

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Pain and its consequences are regarded as a major and complicated health issue. One of the most common types of nociception is orofacial pain, which affects the face, teeth, lips, tongue, and oral regions [1].
The rostral ventromedial medulla (RVM) is one of the most important brainstem regions that exerts both inhibitory as well as facilitating effects on nociceptive relay [2]. Interestingly, it has been indicated that the RVM subareas, including the reticularis gigantocellularis and raphe magnus nuclei [3], have connections with the hippocampus [4].
Orexin-A and B peptides are detected in several areas of the brain. The orexin-1 and -2 receptors are two types of receptors that mediate orexin-A roles [5]. In addition, previous studies have shown that the orexinergic system has been implicated in many behavioral and physiological processes, including nociception modulation, anxiety, learning and memory functions [6–8]. In addition, the presence of orexin fibers and receptors within the regions involved in these functions has been previously documented [9,10].
Cyclooxygenase 2 (COX-2) is an isoform that has been involved in the neuro-inflammatory and nociceptive processes [11]. This inflammatory mediator has been demonstrated to be rapidly elevated in response to noxious and inflammatory stimuli [12]. Rahbar et al. [13] reported that intradental capsaicin application significantly elevates COX-2 levels in the trigeminal subnucleus caudalis.
Brain-derived neurotrophic factor (BDNF) is a molecule that is involved in various neurophysiological functions such as nociception [14,15] and learning and memory [16]. BDNF is believed to be present in various regions of the brain, particularly those related with cognitive processes, such as the hippocampus [17]. BDNF and its receptors are expressed in nociceptive fibers and orofacial nociception-reliant areas such as the trigeminal ganglion and the spinal trigeminal nucleus [14].
By considering the aforementioned point of views, the purpose of this study was to investigate the molecular mechanism by which orexin-A signaling in the RVM and hippocampus affects capsaicin-induced pulpal nociception in male rats.
MATERIALS AND METHODS
1. Animals
Thirty-six male Wistar rats were randomly distributed into six groups (6 animals per group). The rats were kept in typical animal housing, including a temperature of 24°C and a 12-hour light-dark cycle. The experimental methods and procedures were reviewed and confirmed by the Ethics Committee of the Kerman Neuroscience Research Center, Kerman, Iran (code: IR.KMU.REC. 11-96). At the end of the tests, the rats were anesthetized and the hippocampus was extracted, frozen in liquid nitrogen, and stored at –80°C until processing. Then, immunofluorescence detection was performed on slices of RVM samples.
2. Animal groups
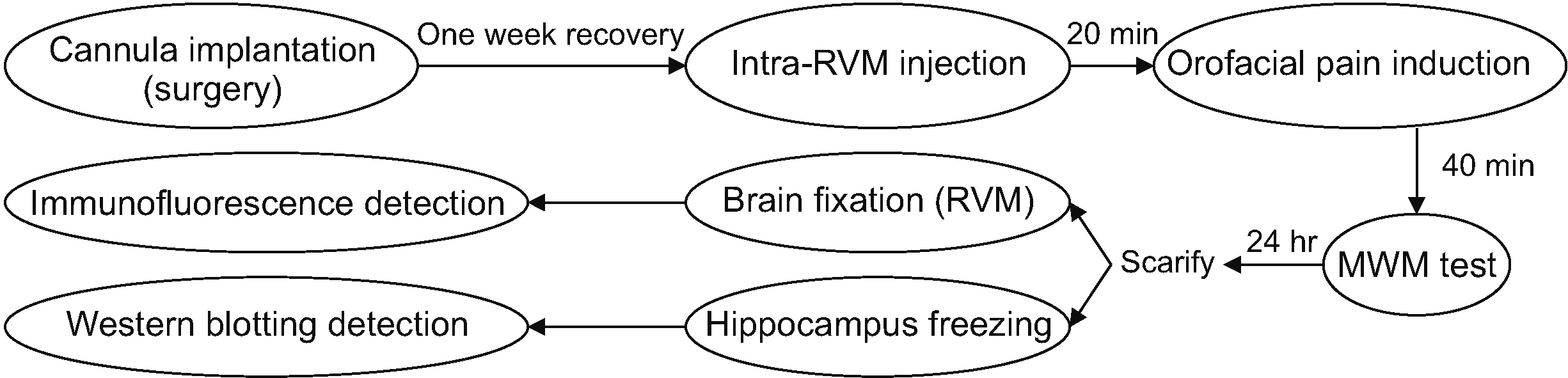
Six groups of animals were designated (n = 6): control rats, capsaicin rats, a capsaicin vehicle group (sham), a CSF + Caps group, an ORX 50 pmol/1 μL/rat + Caps group, and an SB 80 nmol/1 μL/rat + ORX 50 pmol/1 μL/rat + Caps group. The number of observations for western blotting and immunofluorescence experiments was n = 4. Drugs were microinjected 20 minutes before capsaicin application. The experimental procedure is presented in Fig. 1.
