

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Cardiovascular disease (CVD) is the leading cause of death worldwide; it includes various disorders such as stroke, cardiac muscle diseases, and vascular system diseases and affects blood supply to pivotal organs, including the heart and brain.1) Currently, the most effective treatment methods for patients utilize various technologies and devices to assist the ventricular system; these include catheter interventions and implantable cardio-defibrillators with resynchronization cardiac systems.2)3) Although these technologies were developed to treat CVD particularly, they have shown only moderate efficacy in reducing the rate of mortality from CVD events. Since the 1990s, many clinicians and researchers have worked to develop the concept of “cell therapy,” the main objective of which is to restore the functions of dying organs through a mechanism known as cardiovascular regeneration. This new and innovative therapy was first reported using patient-derived autologous bone marrow cells. There have been many reports on the improvement of therapeutic efficacy using cell therapy, including different cell sources, genes, materials, reagents, chemicals, and transplantation methods.4)
Although many studies have used bone marrow-derived cells,5)6)7)8)9)10)11)12)13)14) cardiac progenitor cells (CPCs),15) and mesenchymal stem cells,16)17) these candidates have shown disappointing clinical results in terms of regenerative ability, feasibility, and patient safety.18) These cell types are difficult to use in cell therapy owing to the impairment of self-revascularization potential by the surrounding ischemic environment, and due to the loss of transplanted cells.19) Therefore, it is necessary to enhance the regeneration process via direct cell transplantation or through the administration of growth factors and cytokines. Recently, to overcome this drawback, pluripotent stem cells (PSCs) such as embryonic stem cells (ESCs) and induced pluripotent stem cells (iPSCs) are emerging as beneficial cell sources for cardiac regeneration. These cells have unique and genuine characteristics; they can differentiate into several cardiac lineage cells, including cardiomyocytes (CMCs).20) Based on the unprecedented notion that somatic cells can be reprogrammed using defined factors, numerous studies have reported the ability of these iPSCs to differentiate into functional and mature cardiac lineage cells. This review describes the different types of PSC-derived CMC and outlines key markers that can be used to identify PSC-derived cardiac lineage cells. Furthermore, this paper will review recent findings on the direct conversion of adult somatic cells into cardiac lineage cells while highlighting their therapeutic potential in regenerative medicine and future treatment of CVDs.
PLURIPOTENT STEM CELLS-DERIVED CARDIOMYOCYTES
Stem cells are classified based on their ability to differentiate into mature cells. Notably, cardiac lineage cells, such as CMCs derived from iPSCs, can be obtained easily from individual patient and can be used to generate disease-specific models. These patient-derived disease models can be used to understand the genetic and molecular mechanisms underlying hereditary CVD phenotypes; they can also reveal novel therapeutic targets for personalized medicine.21)22)23)24)25)
Somatic stem cells (SSCs) are non-ESCs that are found in postnatal development; they reside in a location known as the “stem cell niche”.26)27) SSCs are present in various tissues, such as the bone marrow, skeletal muscles, cord blood, peripheral blood, adipose tissue, lungs, and heart.28)29)30)31)32)33)34)35)36)37)38)39) However, they have limited multipotent capacity, and their main roles are to maintain the homeostasis of fully differentiated cells and regenerate damaged tissues. The limited ability to differentiate into desired target lineages and the difficulty of obtaining pure stem cells showed that new methods need to be developed to facilitate therapeutic application.
ESCs are classified as PSCs derived from the inner cell mass of blastocysts (Table 1). ESCs can be cultured in a stringently controlled environment, which means that they can divide and proliferate without undergoing differentiation. Therefore, these ex vivo cultured cells serve as an excellent candidate for cell therapies for various chronic diseases, including metabolic syndrome, brain disorders, and cardiomyopathies. However, due to serious ethical issues and technical problems, the use of ESCs in clinical settings is highly restricted.40) ESC-derived differentiated target cells are used for cell transplantation. However, this can induce teratoma formation in target tissues due to presence of undifferentiated cells among transplanted cell populations. This has raised safety concerns regarding the use of these cells for transplantation.41)42)43) As for ethical concerns, the use of human ESCs is debatable with many advocates anxious of using a live embryo samples for isolation of human ESCs. Taken together, these challenges have hindered the use of human ESCs and live human embryos in laboratories.
