

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
The gut microbiome is a collection of microorganisms that live in the human digestive system in a symbiotic relationship with their host. The gut microbiome plays critical role in glucose metabolism and low-grade inflammation associated with obesity and related metabolic disorders by the transport of lipopolysaccharide-induced extracellular vesicles (EVs) [1]. Gut microbial communities are also associated with obesity. The composition of the microbial diversity differs between lean subjects and obese subjects. In overweight and obese subjects, the abundance of organisms belonging to the phylum Firmicutes is significantly increased, while that of organisms belonging to the genus Bifidobacterium and the phylum Bacteroidetes is significantly decreased; thereby, the ratio of Firmicutes to Bacteroidetes is changed [2]. The composition of the gut microbial community mainly depends on the host, but it can also be altered by exogenous and endogenous factors, including changes in dietary habits, health conditions, and surgery [3]. Bariatric/metabolic surgery for treating simple obesity or morbid obesity with type 2 diabetes mellitus (T2DM) is the representative example. It can cause physiological changes in food choices and preferences, reduced food intake, and malabsorption, which results in significant alterations in the gut microbial community of the alimentary tract [45]. However, recent data have reported that the gut microbiome is not fully restored even after bariatric surgery, and the specific mechanism underlying gut microbiome modifications by bariatric/metabolic surgery has not been elucidated.
Bisphenol A (BPA) is an organic synthetic compound that is popularly used for manufacturing polymers such as polycarbonate (PC), epoxy resins, and polyvinyl chloride (PVC) [6]. Many food items are contaminated with BPA, and this is true especially in canned foods or beverage bottles because foods are inevitably in contact with internal coating materials of food packages containing PC, epoxy resins, or PVC to store food which have a waterproof effect [7]. When the BPA-contaminated food is digested, BPA is first rapidly metabolized and absorbed from the gastrointestinal tract then gets trapped in adipose tissue due to its lipophilic nature [8]. BPA accumulation in the body has been recently deemed as an environmental obesogen [9]. The results from the Canadian Health measure survey showed that urinary BPA was positively associated with body mass index (BMI)-defined obesity, with an odds ratio of 1.54 [10]. In addition, several epidemiological studies showed that long-term BPA exposure is positively related to increased T2DM risk [1112]. Provvisiero et al. [13] reported that even exposure to low doses of BPA may increase T2DM by inducing impairment of insulin and glucagon secretion by affecting muscle, hepatic, and adipose cell function, and by triggering an insulin-resistant state.
Recently, the influence of environmental contaminants such as BPA on the gut microbial community has been suggested [14]. Dietary BPA causes significant reduction in the class Clostridia, which is often observed in diabetic patients. However, studies on the effects of BPA on gut microbial changes after bariatric/metabolic surgery have not been reported. In an attempt to investigate the influence of BPA on changes in the gut microbiome composition following bariatric/metabolic surgery, the present study adopted a next-generation sequencing approach to characterize the gut bacteria by analyzing the bacterial 16S ribosomal RNA (rRNA) sequences from fecal samples of rats subjected to 2 different diets and 2 surgical procedures; namely, a high-fat diet (HFD) or HFD mixed with BPA, and sleeve gastrectomy (SG) or Roux-en Y gastric bypass (RYGB).
Go to : 
METHODS
This study was approached by the Institutional Animal Care and Ethical Use Committee of the laboratory animal research center of Ajou University Medical Center (No. 2015-0051) and all experiments were conducted in accordance with the Guide for the Care and Use of Laboratory Animals.
Animals
Twenty-four 6-week-old male Wistar rats (160–200 g) were purchased from Orient (Seongnam, Korea) and were housed in polypropylene cages with sterile bedding, standard rodent chow, and glass water bottles. The rats were acclimatized to the laboratory environment for 1 week before the experiment commenced. Rats were then randomly assigned to 2 groups: the HFD group and the BPA group (fed HFD mixed with BPA). The HFD was purchased from Harlan Korea Laboratories, Ltd., (Seoul, Korea) which comprise 4.6 kcal/g, with 19.0% protein, 44.8% fat, and 36.2% carbohydrates. BPA (>99% purity BPA) was purchased from Sigma-Aldrich Chemical Industries (St. Louis, MO, USA). In the BPA group, BPA was blended into the HFD at a concentration of 50 µg/kg, then a pellet was formed again.
