

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Natural killer (NK) cell-based cancer immunotherapy is effective because NK cells can kill various tumor cells without deliberate immunization, activation, or prior sensitization [1-3]. Unlike cytotoxic T lymphocytes, which can only be used in an autologous setting because they may cause graft-versus-host disease (GvHD), NK cells can be used in allogeneic settings. Therefore, they are useful off-the-shelf therapeutics [4, 5].
For allogeneic NK cell-based cancer immunotherapy, diverse sources of NK cells, including peripheral blood (PB), umbilical cord blood (UCB), bone marrow, cell lines, human embryonic stem cells, and induced pluripotent stem cells, can be used [6-8]. Among these, UCB is an effective NK cell source; it contains a more distinct NK cell progenitor population that may have a better proliferative capacity than the PB population [9]. However, two major impediments to UCB NK cell immunotherapy are the relatively low number of NK cells in a single UCB unit and limited UCB volume. Thus, the development of effective expansion methods to acquire an adequate number of NK cells for UCB NK cell therapy is essential.
Diverse culture conditions, including cytokine combinations, co-stimulatory molecules, and various genetically engineered feeder cells, such as K562-41BBL-mbIL15, K562-mbIL21, K562-OX40L, and K562-OX40L-mbIL18/21 cells, have been investigated to identify conditions that improve ex vivo NK cell expansion [10-13]. In addition, various culture media and supplements are known to influence NK cell expansion and function [14, 15]. Roswell Park Memorial Institute (RPMI) 1640 culture medium supplemented with fetal bovine serum (FBS) (RPMI complete; RC) is widely used as a standard culture medium for NK cell expansion [10, 11, 16]. Pierson, et al. [15] reported that Dulbecco’s modified Eagle’s medium (DMEM) containing Ham’s F12 Nutrient Mix and supplemented with human AB serum, GlutaMAX, β-mercaptoethanol, ethanolamine, and ascorbic acid (DMEM/F12+Supplements; DS) further enhanced the proliferative capacity of NK cells compared with RC. Their study verified that medium supplementation with human serum considerably supports NK cell proliferation. Moseman, et al. [14] extensively compared commercially available serum-free media (X-VIVO, SCGM, OpTimizer, ABS-011, TexMacs, and AIM V) and reported that AIM V supplemented with Immune Cell Serum Replacement (ICSR) (AIM V+ICSR; AI) enhanced the proliferative capacity and functionality of NK cells. Although there is no consensus on the best medium for NK cell expansion, it is clear that the culture medium and wide range of supplements markedly affect NK cell expansion and functionality. Therefore, comparative analyses of the effects of culture media and supplements are needed.
In this study, we selected three media, i.e., an FBS-supplemented medium (RC), a human serum-supplemented medium (DS), and serum-free medium (AI), and compared their effects on the expansion rate and functionality of UCB NK cells cultured under stimulation with K562-OX40L-mbIL-18/21 feeder cells and IL-2 and IL-15.
MATERIALS AND METHODS
UCB samples
This prospective study was conducted between March 2020 and April 2021. Anonymized healthy UCB samples were obtained from the Seoul Metropolitan Government Public Cord Blood Bank (Allcord) upon approval. This study was approved by the Institutional Review Board of Samsung Medical Center, Seoul, Korea (approval No. 2020-03-057). All experiments were conducted in accordance with relevant guidelines and regulations.
Isolation of UCB mononuclear cells
UCB mononuclear cells were isolated by density gradient centrifugation (25 minutes, 37°C, 500×g) using Ficoll-Hypaque (d=1.077, Lymphoprep; Axis-Shield, Oslo, Norway) and washed twice with phosphate-buffered saline (Welgene, Gyeongsan, Korea). All protocols were performed in accordance with relevant guidelines and regulations.
Culture media and supplements
Isolated UCB mononuclear cells were cultured in RPMI 1640 (Hyclone Laboratories, Logan, UT, USA), a mixture of DMEM/Ham’s F-12 (Gibco, Grand Island, NY, USA; 2:1 v/v), and AIM V serum-free (Gibco) basal medium. RPMI 1640 medium was supplemented with 10% heat-inactivated FBS (Gibco), 100 U/mL penicillin, 100 μg/mL streptomycin (Lonza, Walkersville, MD, USA), and 4 mmol/L glutamine (Gibco) (RC). DMEM/Ham’s F-12 (2:1) was supplemented with 10% heat-inactivated human AB serum (Thermo Fisher Scientific, Waltham, MA, USA), 25 µM beta-mercaptoethanol (Thermo Fisher Scientific), 50 µmol/L ethanolamine (Sigma-Aldrich, St. Louis, MO, USA), 20 µg/L ascorbic acid (Sigma-Aldrich), and 5 ng/L sodium selenite (Sigma-Aldrich) (DS). AIM V was supplemented with 5% ICSR (Life Technologies, Grand Island, NY, USA).
Cell lines
K562 and Raji cells were acquired from the American Type Culture Collection (Manassas, VA, USA) and were cultured in RC medium at 37°C in a humidified 5% CO2 incubator. K562-OX40L-mbIL18/21 feeder cells were generated by transducing parental K562 cells with lentiviral vectors encoding OX40L, IL-18, and IL-21 [13].
NK cell expansion
To eliminate donor-to-donor variation in NK cell expansion, isolated UCB mononuclear cells from each donor were split into same-size aliquots for paired expansions in each culture medium assessed. The numbers of donors used for NK cell expansion are indicated in the figure legends. The isolated UCB mononuclear cells were expanded in RC, DS, and AI. K562-OX40L-mbIL18/21 feeder cells were irradiated with 100 Gy of radiation and co-cultured with the UCB mononuclear cells in a 24-well plate. In the first week, 10 U/mL recombinant human IL-2 (PeproTech, Rocky Hill, NJ, USA) was added to each culture. After one week, the IL-2 level was increased to 100 U/mL and an additional 5 ng/mL of soluble IL-15 was added to each culture, as previously described [13]. UCB NK cells were stimulated with K562-OX40L-mbIL18/21 cells on days 0, 7, 14, and 21. The culture medium was replaced every 2-3 days, and the cells were continuously cultured until day 28.