3. Surgical method
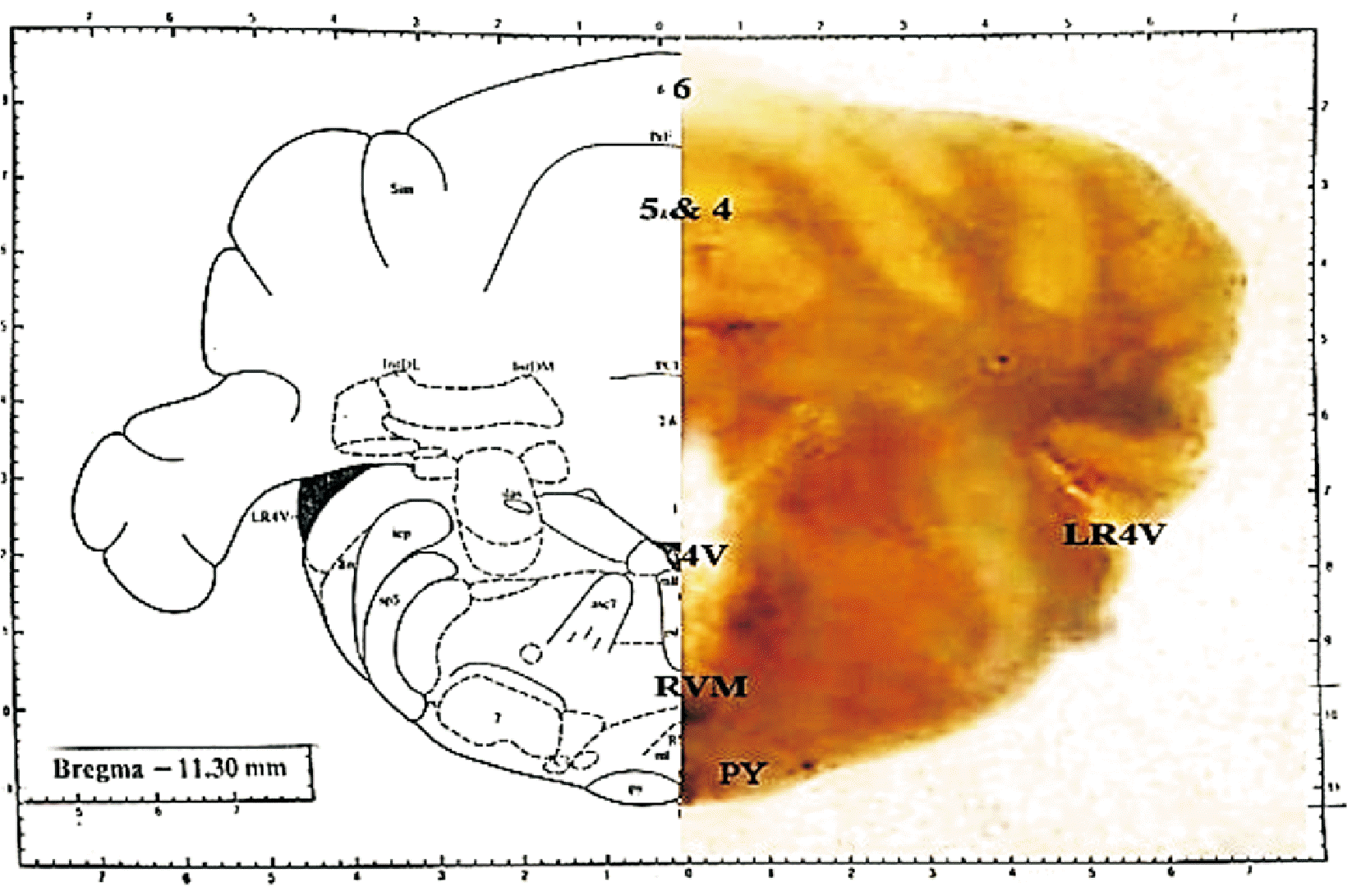
Intraperitoneal (i.p.) infusions of ketamine (65 mg/kg) and xylazine (10 mg/kg) were used to anesthetize the rats. The animals were mounted onto a stereotaxic device (Stoelting Co., Wood Dale, IL) and the guiding cannula point was targeted toward the RVM area (posterior to the bregma: 11.3 mm, lateral from the midline: 0 mm, and deep to the cortical surface above the RVM: 10.3 mm) (Fig. 2).
