

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Neutrophils play a pivotal role in bacterial infections as a first line of defense against invading microbes [1,2], However, key neutrophil functions regarding microbial clearance (chemotaxis, phagocytosis, respiratory burst, and intracellular killing of bacteria) are frequently compromised in severe bacterial infections [3-10]. Further, among the patients with bacterial infections, those exhibiting reduced intracellular killing of bacteria by neutrophils display higher infection-related mortality [4]. Meanwhile, impaired intracellular killing of Staphylococcus aureus (S. aureus) by neutrophils precedes nosocomial infections in critically ill patients [11]. In all of these circumstances where intracellular killing of bacteria by neutrophils is depressed, substances that enhance neutrophil functions could be helpful as an adjunctive immunomodulatory therapy [12].
Previously we reported that lysophosphatidylcholine (LPC), an endogenous lipid, enhances neutrophil bactericidal activity via the glycine (Gly)/glycine receptor (GlyR) α2/TRPM2/p38 mitogen-activated protein kinase (MAPK) pathway [13]. LPC via G2A (from G2 accumulation, also known as the G protein-coupled receptor GPCR132) induces the translocation of intracellularly located GlyRα2 to the plasma membrane [14]. Extracellular glycine is necessary for LPC-induced enhancement of neutrophil bactericidal activity and LPC-induced p38 MAPK activation [13].
However, the effect of glycine per se (in the absence of LPC) on neutrophil bactericidal activity is unknown. In this study, we show that glycine enhances neutrophil bactericidal activity by enhancing azurophil granule-phagosome fusion via the GlyRα2/reactive oxygen species (ROS)/calcium/p38 MAPK pathway, proposing glycine as a useful antimicrobial agent that enhances neutrophil bactericidal activity.
Go to : 
METHODS
Preparation of neutrophils
Venous blood was taken from healthy volunteers in accordance with a protocol approved by Ethnic committee of Hallym University. Neutrophils were purified from heparinized venous blood from healthy volunteer donors. In brief, neutrophils were isolated by density centrifugation in Histopaque-1077 followed by dextran sedimentation. Residual erythrocytes were eliminated with hypotonic lysis. The purity of neutrophils counted by Diff Quik staining was > 95% on average. The viability of neutrophils stained with trypan blue was > 99%. After the isolation procedure, neutrophils were dispersed in RPMI 1640 supplemented with 5% fetal bovine serum (FBS).
Assay of intracellular killing of E. coli by human neutrophils
Bactericidal activity of human neutrophils was measured by a slightly modified version of a previously described method [13,14]. In brief, neutrophils (4 × 106/ml, 0.5 ml) were seeded on a 13-mm plastic coverslip (coated overnight with 0.05% poly-L-lysine) in a 24-well plate for 1 h in a humidified CO2 incubator (5% CO2 and 95% air atmosphere). Coverslips were then transferred to a 60-mm dish (6 coverslips/dish) containing 2 × 107 E. coli (overnight-cultured E. coli [DH5-α] was opsonized with 10% human serum for 30 min) at a neutrophil/E. coli ratio of 1:10 for 20 min for E. coli uptake. After the 20 min uptake, 18.3 ± 1.9% (mean ± standard error [SE], n = 2) of E. coli was removed by adherent neutrophils. Coverslips were then transferred to a 24-well plate with 500 μl of RPMI-1640 (supplemented with 5% FBS) containing vehicle or glycine (133, 300, 500, and 1,000 μM). After 15 min, the coverslips were transferred to a 24-well plate with ice-cold distilled water for 1 h for lysis of neutrophils (colony forming units [CFUs] after 15 min of incubation). To evaluate the total number of phagocytosed E. coli, some coverslips were directly transferred to ice-cold distilled water after 20 min of E. coli uptake without an additional 15 min of incubation with glycine (CFUs before 15 min of incubation). E. coli was plated in lysogeny broth agar plate and cultured overnight at 37°C, after which viable colonies were counted. The percentage of bacteria killed was calculated as 100× (1-CFUs after 15 min of incubation/CFUs before 15 min of incubation). To determine the effects of various inhibitors, assay of bactericidal activity was performed in the presence of strychnine (1 μM), diphenylene iodonium (DPI) (1 μM), protease inhibitor mixture (leupeptin [10 μg/ml], N-tosyl-L-leucin chloromethyl ketone [10 μg/ml], pepstatin A [10 μg/ml], aprotinin [10 μg/ml], and phenylmethylsulfonyl fluoride [1 mM]) or SB203580 (10 μM). All media were supplemented with 5% FBS.