In the culture medium shifting experiment, the culture medium was shifted on day 14. During the shift, AI was added to the DS plate, and DS was added to the AI plate, along with 100 U/mL IL-2 (PeproTech) and 5 ng/mL IL-15 (PeproTech). Expanded UCB NK cells were harvested on day 28 for further experiments. Cell expansion is presented as “fold expansion,” which was calculated by dividing the total number of CD56+/CD3– live NK cells in each week by the number of CD56+/CD3– NK cells on day 0.
Cytokines and antibodies
Recombinant human IL-2 and IL-15 (PeproTech) were used to expand the NK cells. Allophycocyanin (APC)-Cy7-conjugated anti-human CD3, phycoerythrin (PE)-Cy7-conjugated anti-human CD56, PE-Cy5-conjugated CD56, and fluorescein isothiocyanate (FITC)-conjugated CD3 (eBioscience, San Diego, CA, USA) were used to measure the purity of NK cells (CD56+/CD3–). Pacific blue-conjugated anti-human CD16, Pacific blue-conjugated anti-human NKp46 (eBioscience), APC-conjugated anti-human DNAM-1, APC-conjugated anti-human NKG2A, and PE-conjugated anti-human NKp30 (BD Biosciences, Franklin Lakes, NJ, USA) were used to measure UCB NK cell receptor levels.
Flow cytometry-based functional assays
For functional assays, expanded NK cells were harvested on days 14 and 28 to measure their cytotoxicity and antibody-dependent cellular cytotoxicity (ADCC) against target tumor cells (K562 and Raji cells) by flow cytometry using carboxyfluorescein diacetate succinimidyl ester (CFSE; Life Technologies) staining of the target cells, as previously described [12, 13]. Briefly, K562 and Raji target cells were stained with 0.5 μM CFSE in fluorescence-activated cell sorting (FACS) buffer at 37°C for 10 minutes and then washed twice with RPMI medium. For the ADCC assay, Raji cells were stained with 0.5 μM CFSE in FACS buffer at 37°C for 10 minutes and then washed twice with RC medium. The CFSE-stained Raji cells were then supplemented with 5 μg/mL rituximab. The target cells (2×104) were transferred into a 96-well U-bottom plate in triplicate and mixed with various numbers of expanded NK cells (0.5:1, 1:1, and 2:1 effector-to-target ratios). The plates were centrifuged at 400×g for 3 minutes and then incubated for 4 hours for functional analysis and for 30, 90, 150, and 240 minutes for the time-course killing assay at 37°C in a 5% CO2 incubator. After the incubation, the co-incubated cells were transferred to FACS tubes. One microliter of 1 mg/mL propidium iodide (Sigma-Aldrich) was added to each tube before data acquisition. All functional assay data were acquired on a FACSVerse instrument (BD Biosciences) and analyzed using the Kaluza software version 2.1. The numbers of donors used in the NK cell functional assays are indicated in the figure legends.
Live cell imaging for cytotoxicity assay
Live imaging for cytotoxicity was conducted as previously described [17]. Briefly, clean coverslips (18×18 mm) were treated with air plasma (100 W, Femto Science, Hwaseong, Korea) for 1 minute and coated with a protein solution containing fibronectin (2.27 mM, Sigma) and anti-CD44 (0.25 mg/mL, clone no. IM7, eBioscience) at 37°C for 30 minutes. K562 cells were stained with 1 mM CellTrace Far Red (Invitrogen) according to the manufacturer’s instructions. The stained K562 cells (0.1×106 cells) and NK cells (0.01×106 cells) were mixed and seeded on the antibody-functionalized coverslips. The samples were then centrifuged to capture the cancer cells from the surface (400×g, 3 minutes).
Fluorescence microscopy
A modified Olympus IX 83 epi-fluorescence microscope equipped with a 40× (UPlanFL N, NA=1.30) objective lens and a Zyla 4.2 sCMOS camera (Andor) was used for imaging experiments. For fluorescence imaging, a U-LH75XEAPO Xenon lamp and 75 W (Olympus) and Cy5 (EX BP 620/60, BS 660, and EM BP 770/75) filter sets were used. The microscope was automatically controlled by a micro-manager, and the stages were equipped with a Chamlide TC incubator system (Live Cell Instrument, Seoul, Korea) to maintain cell culture conditions (37°C, 5% CO2). The images acquired were analyzed and processed using the ImageJ software (NIH, Bethesda, MD, USA).
Intracellular component analysis
To measure granzyme B and perforin levels, day-28 unstimulated expanded UCB NK cells were harvested and stained with APC-Cy7-conjugated anti-human CD3 and PE-Cy5-conjugated anti-human CD56 (eBioscience/BD Biosciences) on ice for 15 minutes. After surface receptor staining, the cells were washed, permeabilized, and fixed using a Cytofix/Cytoperm kit (BD Biosciences) according to the manufacturer’s instructions. Then, the cells were stained with FITC-conjugated anti-human granzyme B and PE-conjugated anti-human perforin (BD Biosciences) on ice for 30 minutes. Finally, the cells were washed and analyzed using the FACSVerse cell analyzer.
Statistics
Differences between the groups in terms of the purity, fold expansion, receptor expression, and cytotoxicity of UCB NK cells were statistically analyzed using two-tailed Student’s paired t-test for two groups and one-way ANOVA for more than two groups, using GraphPad Prism 7 (GraphPad Software, San Diego, CA, USA). P≤0.05 was considered significant.
RESULTS
The culture medium substantially affects the expansion rate and cytotoxicity of NK cells
To investigate the role of the culture medium in NK cell expansion, UCB mononuclear cells were cultured with K562-OX40L-IL18/21 feeder cells in three different culture media: RC, DS, and AI. The fold expansion and purity of each culture were checked weekly until day 28 (Fig. 1A, B). Compared with NK cells cultured in RC, those cultured in DS showed a remarkable increase in fold expansion between days 21 and 28. NK cells cultured in AI also exhibited an increased proliferation rate but not as high as that of cells cultured in DS (Fig. 1A). NK cell purity was not as substantially influenced by the culture medium as fold expansion: NK cells cultured in RC and DS exhibited a slightly higher purity (percentage of CD56+/CD3– cells) than those cultured in AI (Fig. 1B). The fold expansion of NK cells cultured in DS on day 28 was nearly five times greater (38,240) than that of NK cells cultured in RC (8,124), suggesting that an appropriate culture medium is critical for cost-efficient NK cell production.