Each group (n = 12) was housed individually in single cages at an ambient temperature of 22℃ ± 1℃, the humidity of 60% ± 10%, and a 12-hour light:12-hour darkness cycle. The rats were allowed to eat and drink ad libitum with the assigned diet. After 40 weeks of feeding, the rats in each group were divided into 2 groups and underwent SG and RYGB (Fig. 1A). After 12 weeks from surgery, the rats were sacrificed after 16-hour overnight fasting. Fresh fecal samples were collected at weeks 40 (before the surgery) and postoperative weeks 12 (after the surgery) and stored at –80℃ until use.
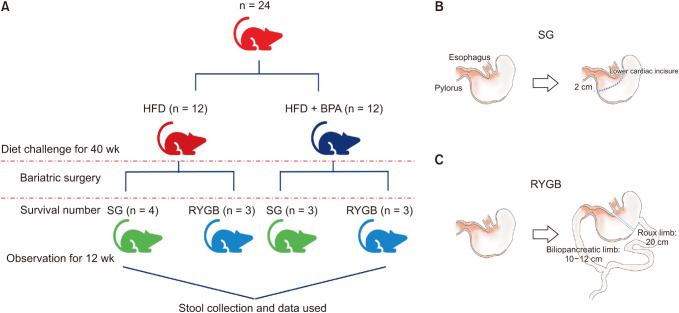 | Fig. 1Experimental design and surgical procedures. (A) Schematic representation of the experimental groups depending on type of diet and surgical procedures. (B) Schematic diagram illustrating sleeve gastrectomy (SG). (C) Schematic diagram illustrating Roux-en Y gastric bypass (RYGB). HFD, high-fat diet; BPA, bisphenol A.
|
Body weight and glucose level measurements, glucose tolerance test
Body weight and food consumption were measured weekly during the experiments. A drop of blood was obtained from the tail vein, and fasting blood sugar (FBS) after 16-hour overnight fasting was measured every 2 weeks using a glucometer (Accu-Chek Performa, Roche, Basel, Switzerland). To assess glucose homeostasis, an intraperitoneal glucose tolerance test (IPGTT) was performed at weeks 40 before the surgery and end of the experiment. After overnight fasting, the rats were injected intraperitoneally with 2 g/kg glucose per body weight (D-(+)-glucose solution, 45% in H2O, sterile filtered, BioXtra Sigma G8769; Sigma-Aldrich); blood glucose level was determined at the indicated time points of 0, 30, 60, 120 minutes.
Surgical techniques
After an overnight fast with freely available water, the rats were anesthetized by respiratory anesthesia using isoflurane. With the rats in a supine position, a 4-cm midline incision was made below the xiphoid. For the SG procedure, after the greater curvature including entire fundus of the stomach (approximately 70%–80% of the gastric volume) was removed using a linear stapler (ECHELON FLEX ENDOPATH stapler, 60-mm white cartilage; Ethicon, Raritan, NJ, USA). Short gastric vessels were then ligated with 4-0 silk (Fig. 1B). For the RYGB procedure, a small gastric pouch was created by transecting the stomach above the limiting ridge after identification of the esophagogastric junction. The Treitz ligament was identified and measured approximately 10 cm distant from where the duodenum and jejunum were transected. After approximately 20-cm length of Roux limb was formed, end-to-side gastrojejunostomy and jejunojejunostomy were made by 6-0 PDS*II (Ethicon) sutures in an interrupt manner (Fig. 1C). The abdominal wall was closed layer by layer using continuous 3-0 silk and 3-0 nylon sutures. Three days after the surgery, all the surgical rats were provided with the assigned diet and water ad libitum.