Clearance of intraperitoneally injected E. coli
Male ICR mice at the age of 6 weeks (25–28 g body weight) were used. Procedures for the animal experiments were approved by the Animal Experimentation Committee at Hallym University. Mice were intraperitoneally injected with E. coli (5 × 108). Mice were pretreated subcutaneously with PBS or strychnine (0.4 mg/kg) for 30 min before E. coli injection. At 30 min after E. coli injection, vehicle (PBS) or glycine (30 mg/kg) was subcutaneously injected. Eight hours after E. coli injection, mice were sacrificed, and peritoneal lavage fluid was collected and cultured overnight on blood agar base plates (Trypticase Soy Agar Deeps; BD, Franklin Lakes, NJ, USA) at 37°C and the numbers of CFUs were counted.
Measurement of 36CI– Uptake
Measurement of 36CI– uptake was performed in accordance with a previously described method [15]. Briefly, 6 × 106/ml neutrophils were incubated in HEPES buffer at 37°C for 1 h (in mM: 20 HEPES, 118 NaCl, 4.7 KCl, 1.2 MgSO4 and 2.5 CaCl2, pH 7.4). Neutrophils pretreated with phorbol myristate acetate (PMA) (2 μg/ml) for 15 min were exposed to 2 μCi/ml 36CI– in the presence or absence of glycine (1 mM) for 10 s and transferred to a Wheaton 25 mm vacuum filter system. 36CI– uptake was terminated by washing twice with ice-cold HEPES buffer through 25 mm glass microfiber filters (Whatman GF/A; Cytiva, Marlborough, MA, USA). Glass fiber filters were placed in scintillation vials, and proteins were solubilized with 1.6 ml of 1 N NaOH for 1 h. EcoLume (8 ml) was added and radioactivity was measured using a liquid scintillation detector (LS-3801; Beckman, Fullerton, CA, USA). In some experiments, neutrophils were pretreated for 30–60 min with strychnine (1 μM) before exposure to 2 μCi/ml 36CI– in the presence of glycine (1 mM) for 10 s.
Nitrogen cavitation
Purified neutrophils were resuspended in ice-cold relaxation buffer (minus EGTA, 100 mM KCl, 3 mM NaCl, 1 mM ATP[N]2, 3.5 mM MgCl2, 10 mM piperazine N, N’-bis(2-ethansulfonic acid), pH 7.3) and pressurized with N2 for 20 min at 350 psi with constant stirring in a nitrogen bomb (Parr Instrument Company, Moline, IL, USA) at 4°C. Cavitates were admixed with EGTA, pH 7.4, sufficient for a final concentration of 1.25 mM and centrifuged for 15 min at 500 g to remove nuclei and unbroken cells. Samples were kept on ice until fractionation.
Subcellular fractionation
Nuclei and unbroken cells were pelleted, and the supernatant was obtained by centrifugation of the cavitate at 500 g for 15 min at 4°C. A density gradient of 9 ml of Percoll was prepared (1.120 g/ml, 1.090 g/ml, and 1.050 g/ml). The supernatant, cavitated neutrophils, was loaded onto three separate Percoll density gradients and centrifuged at 48,000 g for 20 min in a P28S rotor in a Himac CP80MX centrifuge (Hitachi, Tokyo, Japan). After Percoll density gradient centrifugation, four distinct bands were observed from the bottom designated the α-band (azurophil granules), the β1-band (specific granules), the β2-band (gelatinase granules), and the γ-band (secretory vesicles and plasma membranes). Each fraction was collected and ultracentrifuged at 35,000 rpm for 120 min to remove the Percoll. After centrifugation, the granule layer was visible as a disk form above the Percoll pellet.