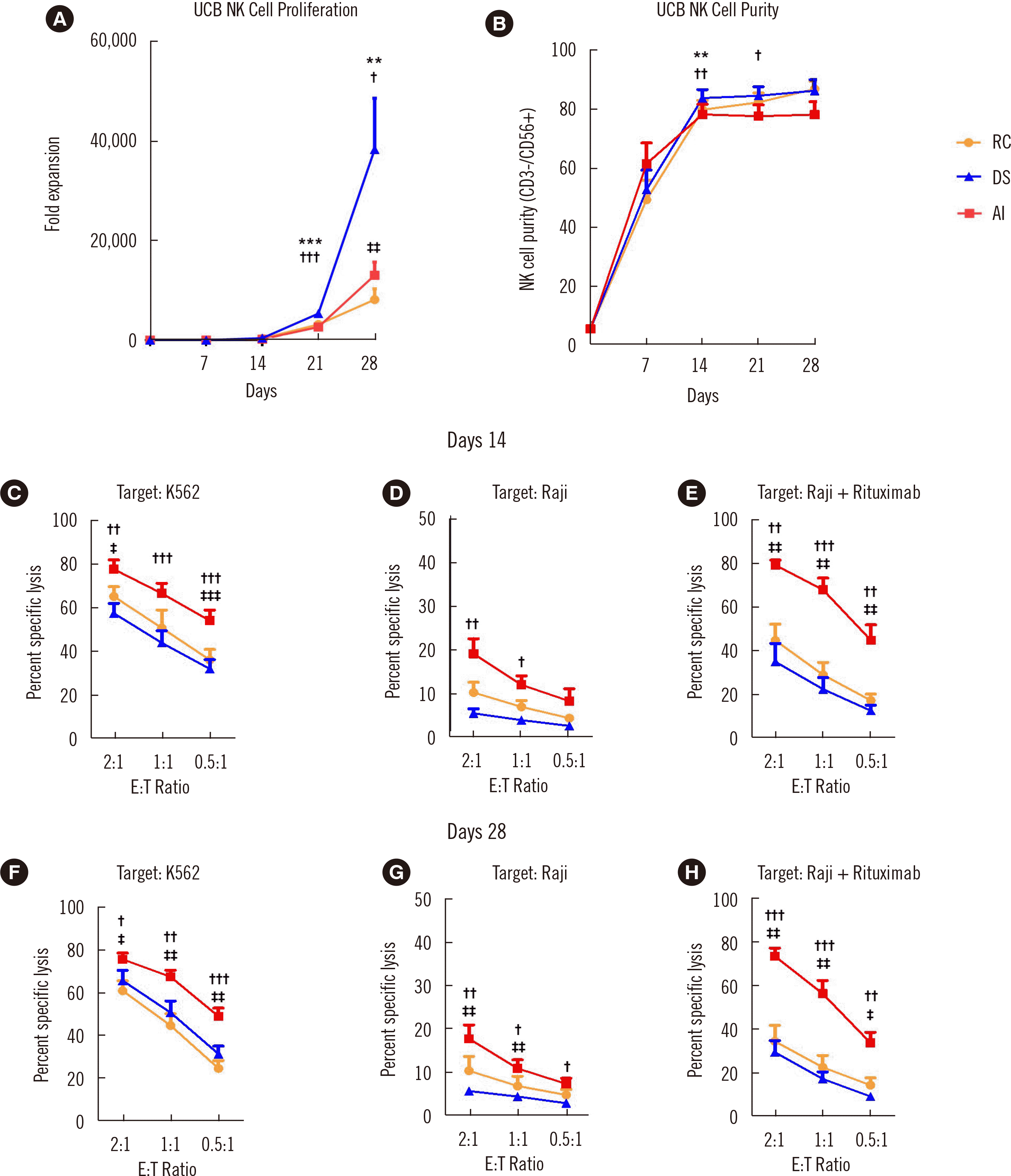
Fig. 1
The culture medium substantially affects NK cells expansion and functions. (A) Fold expansion of UCB NK cells cultured in RC (orange), DS (blue), or AI (AIM V + ICSR) (red) culture media (N=15). (B) Purity (CD56+/CD3– NK cells) of UCB NK cells cultured in RC, DS, or AI (N=15). (C–E) Flow cytometry-based assay of the cytotoxicity of UCB NK cells cultured in RC, DS, or AI media toward K562 cells (N=8) (C), Raji cells (N=6) (D), and Raji cells+rituximab (N=7) (E) on day 14. Cytotoxicity assay of UCB NK cells toward K562 cells (N=8) (F), Raji cells (N=6) (G), and Raji cells+rituximab (N=7) (H) on day 28. Data on fold expansion and purity are the mean and SEM. *,†,‡P<0.05; **,††,‡‡P<0.01; ***, †††, ‡‡‡P<0.001; *DS vs. RC; †DS vs. AI; ‡AI vs. RC.
Abbreviations: RC, RPMI medium + FBS; DS, DMEM + Ham’s F12 + various supplements; AI, AIM V + ICSR; UCB, umbilical cord blood; NK, natural killer; RPMI, Roswell Park Memorial Institute; FBS, fetal bovine serum; DMEM, Dulbecco’s modified Eagle’s medium; ICSR, Immune Cell Serum Replacement.
![]()
In addition to fold expansion, the anti-tumor activity of NK cells is critical for NK cell-based adoptive cell cancer immunotherapy. We compared the anti-tumor activities of NK cells expanded in the different media using direct cytotoxicity and ADCC assays on days 14 and 28. K562 (chronic myeloid leukemia) and Raji (B-cell lymphoma) cells were used as target cells, and rituximab was added to the Raji cells. Interestingly, NK cells cultured in AI exhibited significantly higher cytotoxicity on both days 14 and 28 than those cultured in RC and DS under all conditions (Fig. 1C–E for day 14 and Fig. 1F–H for day 28). ADCC was nearly twice as high in NK cells cultured in AI than in those cultured in RC and DS (Fig. 1E, H).