4. Drugs and microinjections
Orexin-A (Tocris Bioscience, Bristol, UK) was added to synthetic CSF. The orexin-1 receptor (OX1R) antagonist SB-334867-A (Tocris Bioscience) was dissolved in dimethyl sulfoxide and diluted in synthetic CSF. The drugs were injected into the RVM with a polyethylene syringe (volume: 1 μL). Capsaicin (Sigma-Aldrich, St. Louis, MO) was added to a solution containing 80% distilled water, 10% ethanol, and 10% Tween 80. Capsaicin (100 μg/10 μL) was intradentally administrated in the rats’ mandibular incisors.
5. Orofacial-nociceptive induction
The following steps were performed for the induction of nociception:
Short-term anesthesia was induced with CO2.
A small cavity was created (1.5 × 1.5 × 1.5 mm) on the one-third distal side of the right mandibular incisor using a dental fissure bur (D&Z OHG Company, Wisbaden, Germany).
A tiny cotton pellet damped with capsaicin mixture was placed on the constructed cavity under a light-cured glass-ionomer (Fuji II; GC Corporation, Tokyo, Japan) to provide noxious stimulation.
The cumulative time spent on nociceptive behaviors was measured as total nociceptive behavior time [18].
6. Spatial learning and memory test
The Morris water maze (MWM) task was used to evaluate the learning parameters. The water maze consists of a circular tank (black in color, depth 35 centimeters, water temperature 21°C ± 1°C) that is subdivided into four quadrants of similar size. In the target quadrant, a platform (diameter: 10 cm) was hidden 2 cm under the water’s surface. To facilitate spatial navigation, prominent visual cues were placed on the room’s walls. The digital tracking system (Noldus Information Technology, Wageningen, The Netherlands) was designed to monitor and record animal activity automatically. The escape latency and the distance travelled parameters were examined.
7. Western blotting test
The hippocampal tissues were homogenized in radioimmunoprecipitation lysis buffer, which also included protease inhibitors. The homogenized tissues were centrifuged at 14,000 rpm for 15 minutes at 4°C. The supernatant was then centrifuged twice or three times. Before protein samples were transferred to polyvinylidene difluoride membranes, 40 µg of each protein sample was separated electrophoretically on sodium dodecyl sulfate-polyacrylamide gel. In the next step, the membranes were blocked (2 hours) with 5% nonfat dried milk in Tris-buffered saline with Tween 20 (TBS-T), and then incubated (overnight, at 4°C) with primary antibodies (COX-2 and BDNF) (1:1,000). After three washes in TBS-T, the membranes were incubated for 60 minutes with a secondary antibody (1:15,000) attached to horseradish peroxidase. The antibody-antigen complexes were recognized using an ECL-Advance system and Lumi-Film chemiluminescent detection film (Roche, Mannheim, Germany). Lab Work analysis software (UVP, Cambridge, UK) was used to examine the band data. As a loading control, β-actin (1:10,000) was used. The results were reported as a protein/β-actin ratio for each sample.
8. Immunofluorescence detection
The animals were anesthetized with a ketamine/xylazine combination (i.p. injection) twenty-four hours after the induction of nociception, and the brain tissues were fixed with a 0.9 percent saline solution (100 mL) followed by a 4% paraformaldehyde solution via cardiac perfusion. After that, the brains were post-fixed and incubated overnight in the same fixative. The fixed tissues were dehydrated in ethanol solutions and then cleaned in xylene before being embedded in paraffin. A microtome was used to make thick tissue slices (4–6 microns) from the RVM area in accordance with the rat brain map (Paxinos and Watson). The slides were dewaxed in xylene and rehydrated in graded alcohols prior to immunostaining. Antigen retrieval was done in citrate buffer (pH 6.0) because formalin fixation and paraffin embedding make it hard to see specific antigenic determinants.
The slices were then treated for 5–10 minutes in 0.1%–1% hydrogen peroxide. In the next step, the slides were immersed in 1.5% normal blocking serum for 1 hour and washed again with PBS (twice for 5 minutes). Afterward, specimens were incubated (overnight at 4°C) with 1:500 dilutions of primary antibodies for COX-2 and BDNF dilutions (Santa Cruz Inc., Dallas, TX). The slides were then rinsed in PBS (twice for 5 minutes) and incubated with fluorochrome-conjugated secondary antibody (1:1,000) (Santa Cruz Inc.) in a dark chamber for 90 minutes.
The slices were then rinsed in PBS after being treated with propidium iodide. Finally, the samples were mounted in glycerol/PBS and viewed at 20 and 40× magnification using an Olympus fluorescent microscope (Olympus, Tokyo, Japan).
9. Statistical analysis
The statistical significance of nociception behavior, learning and memory, immunofluorescence, and western blotting tests between animal groups was determined using a one-way ANOVA, followed by Tukey’s post-hoc test. A significance level of P < 0.05 was considered statistically significant.