Extracellular vesicles isolation and DNA extraction from stool samples, processing, and analysis
The stool sample was mixed with phosphate-buffered saline and diluted at a 1:10 ratio (1 g:10 mL) and maintained at 4℃ for 24 hours. After dilution, the sample was centrifuged (10,000 ×g, 10 minutes, 4℃) to remove bacterial fraction and the EV portion was isolated. The supernatants containing EVs were sterilized by filtering through a 0.22-µm filter and 100℃ boiling. The remaining particles and waste were removed by centrifugation at 13,000 rpm for 30 minutes at 4℃. Bacterial DNA was extracted using a DNA isolation kit (DNeasy PowerSoil Kit, QIAGEN, Hilden, Germany) and quantified using a QIAxpert system (QIAGEN).
For microbiome analysis, 16S rRNA gene amplicon metagenomic analysis was performed. The prepared bacterial DNA was used for polymerase chain reaction (PCR) amplification of the 16S rRNA gene using a primer set of 16S_V3_F (5′-TCGGCGATCGATGAGAGACCCT). The PCR product was used to construct the 16S rRNA gene library according to the MiSeq System guidelines (Illumina, San Diego, CA, USA). The 16S rRNA gene library for each sample was quantified using QIAxpert, was pooled isotropically and used for pyrosequencing with the MiSeq System according to the manufacturer’s recommendations.
Paired-end reads that matched the adapter sequences were trimmed via Cutadapt version 1.1.6 [15]. The resulting FASTQ files containing paired-end reads were merged with CASPER and then quality filtered based on the Phred (Q) score-based criteria [1617]. Any reads shorter than 350 bp or longer than 550 bp after merging were also discarded. To identify chimeric sequences, a reference-based chimera detection step was conducted with VSEARCH against the SILVA gold database [1819]. Next, the sequence reads were clustered into operational taxonomic units (OTUs) using VSEARCH with a de novo clustering algorithm under a threshold of 97% sequence similarity. Finally, representative sequences of the OTUs were classified using the Greengenes database (ver. 13_8) with UCLUST (parallel_assign_taxonomy_uclust.py script on QIIME version 1.9.1) under the default parameters [20]. The Chao indices, an estimator of taxa richness per individual, were estimated to measure the diversity in each sample.
Statistical analysis
All data are expressed as the mean ± standard deviation. Statistical analyses were performed using GraphPad Prism 8.0.1 software and the “phyloseq” package of R software ver. 3.4.1 (R Foundation for Statistical Computing, Vienna, Austria). Intergroup comparisons were evaluated with unpaired t-tests for continuous variables, and multiple t-tests (1 per row) were used to compare the microbial compositions. P-values less than 0.05 (2-sided significance testing) were considered statistically significant.
Go to :
RESULTS
Impact of bisphenol A on body weight changes
Before the bariatric/metabolic surgery, 1 rat in the BPA group died of unclear causes in the 38th week and was excluded from the analyses. The body weight changes showed a similar trend, increased from about 200 to 800 g, regardless of BPA exposure. However, the rats in the BPA group gained more body weight than those in the HFD group (624.3 ± 172.0 g vs. 610.6 ± 167.1 g, P < 0.001) (Fig. 2A).