Table 1
Overview of PSCs
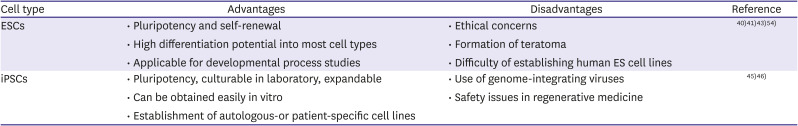
| Cell type | Advantages | Disadvantages | Reference |
|---|---|---|---|
| ESCs |
• Pluripotency and self-renewal • High differentiation potential into most cell types • Applicable for developmental process studies |
• Ethical concerns • Formation of teratoma • Difficulty of establishing human ES cell lines |
40) 41) 43) 54) |
| iPSCs |
• Pluripotency, culturable in laboratory, expandable • Can be obtained easily in vitro • Establishment of autologous-or patient-specific cell lines |
• Use of genome-integrating viruses • Safety issues in regenerative medicine |
45) 46) |
For their practical applicability in a clinical environment, large-scale production of cells, with tightly regulated protocols and culture environments, is required. Therefore, researchers in stem cell field have focused on exploring other options for industrial and clinical applications, such as converting adult somatic cells into PSCs. This process of requisite pluripotency, which is called “cell reprogramming,” represents a paradigm transition from a traditional method of cell therapy and in the current understanding of cellular biology and the plasticity of the mature and differentiated states of somatic cells.
Sir John Gurdon first successfully reprogrammed cells by cloning a frog using somatic cell nuclear transfer; he was subsequently awarded the Nobel Prize in Medicine in 2012.44) Since then, this discovery has led stem cell researchers to develop the concept of reprogramming by inducing pluripotency in somatic cells (Table 1). In 2006, Dr. Shinya Yamanaka (Nobel Prize in Medicine, 2012) and his colleagues generated the first iPSCs by inserting the defined transcription factors (OCT4, SOX2, KLF4, and c-MYC [OSKM]), which are known pluripotency-maintenance genes, into the nuclei of somatic cells using retrovirus.45)46) These human iPSCs (hiPSCs) are almost equivalent to human ESCs in terms of their self-renewal capacity and potential to differentiate into multiple lineages. This discovery, combined with the development of cellular reprogramming strategies, has made it possible to use patient-specific iPSCs for cell therapy while overcoming ethical issues and concerns regarding immune rejection. In the early studies on reprogramming of somatic cells, genome-integrating viruses were used, which limited their clinical applicability due to random insert mutations and tumor formation. However, this technology has now advanced to the stage where iPSCs can be generated using non-genetic methods, including plasmid vectors, adenoviruses, Sendai viruses, proteins, and modified ribonucleic acids (RNAs).47)48)49)50)51)52)53)54) It is thus necessary to establish a well-settled differentiation protocol to handle iPSCs with favorable properties.
Despite the development of promising surgical interventions and pharmacological treatments for CVDs, heart transplantation remains the last therapeutic option for patients with end-stage heart failure. Therefore, approaches that could refurbish the failing population of CMCs to rescue damaged myocardium could be used to recover heart function. The CMCs derived from hiPSCs represent promising therapeutic sources for CVDs. Since ESCs and iPSCs are similar in characteristics and differentiation potential, the current study to differentiate hiPSCs into CMCs are in parallel with previously established protocols for deriving CMCs from human ESCs.
IN VITRO DIFFERENTIATION OF PLURIPOTENT STEM CELLS INTO CARDIOMYOCYTES
For their clinical application, the PSC-derived CMCs need to be highly pure and enriched, largely scalable for mass production, and cultured in a xenogeneic-free environment. There are traditional small-scale protocols for CMC differentiation, including formation of embryoid body (EB); the co-culture of endodermal cell lines (END-2) with undifferentiated PSCs; and the treatment of defined cardiogenic growth factors to confluent PSC monolayer.55) In general, EB-based formation assays are the most used methods to generate CMCs from iPSCs in laboratories. This type of assay involves undifferentiated iPSCs as aggregates in a suspended culture, which causes them to form EB structures. These EBs are cultured in condition which serum is absent to provide pluripotency. In the presence of several cytokines including activin A and bone morphogenetic protein 4 (BMP4), EBs differentiate into mature CMCs.55)56)57) The co-culture of visceral endoderm-like cells with PSCs induces CMC differentiation; the visceral endoderm is an extraembryonic tissue that is formed prior to gastrulation during embryogenesis, which pivotal factors that are involved in development are secreted. Mummery et al. reported that CMC differentiation can be induced by co-culturing visceral endoderm-like cells with human and mouse PSCs.58)59)60) Lastly, the subsequent addition of growth factors to PSCs has been shown to induce direct CMC differentiation by recapitulating the cardiac development during embryogenesis in vitro. Stimulating human PSCs (hPSCs) with supplementing growth factors like basic fibroblast growth factor, Wnt3, BMP4 and activin A, followed by DKK1 or other Wnt inhibitors, has been shown to induce cardiac differentiation.56)57)61) Subsequently, the addition of other regulators, such as vascular endothelial growth factor,56) CHIR and IWR-1,62)63) Noggin,64) transforming growth factor-β signaling inhibitor, or sonic hedgehog signaling activation,65) has also been found to increase the differentiation efficiency of CMCs.