Flow cytometry
To analyze the surface expression of GlyRα2, neutrophils (3 × 106/ml) were stimulated with E. coli for 5 min in RPMI-1640. Then, the neutrophils were fixed with Cytofix (BD Pharmingen) for 5 min. After washing with Pharmingen Stain buffer (bovine serum albumin, BSA) (BD Pharmingen), the neutrophils were stained with Alexa Fluor 488 carboxylic acid succinimidyl ester-conjugated anti-GlyRα2 for 30 min at room temperature. Anti-GlyRα2 antibody was conjugated with Alexa Fluor 488 carboxylic acid succinimidyl ester, in accordance with the manufacturer’s protocol. Neutrophils were washed in Pharmingen Stain buffer (BSA) (BD Pharmingen) and acquired on Guava Easycyte (GE Healthcare, Chicago, IL, USA). Analysis was performed using FCS Express V3 (De Novo Software, Los Angeles, CA, USA). Alexa 488-conjugated mouse IgG1 (BD Pharmingen) was used as an isotype control. To analyze the surface expression of flavocytochrome b558, neutrophils (3 × 106/ml) were stimulated with E. coli at a ratio of 1:10 for 20 min in RPMI-1640. Then, the neutrophils were washed with Pharmingen Stain buffer (BSA) (BD Pharmingen) and stained with PE-labeled anti-flavocytochrome b558 (MBL International, Woburn, MA, USA) for 1 h, and acquired on Guava Easycyte (GE Healthcare). Analysis was performed using FCS Express V3 (De Novo Software).
ROS measurement
Intracellular H2O2 levels were measured with a fluorescent probe, 2´, 7´-dichlorodihydrofluorescein diacetate (DCF-DA), as described previously [16]. Neutrophils were allowed to ingest E. coli for 20 min. After washing out unengulfed E. coli, neutrophils were exposed to vehicle or glycine for the next 15 min in the presence or absence of DPI, further incubated with DCF-DA (10 μM) for 10 min, washed twice with PBS, and then the fluorescence was measured.
Fluorescence measurement of Fluo-3
[Ca2+]i was measured using the fluorescent Ca2+ indicator Fluo-3. Neutrophils were loaded with Fluo-3 AM (5 μM) in HEPES physiological salt solution (HEPES-PSS) (in mM) (NaCl 140, KCl 5, MgCl2 1, CaCl2 1, glucose 10, HEPES 10) for 30 min at 37°C. After washing with HEPES-PSS, Fluo-3 AM-loaded neutrophils were resuspended in HEPES-PSS supplemented with 5% FBS and 133 μM glycine and plated on 96-well plates at a cell density of 3 × 106/ml. Traces of intracellular calcium in Fluo-3 AM-loaded neutrophils were measured at 490 nm/526 nm using a Spectramax M2/e fluorescence microplate reader (Molecular Devices, San Jose, CA, USA). Fluorescent emission readings were recorded every 10 sec. Raw fluorescence was subtracted with average fluorescence during the first 5 min. E. coli at a ratio of 1:10 was added for 20 min. Then, 1 mM glycine was added.
Transfection with microRNA-adapted shRNA of TRPM2
shRNAmir targeting TRPM2 was purchased from Open Biosystems (Huntsville, AL, USA). The sequences were as follows: shRNAmir against TRPM2; 5´-TGC-TGT-TGA-CAG-TGA-GCGACC-TGC-TAT-CCT-GGG-AGA-TCT-ATA-GTG-AAG-CCA-CAG-ATG-TATAGA-TCT-CCC-AGG-ATA-GCA-GGG-TGC-CTA-CTG-CCT-CGG-A-3´. Neutrophils were transfected with short hairpin RNA using Amaxa Nucleofector Technology (Amaxa Biosystems, Cologne, Germany). Transfection was performed following the manufacturer’s instructions. In brief, 1 × 107 neutrophils were resuspended in human monocyte nucleofector solution (100 μl) (human monocyte transfector kit, cat#VPA-1007; Amaxa Biosystems) at room temperature, followed by the addition of 3 μg of shRNA against TRPM2 or control shRNAmir. Transfection was performed in Amaxa Nucleofector II using program Y-001. Immediately thereafter, neutrophils were diluted in 2.5 ml of human nucleofector medium and incubated for 24 h in a humidified CO2 incubator.