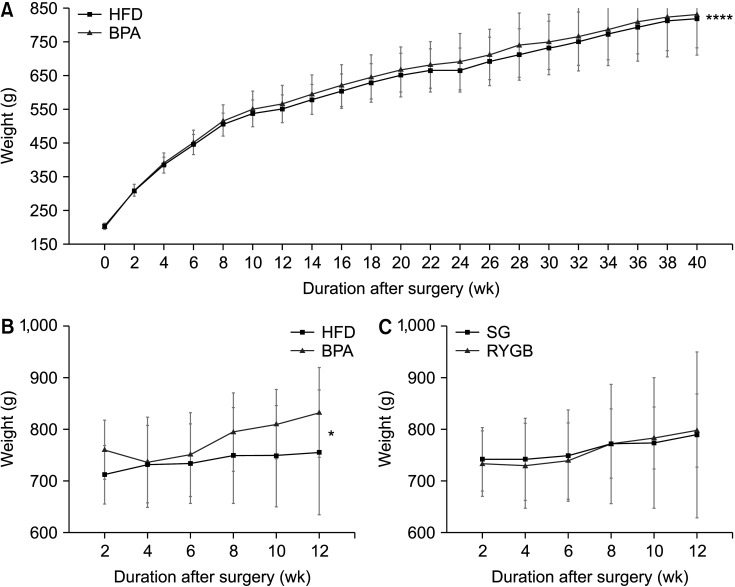 | Fig. 2Body weight changes of rats before and after the surgery. (A, B) Body weight changes between the high-fat diet (HFD) and bisphenol-A (BPA) groups before (A) and after (B) the surgery. (C) Body weight changes between the sleeve gastrectomy (SG) and Roux-en Y gastric bypass (RYGB) subgroups after the surgery.
|
After surgery, 13 rats survived until 12 weeks postoperatively, though fecal samples were collected from 10 rats; 2 each were in the HFD-SG and BPA-SG subgroups and 3 each were in the HFD-RYGB and BPA-RYGB subgroups. Subgroup analyses according to the different diets and types of bariatric surgery showed that the body weights in the BPA group were significantly higher than those in the HFD group (781.1 ± 37.3 g vs. 738.6 ± 16.0 g, P = 0.028); however, there was no difference between the SG and RYGB subgroups (P = 0.896) (Fig. 2B, C).
Long-term exposure to bisphenol A may induce glucose intolerance
At week 40, after a 16-hour overnight fast, FBS was significantly higher in the BPA group than that in the HFD group (90.5 ± 5.4 mg/dL vs. 87.2 ± 4.2 mg/dL, P = 0.033) (Fig. 3A). After the surgery, the FBS of the BPA group was relatively higher than that of the other groups although there was no statistical significance between the HFD and BPA groups or between the SG and RYGB groups (Fig. 3B, C). In the IPGTT measured in week 40, the peak blood sugar levels were higher in the BPA group, especially at 30 and 60 minutes (Fig. 3D) than those in the HFD group; however, the difference did not reach statistical significance. This trend was also observed in the IPGTT measured in week 12 after surgery (Fig. 3E). As shown in Fig. 3F, there were similar blood sugar trends in the SG and RYGB groups, although the difference was not statistically significant.