We tested batch-to-batch variation in the proliferation rate and cytotoxicity of NK cells cultured in two representative media. The remarkable proliferation-promoting effect of DS and the lower functionality of NK cells cultured in DS than of those cultured in AI were consistently observed for different human serum batches. Depending on the serum batch, NK cells cultured in DS showed slightly different fold expansion (Supplemental Data Fig. S1A–C), cytotoxicity (Supplemental Data Figure S1D–F), and ADCC (Supplemental Data Fig. S1G–I); however, the trends of higher fold expansion and lower killing potency than those of NK cells cultured in AI remained.
The culture medium affects receptor expression levels in NK cells
To identify the potential cause of the differences in functionality induced by the different culture media, cytotoxic effector molecules (granzyme B, perforin), activating receptors (DNAM-1, NKG2D, and CD16), natural cytotoxic receptors (NKp46 and NKp30), and an inhibitory receptor (NKG2A) were analyzed using flow cytometry (Fig. 2A–C). Although NK cells cultured in AI exhibited superior cytotoxicity to those cultured in the other media, their granzyme B and perforin expression levels were comparable with those of NK cells cultured in RC. Similar patterns were observed for the activating receptors and natural cytotoxic receptors: NK cells cultured in AI expressed comparable (DNAM-1, NKG2D, CD16, and NKp30) or significantly lower (NKp46) receptor levels than those cultured in RC or DS. NKG2A expression was the lowest in NK cells cultured in AI.
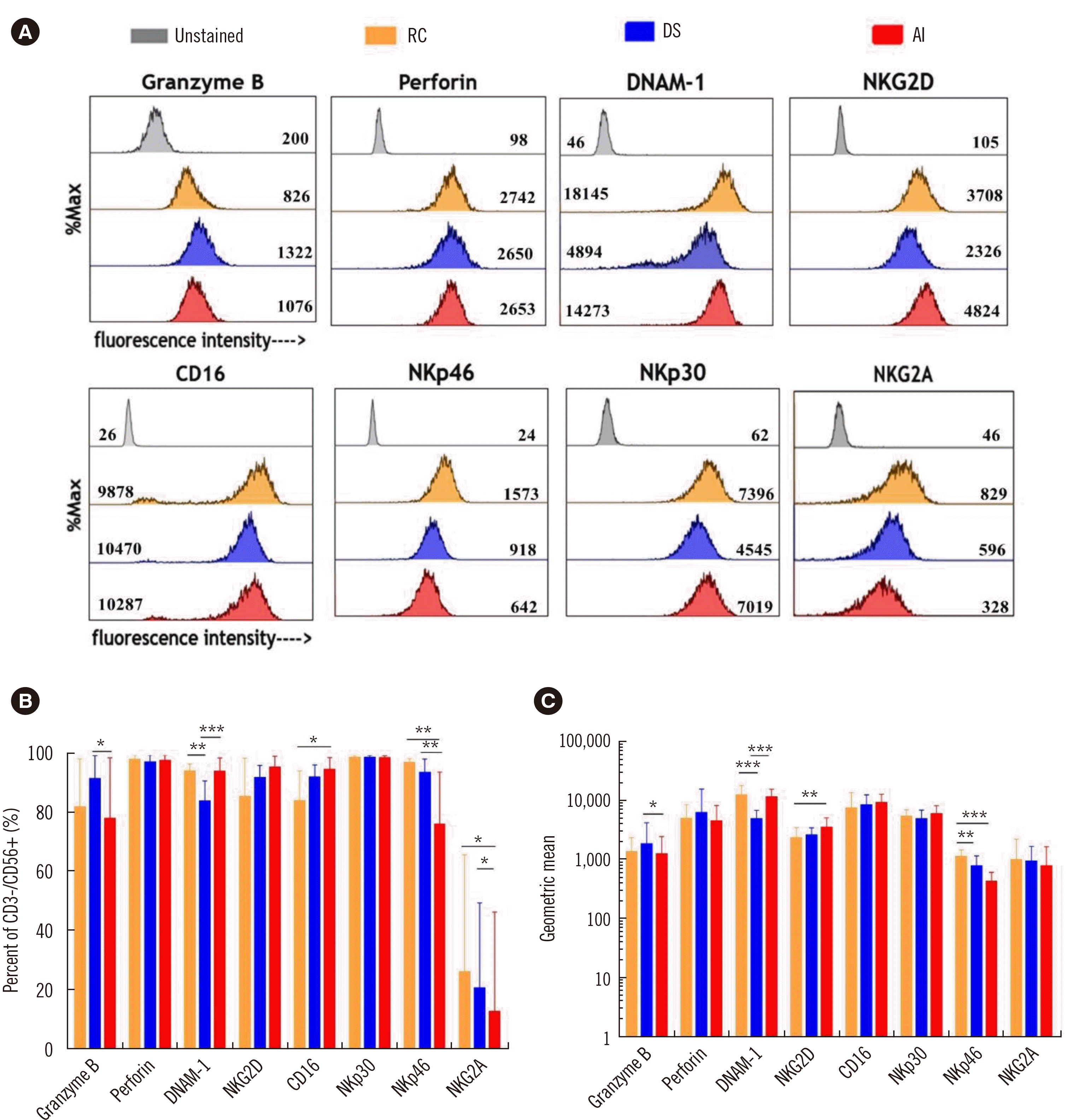
Fig. 2
NK cells cultured in different culture media exhibit different receptor expression levels. (A) Histogram of cytotoxic effector molecule and surface receptor expression of UCB NK cells cultured RC (orange), DS (blue), or AI (red). The geometric means are indicated in the respective histograms. (B) Percent expression levels of cytotoxic effector molecules and surface receptors on day 28 (N=9). (C) Geometric mean expression levels of cytotoxic effector molecules and surface receptors on day 28 (N=9) (*P<0.05; **P<0.01; ***P<0.001).