Measurement of Lucifer yellow uptake
Lucifer yellow uptake into neutrophils was measured as described previously [17]. Neutrophils (4 × 106/ml) were suspended in Hank’s balanced salt solution (HBSS) supplemented with 5% FBS and allowed to ingest E. coli at a ratio of 1:10 for 20 min. After washing unengulfed E. coli, neutrophils were further incubated with glycine or vehicle for 15 min in the presence of Lucifer yellow (0.5 mg/ml). Then, neutrophils were washed two times with cold PBS and lysed with ice-cold double distilled water (DDW) for 1 h. The Lucifer yellow uptake was measured with a Spectramax M2/e fluorescence microplate reader (Molecular Devices) at 428 nm excitation and 536 nm emission.
Fusion of azurophil granules and zymosan
Texas red-labeled zymosan particles (Z-2843; Molecular Probes, Eugene, OR, USA) were incubated in 10% human serum for 20 min at 37°C, washed twice with PBS, pH 7.4, and resuspended in PBS. Neutrophils (4 × 106/ml) adhering on glass coverslips were incubated at 37°C for 1 h in a 24-well culture plate. Coverslips were then transferred to a 60-mm dish containing Texas Red-labeled zymosan at a ratio of 1:10. After 20 min, neutrophils were washed to remove most of the extracellular Texas Red-labeled zymosan. Neutrophils were transferred to a 24-well plate and 500 μM glycine was added. After 15 min, experimental samples were washed twice with PBS and fixed using 2% paraformaldehyde for 15 min at room temperature. The cells were then permeabilized using 0.05% Triton X-100 for 10 min at room temperature. After 1 h of blocking in PBS containing 3% BSA, the neutrophils were incubated for 1 h with an anti-CD63 antibody (556019; BD Biosciences, Franklin Lakes, NJ, USA), which was used at a 1:50 dilution in blocking buffer. After 1 h of incubation at room temperature, cells were washed three times in 1 × PBS and incubated for 1 h with Alexa Fluor 488 anti-mouse secondary antibody, used at a final dilution of 1:500. After three washes in 1 × PBS, the cells were mounted using mounting medium. Azurophil granule-phagosome fusion was calculated by measuring the area of co-localization between CD63 (an azurophil granule marker) and zymosan particles, which was divided by the total intracellular zymosan area in confocal images of neutrophils. Stacks of 12–16 confocal images (each section with 0.52 μM thickness) were collected with an LSM 510 laser-scanning confocal microscope (Carl Zeiss, Jena, Germany). Image analysis was performed using LSM Image Examiner software (Carl Zeiss).
Statistical analysis
All of the statistical data were analyzed by GraphPad Prism (GraphPad Software Inc., San Diego, CA, USA). Survival data were analyzed by the log-rank test. All other data were evaluated either by two-tailed Student’s t-test or by ANOVA. Bonferroni test was used for post-hoc comparison. p < 0.05 was considered to indicate statistical significance.