RESULTS
1. Orexin-A and capsaicin-induced nociceptive behaviors
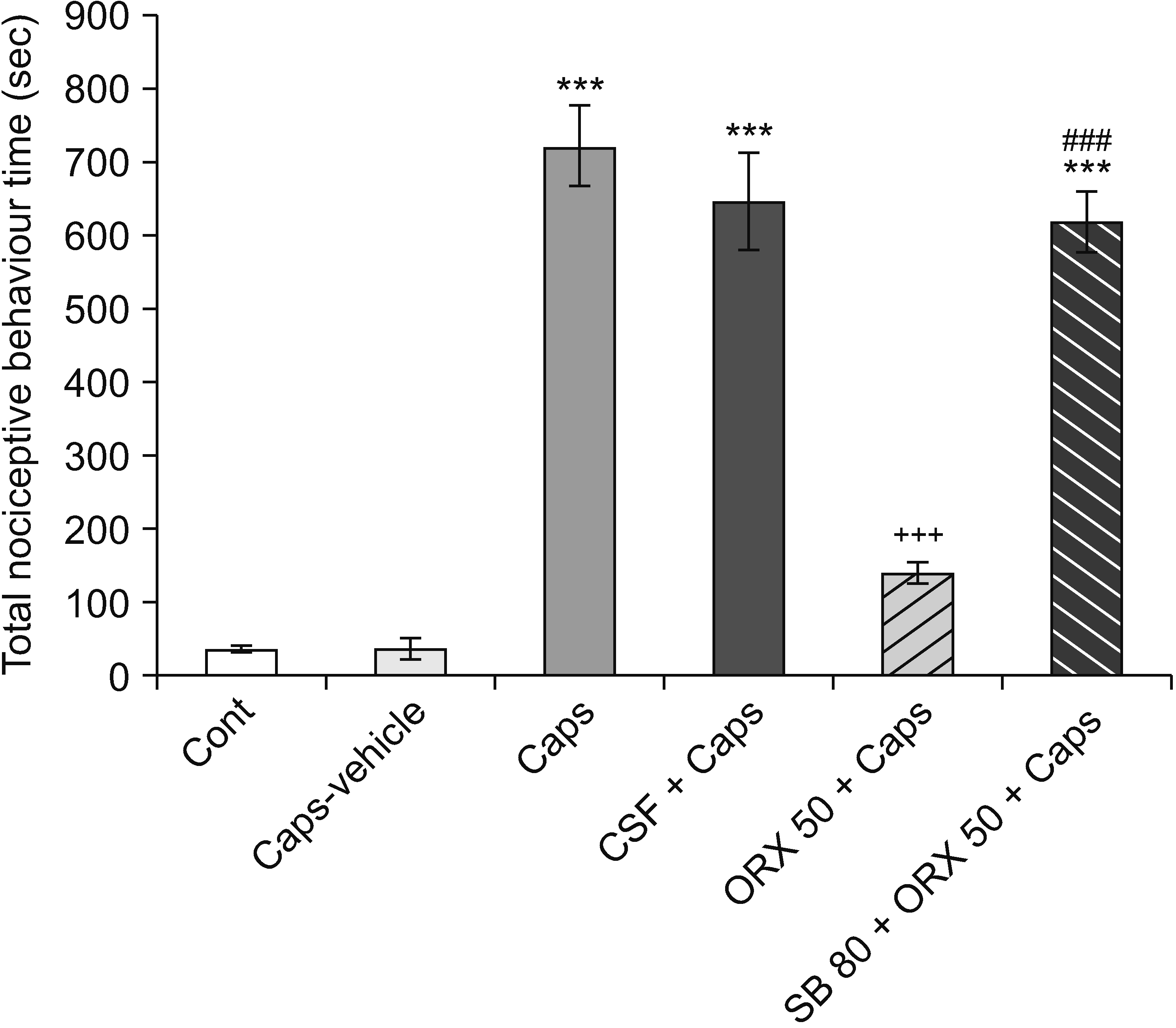
The data showed that during a 40 minutes test procedure, intradental capsaicin significantly increased the total time of nociceptive behaviors in comparison with the control and capsaicin vehicle (sham) rats, indicating that capsaicin has a nociceptive effect on animals (P < 0.001). Administration of orexin-A into the RVM attenuated capsaicin-induced pulpal nociception. Additionally, orexin-A-induced anti-nociception was prevented by pretreatment with SB-334867-A (P < 0.001) (Fig. 3).
2. Orexin-A and spatial learning and memory parameters in capsaicin-treated rats
Our data showed that the sham operation group showed no significant difference from the control group. In addition, the data from the CSF + capsaicin group was the same as that of the capsaicin-treated rats. So, to avoid group duplication, the data for these groups is not shown in the following figures.