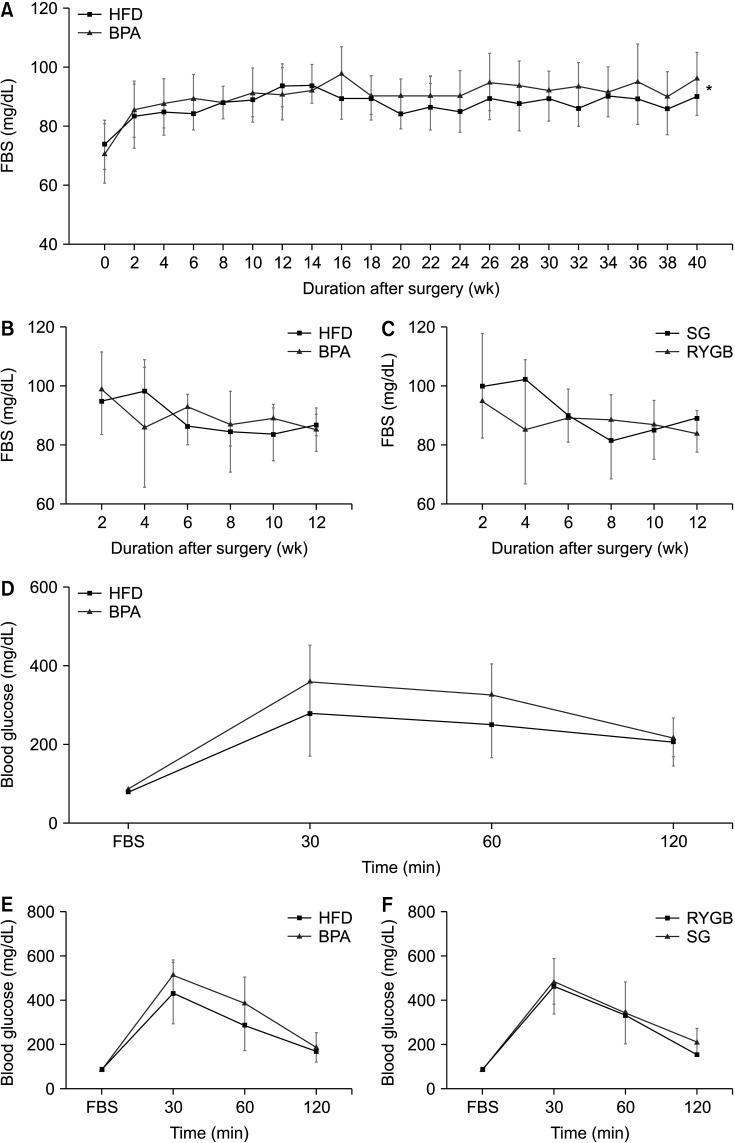 | Fig. 3Sixteen-hour overnight fasting blood sugar (FBS) and glucose level changes in intraperitoneal glucose tolerance test (IPGTT). (A, B) The levels of 16-hour overnight FBS changes in the high-fat diet (HFD) and bisphenol-A (BPA) groups before (A) and after (B) the surgery. (C) Sixteen-hour overnight FBS changes in the sleeve gastrectomy (SG) and Roux-en Y gastric bypass (RYGB) subgroups after the surgery. (D) Blood glucose level changes after IPGTT in the HFD and BPA groups before (D, 40th week) and after (E, 12th week) the surgery. (F) Blood glucose level changes after IPGTT test in the SG and RYGB subgroups after surgery (12th week).
|
Changes in the microbial communities in stool bacteria
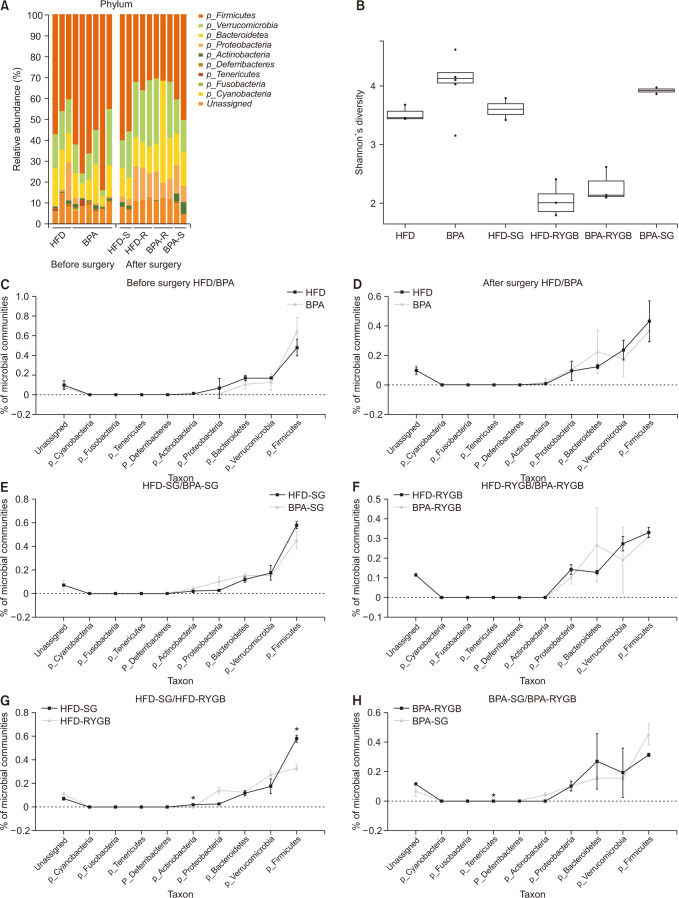
A comparative analysis of the microbial communities before and after surgery among the different subgroups was conducted. A distribution-stacked map of the taxonomic compositions of each rat from the respective groups was constructed at the phyla level (Fig. 4A). The Shannon index showed a gradual decreasing tendency in microbial diversity after surgery; however, there were no remarkable differences (Fig. 4B). Before surgery, there were no significant differences between the proportions of the microbial community compositions in the HFD and BPA groups (Fig. 4C), and the most abundant phyla were Firmicutes, Verrucomicrobia, Bacteroidetes, and Proteobacteria. After