Abbreviations: RC, RPMI medium+FBS; DS, DMEM+Ham’s F12+various supplements; AI, AIM V+ICSR; UCB, umbilical cord blood; NK, natural killer; RPMI, Roswell Park Memorial Institute; FBS, fetal bovine serum; DMEM, Dulbecco’s modified Eagle’s medium, ICSR, Immune Cell Serum Replacement.
![]()
Culture medium shifting is advantageous for obtaining high numbers of highly cytotoxic NK cells
The extensive proliferation of NK cells cultured in DS and enhanced cytotoxicity of NK cells cultured in AI led us to explore the possibility of obtaining high numbers of highly cytotoxic NK cells by media shifting. NK cells were cultured in DS medium for the first 14 days and then in AI medium for 14 days (DS-AI) or vice versa (AI-DS). Their fold expansion, purity, and cytotoxicity were analyzed on day 28. The fold expansion of NK cells cultured in AI-DS and DS-AI was significantly higher than that of NK cells cultured in AI but lower than that of NK cells cultured in DS (Fig. 3A). The purity of NK cells cultured in DS-AI was significantly higher than that of cells cultured in AI (Fig. 3B). NK cells cultured in DS-AI showed better expansion and purity than those cultured in AI-DS. Interestingly, the medium in which NK cells were cultured in the late phase of expansion was found to determine cytotoxicity (Fig. 3C–E). The cytotoxicity of AI-DS- (or DS-AI)-cultured NK cells was comparable with that of DS- (or AI)-cultured NK cells, indicating that DS-AI- and AI-cultured NK cells exhibited superior cytotoxicity to AI-DS- and DS-cultured NK cells. These results indicate that culture medium shifting is advantageous for recovering NK cytotoxicity (Fig. 3A, C–E).
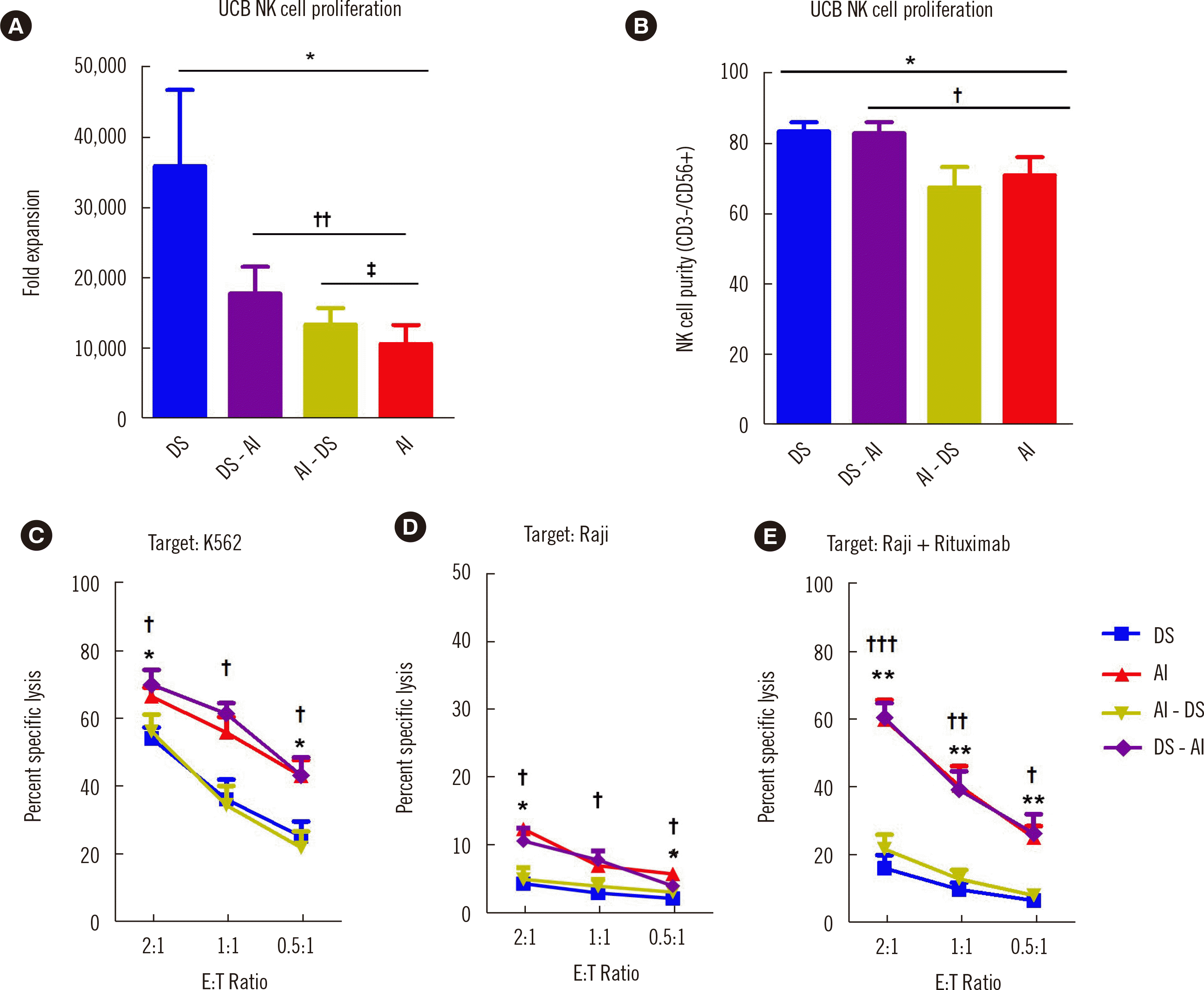
Fig. 3
Culture medium shifting is advantageous for restoring NK cell function. (A) Fold expansion of UCB NK cells cultured in DS (blue), DS-AI (purple), AI-DS (green), or AI (red) (N=8) on day 28. (B) Purity (CD56+/CD3− NK cells) of media-shifted UCB-expanded NK cells cultured in DS (blue), DS-AI (purple), AI-DS (green), or AI (red) (N=11) on day 28. Flow cytometry-based cytotoxicity assay of UCB-expanded NK cells cultured in DS, AI, AI-DS, or DS-AI at the indicated ratios; (C) K562 cells, N=7; (D) Raji cells, N=4; (E) Raji cells + rituximab, N=7 (C–E). *, †P<0.05; **, ††P<0.01; ***, †††P<0.001; *DS vs. AI; †DS vs. DS-AI; ‡AI vs. AI-DS.