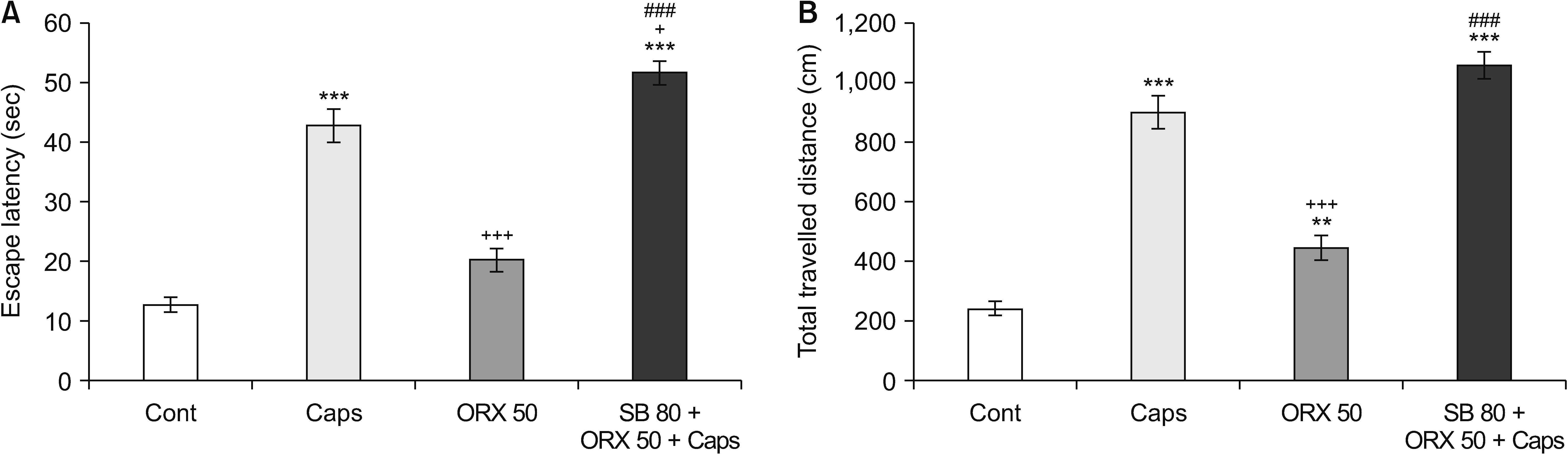
In the MWM test, the results indicate that orofacial pain-like behavior induction by capsaicin significantly increased escape latency and total travelled distance in comparison with the control group (P < 0.001). Moreover, orexin-A (50 pmol/1 μL/rat) injection into the RVM decreased traveled distance as well as latency time in the MWM task, which shows orexin-A has an improving effect on learning and memory processes. Furthermore, the OX1R antagonist (SB-334867-A) completely prevented the precognitive effects of orexin-A in the MWM test (P < 0.001) (Fig. 4).
3. Western blotting analysis
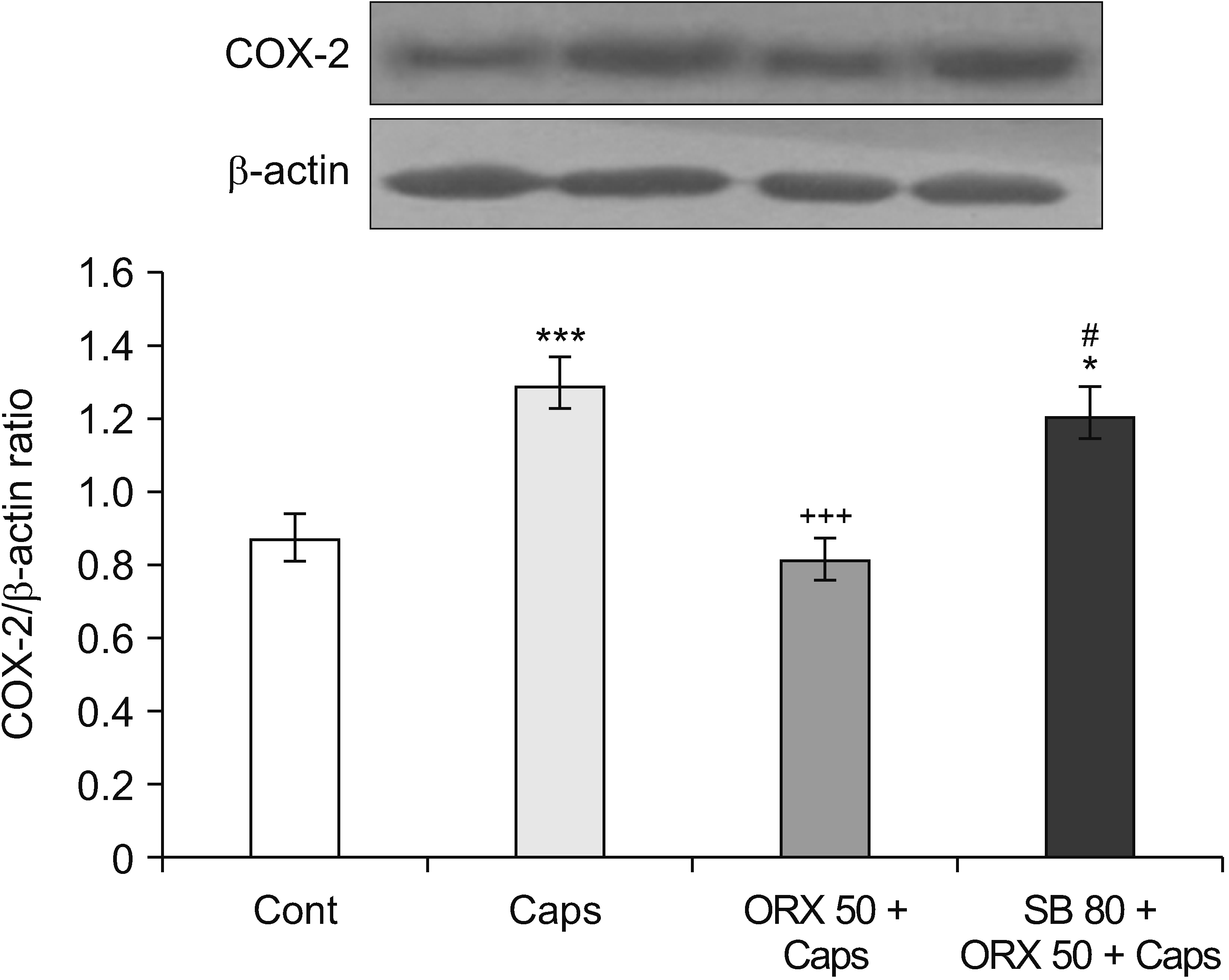
1) The effects of orexin-A on COX-2 levels in the hippocampus after capsaicin treatment
Western blot analysis revealed a significant increase in hippocampal COX-2 levels 24 hours after capsaicin treatment as compared to control rats (P = 0.004). When compared to the capsaicin group, orexin-A treatment decreased nociception-induced COX-2 overexpression (P < 0.001). Additionally, the amount of COX-2 expression in rats treated with SB-334867-A was remarkably similar to that in rats treated with capsaicin (Fig. 5).
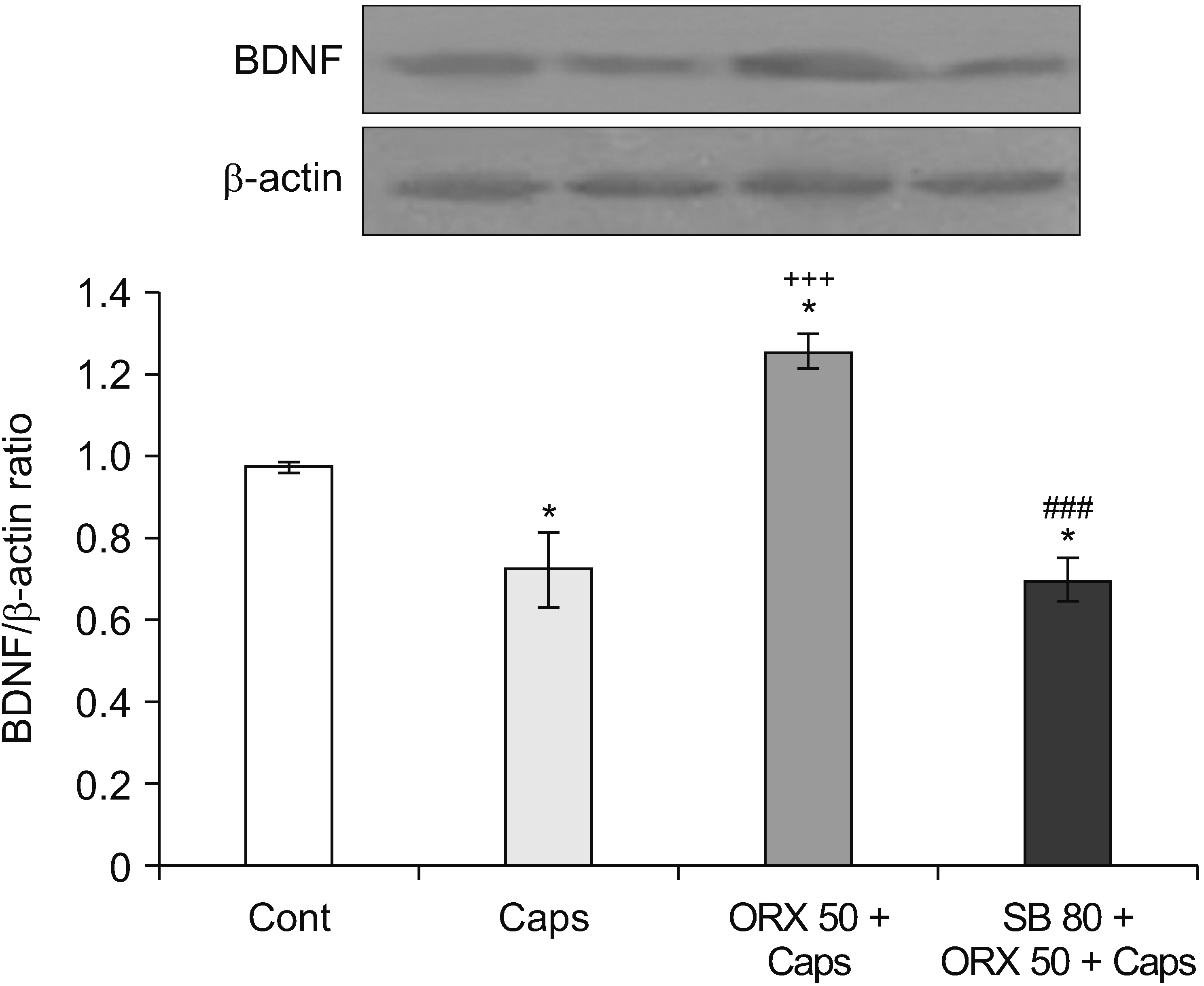
2) The effects of orexin-A on BDNF levels in the hippocampus after capsaicin treatment