surgery, the compositional proportions resembled that from before surgery (Fig. 4D). There were no statistically significant differences in the results from the animals who received the same surgical procedure but different diets (HFD-SG vs. BPA-SG and HFD-RYGB vs. BPA-RYGB) (Fig. 4E, F). The abundance of the phylum Actinobacteria was significantly lower in the HFD-RYGB subgroup relative to that in the HFD-SG subgroup (P < 0.001) (Fig. 4G), and the abundance of the phylum Tenericutes was significantly lower in the BPA-RYGB subgroup relative to that in the BPA-SG group (P < 0.001) (Fig. 4H).
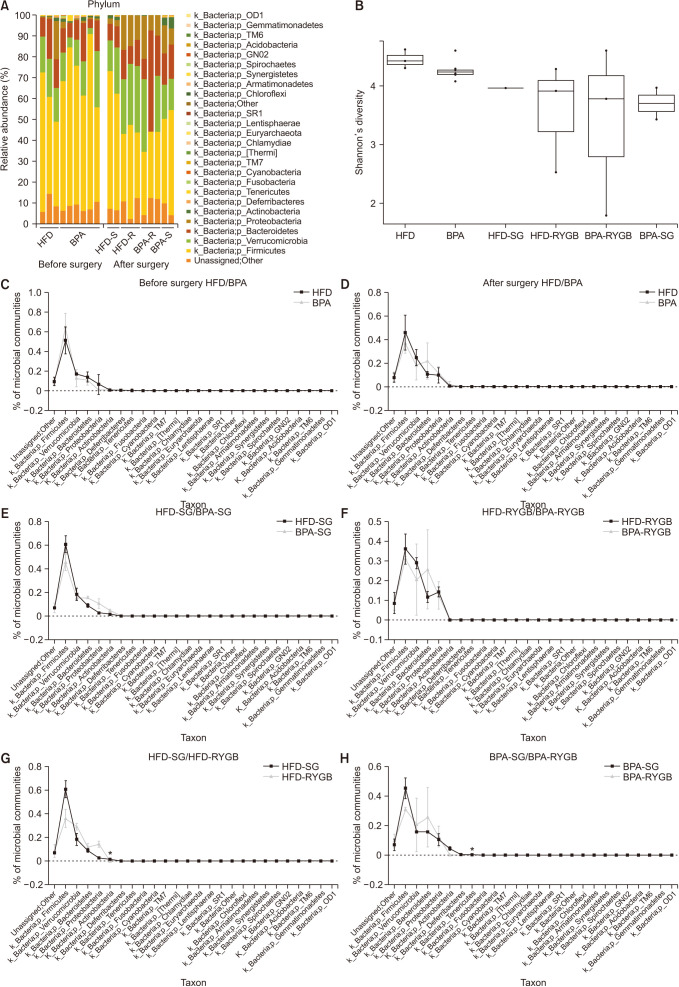 | Fig. 4Changes in the phyla proportions and diversity in the microbial communities in stool bacteria. (A) Relative phyla abundances in the microbial communities of the different groups before and after the surgery. (B) Shannon’s diversity changes among the different groups. (C) Comparison of the microbial community compositions in the high-fat diet (HFD) and bisphenol-A (BPA) groups before the surgery. (D) Comparison of the microbial community compositions in the HFD and BPA groups after the surgery. (E) Comparison of the microbial community compositions in the HFD-sleeve gastrectomy (SG) and BPA-SG subgroups after the surgery. (F) Comparison of the microbial community compositions in the HFD-Roux-en Y gastric bypass (RYGB) and BPA-RYGB subgroups after the surgery. (G) Comparison of the microbial community compositions in the HFD-SG and HFD-RYGB subgroups after the surgery. (H) Comparison of the microbial community compositions in the BPA-SG and BPA-RYGB subgroups after the surgery.