Go to :
RESULTS
Glycine enhancement of neutrophil bactericidal activity
We examined whether glycine enhances neutrophil bactericidal activity. As shown in Fig. 1A, glycine enhanced neutrophil bactericidal activity in a concentration-dependent manner, with a peak at 500 μM (EC50 = 238 μM). The glycine enhancement of neutrophil bactericidal activity was blocked by strychnine (a specific GlyR antagonist, 1 μM), indicating the involvement of GlyR in the action of glycine (Fig. 1A). Next, we examined whether systemic administration of glycine increases bacterial clearance in vivo. As shown in Fig. 1B, subcutaneous injection of glycine (30 mg/kg) at 30 min after intraperitoneal injection of E. coli (5 × 108) significantly decreased the number of CFUs in peritoneal lavage fluid collected 8 h after E. coli injection. The effect of glycine was blocked by pretreatment with strychnine (0.4 mg/kg, s.c.) (Fig. 1B).
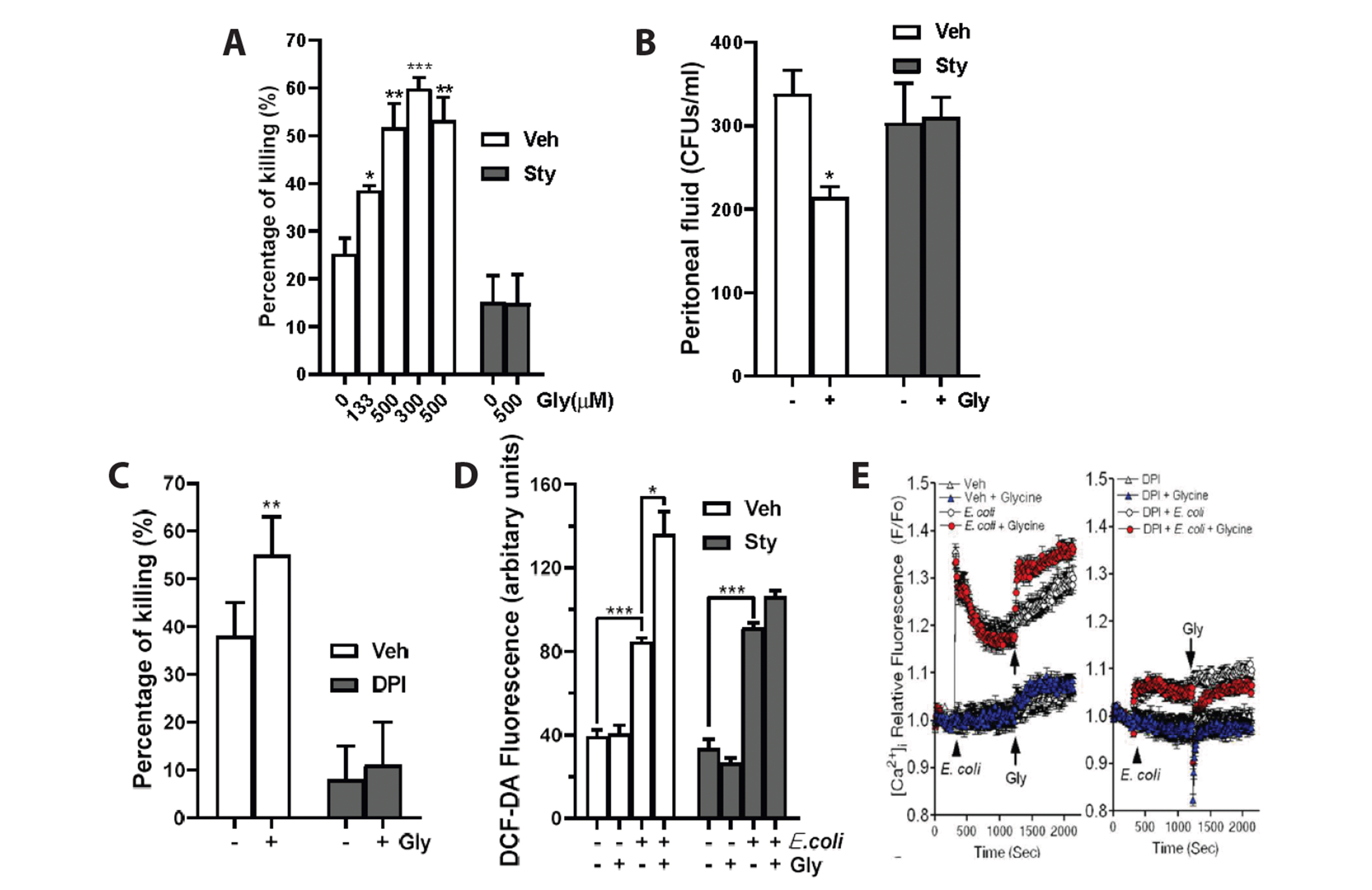 | Fig. 1Glycine increases neutrophil bactericidal activity via enhanced intracellular reactive oxygen species (ROS) production.(A) Glycine concentration-dependently increases neutrophil bactericidal activity. Various concentrations of glycine were added to HBSS. Strychnine blocked glycine-induced enhancement of neutrophil bactericidal activity. The average of three experiments is shown (mean ± SE). (B) Glycine increases the clearance of intraperitoneally injected E. coli in mice. Mice were pretreated subcutaneously with PBS or strychnine (0.4 mg/kg) 30 min before an i.p. injection of E. coli (5 × 108). At 30 min after E. coli injection, vehicle (PBS) or glycine (30 mg/kg) was subcutaneously injected. Eight hours after E. coli injection, mice were killed and the number of CFUs in peritoneal lavage fluid was counted. n = 9−10 per group. *p < 0.05; **p < 0.01; ***p < 0.001 by Student’s t-test. (C) Effect of DPI on glycine-induced enhancement of bactericidal activity. Neutrophils were pretreated with DPI (1 μM) for 1 h before glycine (500 μM) treatment in HBSS medium. (D) Effects of glycine on ROS generation in