Fig. 6 demonstrates that BDNF expression levels in the hippocampus of rats treated with capsaicin were considerably lower than in untreated control rats (P = 0.037). Moreover, BDNF expression was significantly increased in rats that received orexin-A (50 pmol/1 μL/rat). The results also show that SB (80 nmol/1 μL/rat) administration reversed this effect and BDNF expression was significantly decreased compared to orexin-A injected rats (P < 0.001) (Fig. 6).
4. Immunofluorescence staining
1) The effects of orexin-A on COX-2 levels in the RVM after capsaicin treatment
Immunofluorescence labeling revealed a significant increase in the number of COX-2-expressing cells in the RVM of capsaicin-treated rats compared to control rats (P < 0.001). However, orexin-A treatment inhibited capsaicin-induced COX-2 upregulation (P < 0.001). In addition, the central injection of SB-334867, a selective OX1R antagonist, reduced the inhibitory effects of orexin-A and increased the number of positive COX-2 cells as compared with control and orexin-A injected animals (P = 0.018, P = 0.007) (Fig. 7).
2) The effects of orexin-A on BDNF levels in the RVM after capsaicin treatment
The results of immunofluorescence staining in Fig. 8 indicated that the numbers of BDNF-expressing cells in the capsaicin-treated animals were significantly lower than in the control group (P = 0.002). Orexin-A administration significantly increased the number of BDNF immuno-reactive cells in comparison with the capsaicin-treated rats (P = 0.023). Furthermore, in SB-334867-A-treated rats, the number of positive BDNF cells decreased as compared with the control group (P = 0.008).
DISCUSSION