|
Changes in the microbial communities in stool extracellular vesicles
The same method was used to analyze the composition of the microbial communities in stool EVs. A distribution-stacked map of the taxonomic compositions was constructed at the phyla level (Fig. 5A). The Shannon index showed a decreasing tendency in microbial diversity after surgery, especially in the HFD-RYGB and BPA-RYGB subgroups; however, there were no remarkable differences (Fig. 5B). Before surgery, there were no statistically significant differences in the compositional proportions of the microbial communities of the HFD and BPA groups (Fig. 5C), and the most abundant phyla in the gut microbiome were Firmicutes, Verrucomicrobia, Bacteroidetes, and Proteobacteria. After surgery, there was also no statistically significant difference between the HFD and BPA groups (Fig. 5D); furthermore, there were no statistically significant differences among the animals that received the same surgical procedure but different diets (HFD-SG vs. BPA-SG and HFD-RYGB vs. BPA-RYGB) (Fig. 5E, F). The abundances of Actinobacteria and Firmicutes were significantly lower in the HFD-RYGB subgroup than in the HFD-SG subgroup (P = 0.005 and P = 0.002, respectively) (Fig. 5G), and the abundance of Tenericutes was significantly lower in the BPA-RYGB subgroup than in the BPA-SG subgroup (P = 0.003) (Fig. 5H).
 | Fig. 5Changes in the proportions and diversity of the microbial communities in stool extracellular vesicles. (A) Relative phyla abundances in the microbial communities of the different groups before and after the surgery. (B) Shannon’s diversity changes among the different groups. (C) Comparison of the microbial community compositions in the high-fat diet (HFD) and bisphenol-A (BPA) groups before the surgery. (D) Comparison of the microbial community compositions in the HFD and BPA groups after the surgery. (E) Comparison of the microbial community compositions in the HFD-sleeve gastrectomy (SG) and BPA-SG subgroups after the surgery. (F) Comparison of the microbial community compositions in the HFD-Roux-en Y gastric bypass (RYGB) and BPA-RYGB subgroups after the surgery. (G) Comparison of the microbial community compositions in the HFD-SG and HFD-RYGB subgroups after the surgery. (H) Comparison of the microbial community compositions in the BPA-SG and BPA-RYGB subgroups after the surgery.
|
Go to :
DISCUSSION
The present study illustrates that BPA may encourage weight gain, thus limiting the weight loss resulting from bariatric surgery. In addition, BPA negatively affected glucose tolerance and increased the overnight FBS level. However, BPA did not influence the compositional proportions or the diversity in the microbial communities after bariatric/metabolic surgery.
BPA has been at least partially linked to obesity, and it is commonly regarded as a putative environmental obesogen based on some evidence from animal studies and human epidemiologic studies [21]. BPA might stimulate adipocyte differentiation and proliferation and induce adipocyte hypertrophy, thus facilitating weight gain [22]. In the present study, the rats in the BPA group gained significantly more body weight compared with those in the HFD group. This result is consistent with other reports. Rubin and Soto [23] reported that relative to the animals in the control group, the animals perinatally exposed to BPA showed increased body weight. A cross-sectional study of school children revealed that higher urine BPA concentrations were significantly associated with increasing BMI values in all of the subjects; thus, BPA exposure increased BMI in school children [24].