Abbreviations: RC, RPMI medium+FBS; DS, DMEM+Ham’s F12+various supplements; AI, AIM V+ICSR; DS-AI, DS shifted to AI on day 14; AI-DS, AI shifted to DS on day 14; UCB, umbilical cord blood; NK, natural killer; RPMI, Roswell Park Memorial Institute; FBS, fetal bovine serum; DMEM, Dulbecco’s modified Eagle’s medium, ICSR, Immune Cell Serum Replacement.
![]()
Human serum promotes robust expansion but suppresses the functionality of NK cells
As the expression of cytotoxic effector molecules and activating and inhibitory receptors did not explain the enhanced functionality of NK cells cultured in AI, we considered the culture medium composition. The RC and DS media contained 10% serum (10% FBS for RC and 10% human serum for DS), whereas the AI medium was serum-free. As cells cultured in serum-free culture conditions (AI and DS-AI) exhibited significantly higher functionality than those cultured in serum-supplemented conditions (RC, DS, and AI-DS), we assumed that serum promotes proliferation but suppresses the killing potency of NK cells. To confirm this, we added 10% human serum to AI (AI+10% human serum) and observed the proliferation rate and functionality of the cells. The fold expansion of NK cells cultured in AI+ 10% human serum was significantly higher than that of NK cells cultured in AI on day 28 (Fig. 4A). Purity was not altered (Fig. 4B) by the addition of serum. NK cells cultured in AI+10% human serum exhibited significantly lower cytotoxicity than those cultured in AI (Fig. 4C–E).
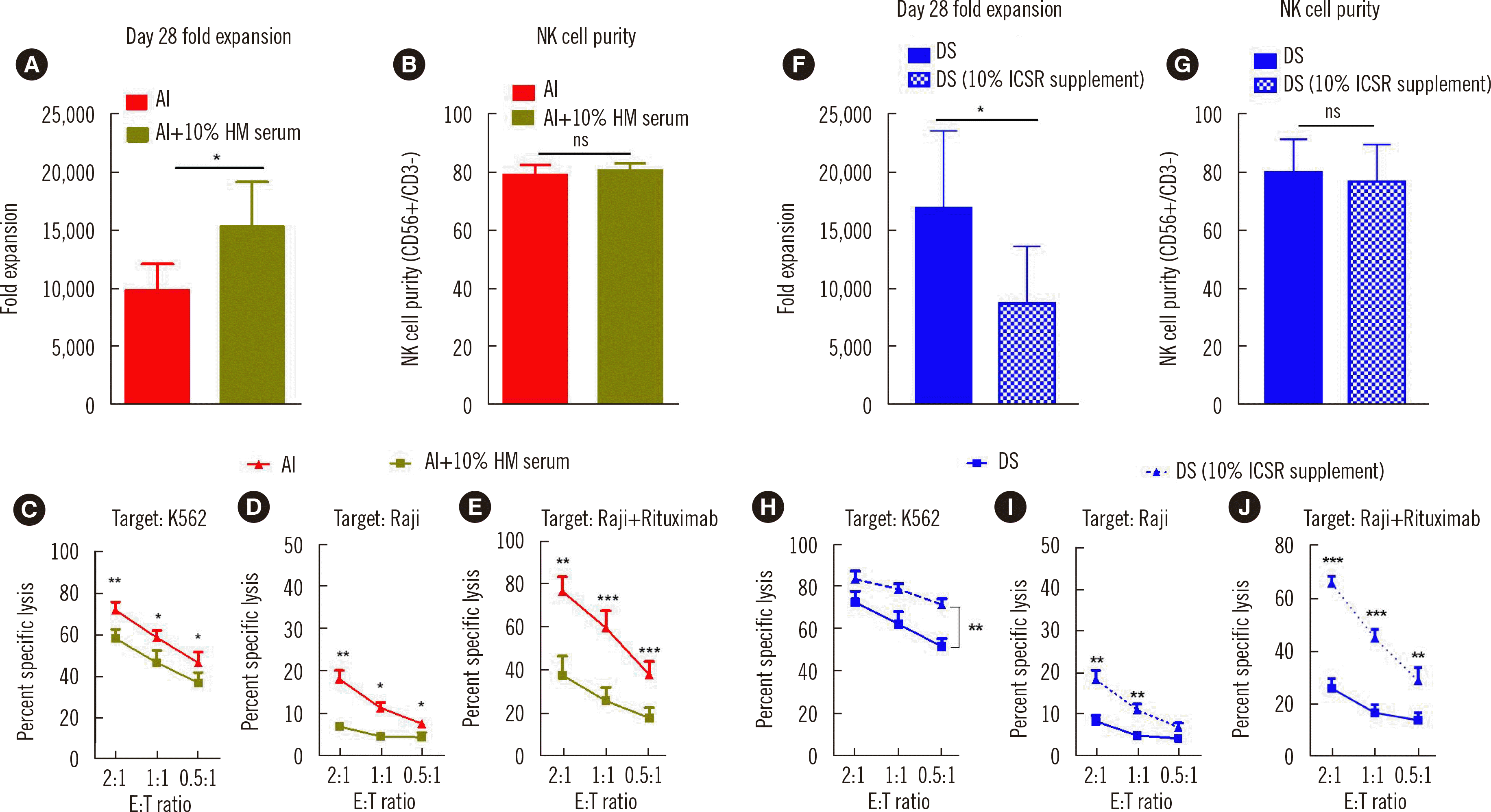
Fig. 4
Human serum in culture media has contrasting effects on NK cell expansion and functionality. (A) Fold expansion of NK cells cultured in AI (red) and AI+10% human serum (dark green) (N=7) on day 28. (B) Purity (CD56+/CD3– NK cells) of UCB NK cells cultured in AI (red) or AI+10% human serum (N=7) on day 28. Flow cytometry-based assay of the cytotoxicity of UCB NK cells cultured in AI + 10% human serum or AI toward (C) K562 cells (N=7), (D) Raji cells (N=4), (E) Raji cells+rituximab (N=5) on day 14. (F & G) Fold expansion (N=5) and purity (N=5) of NK cells cultured in conventional DS medium (10% human serum supplementation) and DS medium in which the 10% human serum was replaced with 10% ICSR. (H–J) Flow cytometry-based cytotoxicity assay of UCB NK cells cultured in the respective media (N=5) on day 14. *P<0.05; **P<0.01; ***P<0.001.