The present study showed that intradental injection of capsaicin increases COX-2 expression in rats’ RVM and hippocampus. Additionally, intra-RVM administrations of orexin-A significantly inhibited capsaicin’s effects. SB-334867, an OX1R antagonist, however, significantly inhibited the effects of orexin-A.
COX-2 proteins have been implicated in a variety of inflammatory nociception conditions [19,20]. COX-2 has also been found to play a role in orofacial nociception models [21]. Ahn et al. [22] reported that the suppression of COX-2 central pathways has anti-nociceptive effects on temporomandibular joint inflammatory conditions in rats.
Capsaicin induces an inflammatory nociception sensation, which promotes elevated COX-2 levels in the caudalis nucleus and trigeminal ganglion cells [23,24]. In addition, COX-2 protein can be overexpressed at the spinal level following carrageenan administration [25].
Recent studies have shown that the RVM is the main site of nociceptive modulation and it presents a heterogeneous area that includes three types of neurons or cells: on-cell (nociception-stimulating neurons), off-cell (nociception-inhibiting neurons), and neutral cells (don’t change in nociception) [26]. Capsaicin administration in rats leads to pain-related behaviors, which are correlated with the enhancement of RVM on-cells activity [27]. Based on these observations, RVM cells, especially on-cells, may be sensitized by capsaicin and, thus, can contribute to increased COX-2 expression in the RVM and hippocampus in orofacial nociception conditions.