These traditional protocols are used for small-scale production and are mainly applicable for research. Although these protocols can achieve high percentage yields of purified CMCs production, they have limited reproducibility and scalability, and the resulting CMCs are mostly heterogeneous. In addition, billions of CMCs are required for high-throughput assays, tissue engineering, and animal models. Although large-scale production can be achieved by scaling up culture conditions, this process is space- and/or cost-inefficient.
The large-scale production of PSC-derived CMCs has been achieved using three-dimensional (3D) platforms that are compatible with industry. These include cultures that rely on matrix, a polymer with sphere cultures and cultures do not depend on matrix platforms, like bioreactors and spinning flasks. Several studies demonstrated successfully generated high yields of functional CMCs from hPSCs on a large scale, using bioreactors and chemical modulators.66)
Taken together, many modulators and factors, including cell populations, mediators that induce cardiac lineage differentiation, and culturing conditions, are pivotal in mediating the differentiation of iPSCs into CMCs. Optimizing these factors is important for achieving highly efficient and successful differentiation of PSCs into CMCs.
MARKERS FOR CARDIAC LINEAGE CELLS
The CMCs that are differentiated from PSCs comprise heterogeneous CPC populations and partially differentiated CMCs. Hence, differentiated CMCs must be purified and enriched. In order to achieve this, it is necessary to determine the origins of cardiac cell types and identify the markers of cardiac lineage cells to understand the origin of heart malformations during development. Furthermore, it is necessary to produce a cell-therapy-applicable cardiac tissue for use in regenerative medicine. In this section, several key markers for cardiac lineage cells during the differentiation of PSCs into CMCs are discussed (Table 2).
Table 2
Markers for generation of PSC-derived cardiac lineage cells
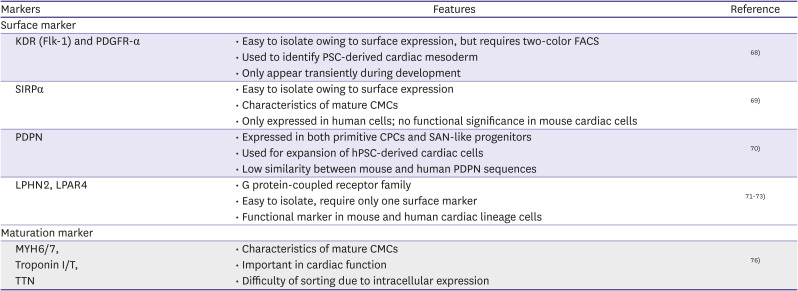
| Markers | Features | Reference | |
|---|---|---|---|
| Surface marker | |||
| KDR (Flk-1) and PDGFR-α |
• Easy to isolate owing to surface expression, but requires two-color FACS • Used to identify PSC-derived cardiac mesoderm • Only appear transiently during development |
68) | |
| SIRPα |
• Easy to isolate owing to surface expression • Characteristics of mature CMCs • Only expressed in human cells; no functional significance in mouse cardiac cells |
69) | |
| PDPN |
• Expressed in both primitive CPCs and SAN-like progenitors • Used for expansion of hPSC-derived cardiac cells • Low similarity between mouse and human PDPN sequences |
70) | |
| LPHN2, LPAR4 |
• G protein-coupled receptor family • Easy to isolate, require only one surface marker • Functional marker in mouse and human cardiac lineage cells |
71) 72) 73) | |
| Maturation marker | |||
|
MYH6/7, Troponin I/T, TTN |
• Characteristics of mature CMCs • Important in cardiac function • Difficulty of sorting due to intracellular expression |
76) | |
CMCs = cardiomyocytes; CPC = cardiac progenitor cell; FACS = fluorescence activated cell sorting; Flk-1 = fetal liver kinase 1; KDR = kinase insert domain receptor; LPAR4 = lysophosphatidic acid receptor 4; LPHN2 = latrophilin-2; MYH = myosin heavy chain; PDGFR-α = platelet-derived growth factor receptor alpha; PDPN = podoplanin; PSC = pluripotent stem cell; SAN = sinoatrial node; SIRPα = signal regulatory protein alpha; TTN = titin.
