAM-1 was significantly suppressed in VSMCs after treatment with both liraglutide alone and the liraglutide and FGF21 combination compared with PDGF treatment (Fig. 7A). As shown in Fig. 7B, the treatment with liraglutide, either alone or in combination with FGF21, significantly suppressed the PDGF-induced phosphorylation of Akt and ERK in VSMCs.
Effect of combined liraglutide and FGF21 treatment on the expression of phosphorylated p-p38 and p-JNK in the liver, pancreas, and white adipose tissue
In the Western blot analysis, there was significantly increased expression of p-p38 in the liraglutide alone treatment in the pancreas and in the liraglutide alone and liraglutide and FGF21 combination treatment in the pancreas and white adipose tissue (WAT) (Supplemental Fig. S1). There was a significant decrease in the expression of p-JNK by FGF21 alone treatment in the liver. Otherwise, there were no changes in the expression levels of these two proteins in the liver, pancreas, and WAT by treatments with liraglutide alone, FGF21 alone, or a low-dose combination of both.
DISCUSSION
Here, a low-dose combination of liraglutide and FGF21 reduced atheromatous plaque area in aorta and decreased body weight and blood glucose concentrations in high-fat diet-induced obese mice compared with controls and caused a concomitant upregulation of the UCP1 and Akt1 mRNA. The low-dose combination of liraglutide and FGF21 suppressed the proliferation of VSMCs, accompanied by suppression of MMP-9 and ICAM-1, thereby reducing the development of atherosclerosis. Furthermore, treatment with this combination therapy downregulated the inflammatory markers MMP-9 and MCP-1, as well as the adhesion molecule ICAM-1. Our in vitro and in vivo experiments showed that the anti-inflammatory and anti-atherosclerotic effects of the GLP-1 analogue and FGF21 combination therapy occurred via the Akt and ERK pathways. To the best of our knowledge, this is the first study that has demonstrated that the combination of a GLP-1 analogue and FGF21 is effective against the vascular complications of diabetes.
A recent study using male C57BL/6 mice reported that the UCP1 protein is expressed in the liver as well as in WAT [26]. In that study, a ketogenic diet upregulated UCP1 in the liver. Another study using Sprague Dawley rats also reported that liraglutide injection increased the expression of the UCP1 gene and UCP1 protein in the liver [27]. UCP1 is a mitochondrial protein that is involved in inflammatory cytokine signaling pathways and is implicated in liver function. Therefore, UCP1 is of great significance for application as a reasonable indicator of metabolic health. Moreover, UCP1 activation in brown adipose tissue is essential for the maintenance of the core body temperature, and is associated with liver steatosis and oxidative stress [28,29]. Moreover, Akt activation results in marked alterations in glucose and lipid metabolism [30]. Here, we showed that the low-dose combination of a GLP-1 analogue and FGF21 exerted body-weight-reducing effects, decreased fasting glucose concentration, and upregulated the UCP1 and Akt1 mRNAs, suggesting that this treatment reduces obesity and improves insulin sensitivity by inducing UCP1 and Akt gene expression. The elevated adiponectin levels and the enhancement in mitochondrial function induced by the combination therapy may play an intrinsic role in the upregulation of UCP1 and Akt1 in the liver [27,31]. There is no clear answer to why treatments with liraglutide alone or FGF21 alone treatment did not increase UCP1 expression. We speculate that these two molecules might act synergistically. A previous study showed that UCP1-dependent thermogenesis was not required for FGF21 to improve glycemic control or to reduce the levels of circulating cholesterol or free fatty acids [32]. Thus, some metabolic actions of FGF21 are not associated with UCP1.
In the current study, the food intake was lower in the liraglutide-alone or liraglutide and FGF21 combination groups than in the control group. The body-weight-reducing effects of GLP-1/FGF21 treatment could be also due to increased energy expenditure, leading to a negative energy balance. Energy expenditure has different elements, such as resting metabolic rate, physical activity, and thermogenesis. It also depends on the oxidation of substrates. It was reported that GLP-1 administration increased whole-body oxygen consumption in rats, an index of energy expenditure and body temperature by 0.3°C [33]. In another animal study, GLP-1 administration increased thermogenesis in brown adipose tissue [34]. Unfortunately, we did not measure body temperature in the present study. Taken together, these preclinical findings indicate that GLP-1 analogues might influence the central regulation of energy balance by affecting dietary intake, energy expenditure, and thermogenesis in particular.
Here, liraglutide, FGF21, and their combination increased adiponectin levels and decreased leptin levels. Decreased serum levels of adiponectin have been suggested as a mechanism underlying obesity-induced insulin resistance [35]. Adiponectin possesses potent anti-inflammatory and anti-atherosclerotic activities via its multiple actions in the vascular system [36]. Leptin is a crucial hormone that is responsible for the endocrine control of energy homeostasis [37]. The development of resistance to leptin is a hallmark of obesity [38]. The favorable changes in adiponectin and leptin afforded by liraglutide alone or the combination treatment with liraglutide and FGF21 suggest their systemic effects on the cardiometabolic system and energy metabolism.