Abbreviations: DS, DMEM+Ham’s F12+various supplements; AI, AIM V+ICSR; NK, natural killer; UCB, umbilical cord blood; ICSR, Immune Cell Serum Replacement.
![]()
To examine the contrasting effects of serum supplementation on NK cell expansion and functionality further, we removed the human serum from DS and added 10% ICSR instead. NK cells cultured in DS + 10% ICSR demonstrated significantly reduced fold expansion (Fig. 4F–G), whereas their functionality was significantly increased compared with that of NK cells cultured in DS (Fig. 4H–J). Lastly, we reduced the human serum level from 20% to 5% in DS medium. In line with the above results, when the human serum level was decreased, the fold expansion of the NK cells decreased, whereas their functionality increased (Supplemental Data Fig. S2A–E).
NK cells cultured in serum-free medium demonstrate rapid target-cell killing
To examine the potential reasons for the enhanced killing potency of NK cells cultured in serum-free medium, we observed the killing behaviors of cells cultured in DS (with serum) and AI (serum-free) using live cell imaging. K562 target cells (green) killed by NK cells (unlabeled) exhibited extensive membrane blebbing and cell shrinkage (Fig. 5A, Supplemental Videos 1A and 1B). The time required for killing, i.e., the time passed between initial NK cell contact and onset of membrane blebbing, was measured for each NK cell that successfully killed target cells [17]. NK cells cultured in AI were more than four times faster in killing than those cultured in DS (Fig. 5B). Thus, the killing capacity of NK cells (the fraction of NK cells that killed more than one target cell during 4 hours of imaging) was significantly higher for cells cultured in AI than for those cultured in DS (Fig. 5C). These results indicate that the superior cytotoxicity of NK cells cultured in serum-free AI is owing to their rapid recognition and killing capabilities. As the live imaging results demonstrated the rapid killing capability of NK cells cultured in AI, we analyzed NK cell cytotoxicity over various periods (30, 90, 150, and 240 minutes) (Fig. 5D). The cytotoxicity of NK cells cultured in AI tended to saturate after 90 minutes, whereas that of NK cells cultured in AI+10% human serum saturated only after 150 minutes. The fold change in cytotoxicity of cells cultured in AI+10% human versus those cultured in AI was the highest at 90 minutes (1.6-fold) and the lowest at 240 minutes (1.1-fold). These results indicate that serum in the culture medium delays the decision time for target cell killing, thus reducing the killing potency of NK cells.
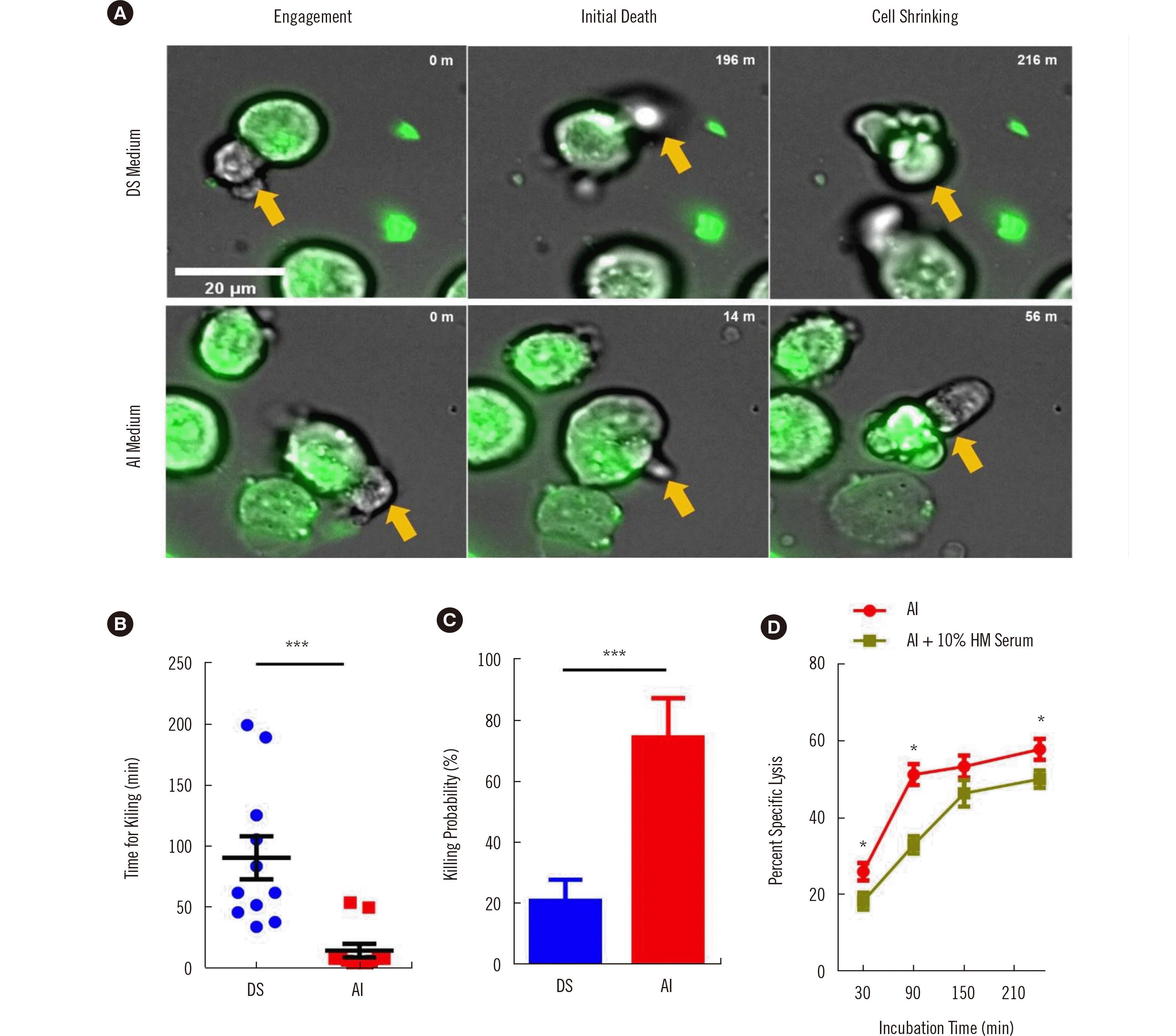
Fig. 5
Killing behaviors of NK cells cultured in different media. (A) Representative time-lapse images of NK cell killing events after cell contact. K562 cells are indicated with green fluorescence and NK cells with yellow arrows. (B) Time for target cell killing (N=11) and (C) killing capacity of NK cells as determined from time-lapse images (N=12). (D) Killing capacity of NK cells cultured in AI or AI + 10% human serum (N=4) at 30, 90, 150, and 240 minutes. *P<0.05; **P<0.01; ***P<0.001.