Furthermore, the results showed that orexin-A microinjection into the RVM significantly reduced capsaicin-induced COX-2 up-regulation in the RVM and hippocampus. These findings support orexin’s anti-inflammatory effect in a rat model of orofacial nociception [28]. Numerous studies have reported that orexin-A microinjection into different brain areas produces an anti-nociceptive effect. Furthermore, administration of OX1R antagonists, including SB-334867, leads to a decrease in orexin-A-induced anti-nociceptive results [29].
The endogenous orexin system improves the survival of mice with decreased levels of multiple chemokines such as interleukin-6 and tumor necrosis factor-alpha [30] or by inhibiting some signaling pathways [31]. Becquet et al. [32] reported that orexin-A administration decreased transforming growth factor beta expression levels. This cytokine has been associated with anti-inflammatory responses in some instances. As a result, orexin-A may play a positive function in inflammation by inhibiting the development of pro-inflammatory molecules such as COX-2.
Our results also revealed that capsaicin-induced orofacial nociception reduced the level of BDNF expression, whereas intra-RVM orexin-A administration showed a reverse effect on BDNF expression levels in the RVM and hippocampus. BDNF has been implicated in a variety of physiological activities, including nociceptive neurotransmission [15]. Furthermore, it has been demonstrated that BDNF plays a critical role in hippocampal-dependent learning and memory function [33]. BDNF mRNA expression has also been demonstrated in areas involved in orofacial nociception modulation, such as the trigeminal nucleus caudalis (Vc) and the trigeminal ganglion [34].
Numerous investigations have established the significance of BDNF in inflammatory and neuropathic nociception [35]. BDNF appears to influence both peripheral and spinal sensitivity to chronic nociception [36]. Duric and McCarson [37] observed that BDNF levels were decreased in the hippocampus during chronic inflammatory nociception. Furthermore, a nociception stimulator such as capsaicin may induce neural damage and toxic effects, which probably cause a reduction in BDNF expression through the increased level of intracellular reactive oxygen species [38,39].
The findings show that the RVM microinjection of orexin-A led to an increase in the expression of BDNF in rats receiving capsaicin. Kooshki et al. [21] reported that orexin-A injection into the Vc increases BDNF in the trigeminal subnucleus caudalis.
High levels of BDNF in the hippocampus nucleus have been shown to improve learning and memory function [16]. In addition, the orexigenic system has a role in memory consolidation. Infusion of orexin-A into the hippocampus enhances memory in a MWM task [40]. These results suggest a positive association between orexin-A-induced high levels of hippocampal BDNF and learning and memory, and it can be speculated that orexin-A has a modulating effect on spatial learning and memory through the increased level of BDNF in the hippocampus. It has been suggested that BDNF interacts with different intracellular signaling pathways, such as extracellular signal-regulated kinase (ERK) signaling [41]. Moreover, ERK has a protective effect on hippocampal neurodegeneration [42]. The research shows that BDNF plays a crucial role in neuroprotection by increasing the transcription of survival-related genes [43].
As a result, the authors assume that orexin A-induced BDNF activation may protect against dental pulp nociception and learning deficits. Orexin’s effects on these processes may be mediated by stimulation of the BDNF system in the RVM and hippocampus, as well as through its connections with other signaling pathways. Based on these findings, it is reasonable to speculate that orexin-A is involved in modulating BDNF signaling following the induction of orofacial nociception. It may modulate synaptic neuropeptide release and the release of substance P [44].
Orexinergic fibers and orexin receptors have been identified in multiple brain regions, including the hippocampus and RVM [29,45]. Additionally, COX-2 and BDNF have been shown to be predominantly expressed in the cell bodies and dendritic regions of hippocampal neurons in the central nervous system [46,47]. Interestingly, the hippocampus and RVM share an anatomical relationship [48]. On the other hand, orexin has an effect on COX-2 and BDNF signaling in the hippocampus [49]. Hence, these findings suggest that orexin-A administered into the RVM may be a critical mediator affecting the hippocampal expression of COX-2 and BDNF. However, additional investigations into the involvement of RVM orexin-A and BDNF/COX-2 signaling pathways in orofacial nociception and learning and memory processes are still needed.
In conclusion, the data from this study indicate that orexin-A appears to exert a significant modulatory influence on dental pulp nociception, as well as on pain-induced learning and memory impairments. Modulation of COX-2 and BDNF expression levels in the RVM and hippocampus may be the mechanism of action. However, additional research is necessary to ascertain the precise mechanism(s) behind these relationships.


 XML Download
XML Download