The functions of the heart depend on interactions between different cardiac cell types. These cells are stringently controlled by precise spatiotemporal regulation during the development of the heart structure. Recent technologies, such as single-cell transcriptomics, have begun to reveal the identity of cell heterogeneity through early developmental stages. Furthermore, the same strategy can be applied to trace lineages during undifferentiated and fully differentiated stages, using hPSCs. Hence, identifying the markers involved in human CMCs is a popular research topic, on which studies are ongoing.
Although CPCs have been identified using multiple markers, the development of efficient methods to obtain and expand iPSC-derived CPCs in vitro remains challenging.67) As most of these markers are intracellular molecules or transcription factors, it is necessary to find a different marker that can be used to purify CMCs. Kattman et al.68) revealed that kinase insert domain receptor (KDR; Flk-1) and platelet-derived growth factor (PDGFR)-α are co-expressed in emerging cardiac mesoderm. However, these markers are only expressed for a short period during development, and practical two-color flow cytometric isolation is needed for enrichment. In addition, many studies have utilized signal-regulatory protein alpha (SIRPα),69) and podoplanin (PDPN)70) as cell surface markers to purify and expand CPC populations derived from hPSCs. However, murine cardiac cells do not express SIRPα; hence, it is difficult to perform studies of functional significance, such as studies using a knockout or overexpression model, via this approach.
Recently, the surface protein G protein-coupled receptor, latrophilin-2 (LPHN2),71)72) and lysophosphatidic acid receptor 4 (LPAR4)73) were reported as novel CPC markers. LPHN2 is a functional marker for isolating CPCs and CMCs derived from PSC. In addition, it has been demonstrated that Lphn2-deficient ESCs and iPSCs do not achieve complete cardiac differentiation. In contrast, LPAR4 expression was observed to be transient during in vitro cardiac differentiation from the undifferentiated state of ES/iPS cells. Based on the transient expression of LPAR4 during differentiation, an optimal protocol has been established for the sequential stimulation and inhibition of LPAR4 signaling.
The expression pattern of maturation-marker genes identified in mice is similar to that in humans.74)75) The transitions of sarcomeric gene isoform occurs from the fetal to adult stages during development. The most well-known marker is the cardiac myosin heavy chain (MHC - also known as MYH). There are two isoforms: α-MHC (also known as MYH6) and β-isoform (β-MHC - also known as MYH7). The β-isoform is the most predominant one in adult CMCs, but this switches in mouse hearts. Troponins consist of fast- (the TNNC2, TNNI2, and TNNT3 genes) and slow-twitch skeletal (the TNNC1, TNNI1, and TNNT1 genes) and cardiac myocytes (the TNNI3 and TNNT2 genes). The heart selectively produces Troponins, TnnI, and TnnT, but TnnC is also produced in the slow-twitch skeletal muscle. Regions of different amino acid sequences (cTnnI and cTnnT) can be used to distinguish these markers from each other. TNNT2 messenger RNA transcribes multiple alternatively spliced transcripts, thereby synthesizing different proteins (cTnnT1-cTnnT4). Among them, cTnnT3 is the dominant isoform in the normal adult heart.76) TroponinI has evolved into three isoforms that are encoded by TNNI1 (slow skeletal, ssTnI), TNNI2 (fast skeletal, fsTnI), and TNNI3 (cardiac, cTnI). In adult CMCs, cTnI is the major isoform that is expressed, although in hiPSC-derived CMCs, ssTnI is the primary isoform. Titin (TTN) also has three isoforms: N2B, N2BA, and fetal cardiac titin (FCT). In hiPSC-derived CMCs, N2BA is predominant, whereas N2B is mainly expressed in adult CMCs.77)78) Interestingly, hiPSC-derived CMCs show lower expression levels of several cardiac-related genes, including sarcoplasmic reticulum ATPase (SERCA2), caveolin 3 (CAV3), and potassium voltage-gated channel (KCNH2), among others.78)79)80)
Measuring the ratio of TNNI3 to TNN1 is one option for verifying the maturation status of hiPSC-derived CMCs.81) A transcriptome-based approach could represent another promising option for more precisely evaluating their mature state. Single-cell transcriptomic analysis can also verify the developmental stages of CMCs during hPSC differentiation. However, these approaches are mostly restricted to mouse PSC-derived CMCs, as limited transcriptomic data are available for fully developed human hearts.80)82)83)84)
DIRECT CARDIAC REPROGRAMMING
The direct reprogramming of fully mature and terminally differentiated somatic cells into another lineage, using a combination of defined factors, has changed the traditional and fundamental concepts of previously established basic knowledge regarding differentiated cells. In recent years, many studies have reported the direct conversion of somatic cells into various cell types. The generation of desired functional cell types that can be transplanted into patients is a promising approach for biomedical applications and clinical fields.