neutrophils. Neutrophils were exposed to E. coli for 20 min. Unphagocytosed E. coli was washed away and neutrophils were further incubated with or without glycine (500 μM) for 15 min. ROS production in neutrophils was measured with DCF-DA. (E) GIyR-ROS-Ca2+ signaling pathway. (Left) Effect of glycine on [Ca2+]i in neutrophils exposed to E. coli. Neutrophils were exposed to E. coli for 20 min, and further incubated with or without glycine (1 mM). (Right) Neutrophils were pretreated with DPI (1 μM) for 15 min before E. coli exposure. CFUs, colony forming units; DPI, diphenylene iodonium; DCF-DA, 2’,7’-dichlorodihydrofluorescein diacetate; Gly, glycine; GlyR, glycine receptor.
|
Glycine-induced enhancement of neutrophil bactericidal activity is dependent on enhanced intracellular ROS production
NADPH oxidase and its products ROS play a pivotal role in neutrophil bactericidal activity [18,19]. Thus, we examined the effect of DPI (1 μM), an inhibitor of NADPH oxidase, on the glycine enhancement of bactericidal activity. As expected, basal bactericidal activity was decreased by DPI (Fig. 1C). Remarkably, the glycine-induced enhancement of bactericidal activity was completely blocked by DPI (Fig. 1C). With the use of DCF-DA, we next examined whether glycine enhances intracellular ROS production in neutrophils that have phagocytosed E. coli. Glycine did not affect ROS production in resting neutrophils (Fig. 1D), but it markedly enhanced intracellular ROS production in neutrophils that had phagocytosed E. coli in a strychnine-sensitive manner (Fig. 1D).
GIyR-ROS-Ca2+ signaling pathway
[Ca2+]i in neutrophils is critical for bactericidal activity [20]; thus, we examined whether glycine enhances [Ca2+]i in neutrophils that have phagocytosed E. coli via the enhancement of ROS production. Glycine (1 mM) did not affect [Ca2+]i in resting neutrophils (Fig. 1E left panel, lower tracings). However, glycine (1 mM) markedly enhanced [Ca2+]i in neutrophils that had phagocytosed E. coli (Fig. 1E left panel, upper tracings) in a DPI-sensitive manner (Fig. 1E right panel).