In our study, long-term exposure to dietary BPA led to increased overnight FBS levels, even after bariatric/metabolic surgery. In addition, IPGTT showed decreased glucose tolerance both before and after surgery in the BPA group, although it was not statistically significant due to small sample size. These findings might be due to the fact that BPA exposure alters glucose metabolism. Alonso-Magdalena et al. [25] reported that BPA exposure aggravated insulin resistance and was associated with decreased glucose tolerance. Furthermore, perinatal BPA exposure resulted in either increased β-cell mass or changes in insulin secretion and insulin sensitivity [26]. Meanwhile, the subanalysis between the SG and RYGB procedures showed that blood glucose levels remained similar in both groups (Fig. 3F); this implies that bariatric surgery was beneficial in improving glucose tolerance, increasing insulin sensitivity, or hepatic glycogen deposition, regardless of BPA exposure [27].
Evidence shows that dietary habits can alter microbial communities. In this study, we aimed to investigate the effect of BPA on microbial communities change that occur after bariatric/metabolic surgery. A previous study reported that a standard diet with BPA can induce a similar shift in the structure of microbial communities as HFD; significant decreases in the phylum Firmicutes, of which most of the 16S rRNA belong to the class Clostridia [14]. However, we failed to find any difference in microbial communities between the HFD group and the BPA group, before and after surgery. This discrepancy might be due to differences in feeding method and BPA concentrations. In a previous study [14], the rats were exposed to BPA via their water supply at a concentration of 120 µg/mL, whereas BPA was blended with HFD at a concentration of 50 µg/kg (≒5 × 10-5 µg/mg).
The compositions of the microbial communities in the stool bacteria and EVs differed in animals undergoing either the SG or the RYGB procedure. We found that the compositions of the microbial communities following RYGB were typified by a markedly lower proportion of Actinobacteria in the HFD group and a significantly lower proportion of Tenericutes in the stool bacteria and EVs in the BPA group, compared with their levels in the SG group, and the phylum Firmicutes was significantly lower in the stool EVs in the HFD-RYGB subgroup compared with that in the HFD-SG subgroup. Interestingly, in the stool bacteria, the phylum Firmicutes did not markedly decrease in the HFD-RYGB group compared with that in the HFD-SG group. It might be due to the fact that EVs can be formed from both active and dead stool bacteria; therefore, it did show a decreasing trend, which might explain this observation.
This was the first pilot study to investigate the effects of BPA on body weight and microbiota after bariatric surgery. The strength of the present study includes the long duration (40 weeks of dietary challenge before the bariatric/metabolic surgery and 12 weeks after the surgery), the route of BPA exposure (oral), and induction of glucose intolerance using a low intake dose of BPA (50 µg/kg). There are several limitations in the present study. Firstly, the numbers of rats monitored after the surgery were relatively small due to the death of animals from postoperative complications such as anastomotic leak. We developed a standardized protocol for surgical procedures, but survival rate was lower than expected, especially in RYGB (37.5%). Fortunately, although several results were not statistically significant, we did succeed in detecting some trends. Secondly, the present study aimed to evaluate whether exposure to BPA affect alterations of gut microbial communities before and after bariatric/metabolic surgery; thereby, changes in insulin or related hormone levels, insulin signaling pathway, and islet cell morphology of the pancreas were not investigated in the present study. The impact of BPA on metabolic disease such as T2DM would be elucidated in further study. Lastly, the surgical procedures differ slightly from those in humans due to the anatomical differences. For instance, we made a small pouch in the RYGB procedure where possible, yet the gastric fundus was preserved for the anastomosis. This might affect the study’s outcomes whereby the results would be interpreted carefully.
Taken together, our results highlight that BPA is a risk factor for obesity and may contribute to glucose intolerance, but it did not affect alterations of gut microbiome after bariatric/metabolic surgery. Further studies to determine the influence of environmental contaminants on the gut microbiome in relation to metabolic disease would be needed.
Go to :
 XML Download
XML Download