Our group showed that a synthetic GLP-1 molecule together with exenatide, another GLP-1 analogue, attenuated the PDGF-induced proliferation of VSMCs [39]. Another study showed that GLP-1 treatment effectively attenuated the proliferation of VSMCs induced by angiotensin II [40]. Proliferation and migration of PDGF-induced VSMCs accompany the formation of vascular injury, which leads to the development of atherosclerosis [41]. Apoptosis is also important for the control of excessive cell proliferation and elimination of harmful cells, being regulated both positively and negatively by growth factors, including IGF-1 and PDGF [42,43]. A study reported that liraglutide inhibits VSMC proliferation by promoting cell-cycle arrest via the activation of AMPK [40]. The imbalance between VSMC proliferation and apoptosis is related to neointimal formation, indicating high proliferation/apoptosis ratio in VSMCs in the arterial wall after injury or inflammatory stimuli [44]. In the present study, we found that liraglutide alone (50 and 100 μM) or in combination with FGF21 inhibited the proliferation of VSMCs in response to PDGF. The liraglutide-alone and liraglutide and FGF21 combination treatments increased the proportion of apoptotic cells and the levels of caspase-3. However, the FGF21 alone treatment (25 and 50 nM) showed a slight decrease in the proliferation of VSMCs but this lacked statistical significance. This might have been because of the use of low-dose FGF21. From a different context, the vascular endothelial cells lining the atheromatous region and apoptosis of vascular endothelial cells are critical in maintaining the stability of atheromatous plaques [45]. Apoptosis of VSMCs directly comprising atheromatous plaques might confer instability, but in contrast, other reports suggest that apoptosis of VSMCs contributes to the reduced development of atheromatous plaques [39,46]. Taken together, our findings suggest that the combination of liraglutide and FGF21 suppresses the proliferation of VSMCs more efficiently by inducing apoptosis.
Atherosclerosis is an inflammatory disease and a major cause of the upregulation of adhesion molecules for leukocytes in VSMCs via altered phenotypic expression at the injured sites [47]. MMPs are proteolytic enzymes that act on a variety of connective tissue proteins, and increasing evidence supports their involvement in the development of atherosclerotic plaques [48]. MCP-1 activates monocytes/macrophages at the site of tissue injury and regulates adhesion molecules [49]. The expression of MMP-9 and ICAM-1 is increased in VSMCs during the development of atherosclerosis [50,51]. In addition, monocytes expressing MCPs affect the growth of cells within atherosclerotic lesions [52]. Importantly, GLP-1-based treatment yielded beneficial anti-inflammatory effects in the liver and vein endothelial cells, and the use of FGF21 prevented the inflammation and oxidative stress associated with atherosclerosis in vivo [53,54]. Here, in vivo experiments showed that the combination of liraglutide and FGF21 downregulated MMP-9, MCP-1, and ICAM-1 and increased the plasma levels of adiponectin. These data support the notion that decreased inflammation is closely related to the inhibition of atherosclerosis in diabetes. Thus, liraglutide and FGF21 may exert a synergistic effect in the prevention of diabetes-associated atherosclerosis, with a possibly higher anti-inflammatory potential than that observed for GLP-1, FGF21, and other monotherapies.
In our experiment, the most noteworthy effect of the GLP-1 analogue was its anti-inflammatory ability. FGF21 induced sizable improvements in hepatic steatosis [55]. Consequently, the GLP-1 analogue and FGF21 combination therapy may be more effective at promoting insulin sensitivity in the liver, compared with the GLP-1 analogue alone. In addition, the combination treatment significantly reduced PDGF-stimulated Akt and ERK1/2 phosphorylation in VSMCs, suggesting its therapeutic applicability in the prevention of atherosclerosis. Our in vivo experiments explored the molecular events downstream of insulin signaling and showed that the liraglutide and FGF21 combination significantly increased Akt and ERK1/2 phosphorylation in the liver compared with high-fat-fed control mice. Because Akt and ERK1/2 are signaling molecules that are involved in the pathophysiology of vascular complications, these results support the beneficial effect of the two agents on vasculature in a synergistic fashion.
Gastrointestinal side effects are common in high-dose GLP-1 analogue therapy [14]. In addition, the FGF21 protein has several limitations as a therapeutic agent, including a short half-life [22]. To reduce the side effects induced by liraglutide treatment and overcome the shortcomings of FGF21, a low-dose combination of the two agents might be an ideal strategy from a safety perspective.
In conclusion, the present study expands our understanding of the effects elicited by the combination of a GLP-1 analogue with FGF21 and provides new insights regarding an ideal therapeutic option for preventing the development of atherosclerosis in patients with diabetes. We showed an additive or partial synergistic effect of the liraglutide and FGF21 combination compared with the control or the single drugs in several in vivo experiments. Thus, FGF21 might be a suitable agent to enhance the effect of GLP-1 analogues. Taken together, our results indicate that, by acting on the Akt and ERK1/2 pathways, the combined treatment with a GLP-1 analogue and FGF21 protects against inflammation and atherosclerosis. These results suggest that the GLP-1 analogue and FGF21 combination is of potential clinical relevance in the treatment of vascular complications related to diabetes.
 XML Download
XML Download