Ieda and colleagues85) used mouse fibroblasts that were engineered to express green fluorescent protein during cardiac reprogramming (Table 3). After screening process, previously reported 14 candidate factors were carefully narrowed down to 3 transcription factors, Gata4, Mef2c, and Tbx5. Subsequently, two study groups have reported that the delivery of a combination of Gata4, Mef2c, and Tbx5 using retrovirus can convert fibroblasts in vivo, residing at sites of infarction, into CMCs. Furthermore, this approach reduces the extent of injury.86)87) Another study reported that adding the transcription factor Hand2 improves the efficiency of the direct conversion of fibroblasts into CMCs, both in vitro and in vivo.88) Subsequently, Jayawardena et al. demonstrated that microRNAs (miRNAs), in combination with miR-1, miR-133, miR-208, miR-499, and Janus kinase (JAK) inhibitors, could convert adult fibroblasts into CMCs both in vitro and in vivo.89) Another group evaluated the functions of CMCs directly converted from fibroblasts, by quantifying robust calcium oscillation, the expression of CMC markers, and spontaneous beating. This study reported that the combination of Hand2, Nkx2-5, Gata4, Mef2c, and Tbx5 represents the most potent arrangement of cardiac lineage-inducing factors.90)
Table 3
Strategy of direct reprogramming for cardiac regeneration

| Strategy used | Mechanisms | Reference |
|---|---|---|
| Delivery of Gata4, Mef2, and Tbx5 (GMT) |
• Plays a key role in early heart development • Exhibit embryonic lethality when mutated |
85) |
| Addition of transcription factors Hand2 and Nkx2-5 with GMT |
• Mesodermal and neural crest-derived structures of the developing heart • Interact with non-coding regions of many genes involved in cardiogenesis • Inhibits cell cycle-inducing genes and enhances cardiac reprogramming |
88) 90) |
| Combination of microRNA and JAK inhibitor |
• Enhances cardiac reprogramming efficiency • Plays a key role in cardiac muscle development and differentiation • Inhibition of fibroblast signatures |
89) |
| Mechanobiology |
• Favorable extracellular environment for reprogramming, e.g., stretch stimulation • Induces changes in the chromatin status and affects cardiac lineage gene expression |
92) |
Mechanobiology is known to play a role in cardiac embryology, diseases, and regeneration,91) but its function in cardiac reprogramming remains unclear. The key factor that determines the success or failure of reprogramming is the extracellular environment. The microenvironment of a mesenchymal stem cell culture can affect its differentiation capacity.92) CMCs derived from hiPSCs exhibit enhanced maturation upon stimulation of their external stretch by a physical stimulant. Transduced fibroblasts seeded on a substrate with microgrooves have also been reported to show improved reprogramming efficiency in CMCs. Another study demonstrated that cells cultured in a 3D hydrogel show improved efficiency of miRNA-mediated direct reprogramming of cells into CMCs, compared to the cells cultured in a conventional 2D culture.93) Although research on direct cardiac reprogramming is continuously evolving, more efforts are still required before this technique can be used in clinical applications. For example, development of safe viral vectors that is applicable in humans is required, and improved efficacy is needed to induce direct cardiac reprogramming from patient-derived adult somatic cells.
CONCLUSION
Stem cell technology and cellular reprogramming are rapidly growing fields that will continue to prove useful in cardiac regenerative therapeutics. The lessons learned from previous studies on traditional PSCs have facilitated the application of hiPSCs in CVD modeling and clinical cell transplantation. The subsequent development of technology to induce the direct differentiation of adult somatic cells into CMCs using defined factors has allowed for ethical problems and the low potential of teratoma formation to be circumnavigated. This represents the overcoming of a major hurdle to the use of iPSCs as primary cells in cell transplantation. The remaining challenges for the use of PSCs and direct reprogramming in clinical applications are as follows: (1) the efficiency of reprogramming must be improved for clinical applications; (2) the safety of direct reprogramming must be demonstrated in large animals, such as pigs and others, before it can be applied to humans; and (3) the molecular mechanisms underlying these cellular events must be deciphered precisely and investigated in more detail. Currently, direct cardiac remodeling events have been mainly studied in myocardial infarction models. Their efficacy should be tested in other CVD models, including dilated cardiomyopathy and other myocardial diseases.
 XML Download
XML Download