Abbreviations: DS, DMEM+Ham’s F12+various supplements; AI, AIM V+ICSR; NK, natural killer; UCB, umbilical cord blood; ICSR, Immune Cell Serum Replacement; HM serum, human serum.
![]()
DISCUSSION
For the success of NK cell-based cancer immunotherapy, ex vivo expansion methods capable of producing high numbers of highly cytotoxic NK cells are essential. We demonstrated that culture medium supplementation significantly modulates the expansion and cytotoxicity of UCB NK cells.
Each culture medium selected in our study had unique features. Maximum expansion was achieved in human serum-supplemented DS culture medium, whereas functions were similar with RC cultured NK cells. On the contrary, maximum cytotoxicity was achieved in serum-free AI medium but proliferative capacity was not outstanding as DS. Moseman, et al. [14] previously reported increased direct cytotoxicity functions of NK cells cultured in AI medium. Since ADCC is also an important function of NK cells, we evaluated ADCC functions distinctively. Surprisingly, the ADCC of NK cells cultured in AI was nearly twice that of RC and DS. Previous study conducted by Li, et al. [18] demonstrated competitive binding between serum IgG and rituximab Fc segment with FcγRIIIa. As such a possible explanation of superior ADCC function could be the absence of serum IgG in the composition of serum free AI medium. On the other hand, inferior ADCC function of NK cells cultured in serum supplemented media (DS and RC) might have been affected by the high concentration of serum IgG.
With respect to the receptors expression of NK cells cultured in different culture media, consistent with our previous results and those reported by other groups, we observed that feeder cells and the cytokines IL-2 and IL-15 upregulated the expression of activating receptors (NKG2D, CD16, and NKp30) in all culture conditions [13, 19, 20]. However, the varying receptor expression levels did not explain the superior functionality of NK cells cultured in AI.
Based on the unique features of culture medium, we next thought to combine both advantages by media shifting. Interestingly, media in which NK cells were cultured in the later phase of expansion were found to determine cytotoxicity. Specifically, functions of NK cells that were medium shifted from DS to AI were enhanced showing similar function with AI cultured NK cells. In vice versa culture condition, (AI to DS condition) function of NK cells was reduced demonstrating similar function with DS and RC cultured NK cells (Fig. 3C-D). Although media shifting was not able to surpass the expansion rate of DS, their functions were fully recovered and expansion was significantly enhanced compared to AI. Taken together, appropriately combining unique advantages of a culture media by media shifting can be a great strategy to fully satisfy obtaining high quantity and high quality of CB-derived expanded NK cells.
Although the exact composition of AI culture media is not fully disclosed, one of the main differences in media composition between AI and DS was the presence of human serum. To further characterize the effect of human serum, we slightly modified the composition of each media by adding a human serum to serum-free media (AI) and extracting human serum components from DS and instead supplementing with Immune Cell Serum Replacement (ICSR). As a result, we found that the differential regulation of NK cell expansion and functionality by DS and AI media is due to the human serum component which promoted the expansion of NK cells while inhibiting their functionality by delaying target cell recognition. Medvec, et al. [21] reported that T cells and CAR-T cells cultured in serum-free medium showed enhanced killing potency both in vivo and in vitro. Our results indicate that like T cells, NK cells are affected in functionality by the serum component.
Human serum composition is extremely complex; it is difficult to ascertain exactly which of its components suppress the killing potency of NK cells. However, it is clear that human serum has dual effects in strongly promoting the proliferation of NK cells but suppressing their functionality. As for other types of culture media supplementation, human platelet lysate-supplemented culture media may be an effective alternative for producing high numbers of highly functional NK cells [22]. However, our main objective in this study was to compare and elucidate the effect of human serum on cell expansion and functionality, and therefore, we did not analyze the effect of human platelet lysate. The effect of human platelet lysate on NK cell expansion and functionality is worth further study.
Ruggeri, et al. [23] have shown that native extracellular protein mixtures, such as bovine serum albumin, hemoglobin, and FBS, lead to the non-specific binding of soluble proteins to lipid bilayers. Thus, a possible explanation for the delayed and suppressed target cell killing of NK cells cultured in serum-containing medium are non-specific interactions between the lipid bilayers of NK cells and extracellular soluble proteins, which may arbitrarily and non-specifically bind to activating and inhibiting receptors of the NK cells, delaying signal production for the activation of the downstream killing mechanisms. Another possible explanation for the rapid and enhanced killing capability of AI-cultured NK cells is the difference in the metabolic fitness state of expanded NK cells cultured in serum-free conditions. Because human serum has numerous components, we could not identify the specific components that reduced NK cell cytotoxicity which is one limitation of our study.
In conclusion, our results suggest that culture medium supplementation with serum promotes the expansion of NK cells while inhibiting their functionality by delaying target cell recognition. Thus, selection of culture media should be concerned as a main factor to determine both of NK cell proliferation and function for adoptive cell therapy. Lastly, we suggest a strategy of media shifting during NK cell expansion in obtaining high quantity and high quality of NK cells.

 XML Download
XML Download