Glycine induces 36CI– uptake and [Ca2+]i increase in neutrophils pretreated with PMA
To further confirm the GlyR-ROS-Ca2+ signaling pathway, we used PMA (a protein kinase C activator)-treated neutrophils. PMA (2 μg/ml) induces the release of neutrophil granules and the resultant translocation of proteins in the granule membrane to the plasma membrane [21]. In the PMA-treated neutrophils, glycine effectively increased 36CI– influx (Fig. 2A) and ROS production (Fig. 2B) in a strychnine-sensitive manner. In the PMA-treated neutrophils, glycine-induced a marked increase in [Ca2+]i (lower tracings), which was inhibited by DPI pretreatment (middle tracings) (Fig. 2C). These results further confirm the GlyR-ROS-Ca2+ signaling pathway in activated neutrophils.
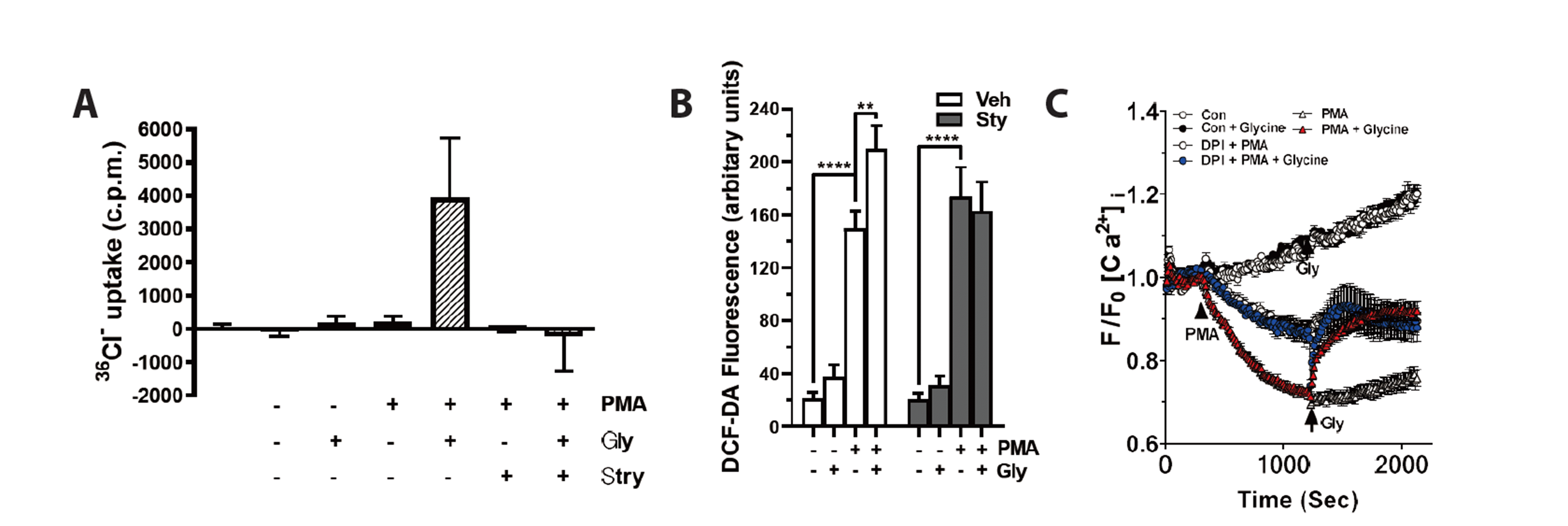 | Fig. 2Glycine induces 36CI– uptake and [Ca2+]i increase in neutrophils pretreated with phorbol myristate acetate (PMA).(A) 36CI– influx in response to glycine (1 mM) in neutrophils. Neutrophils were incubated with 2 μg/ml PMA for 20 min. Uptake was initiated by adding 36CI– solution and influx was terminated by the addition of 4 ml of ice-cold HEPES butter and rapid filtration. Neutrophils were placed in scintillation vials and solubilized with 0.2 N NaOH for 2 h. EcoLume was added and radioactivity was measured with a Beckman LS-5801 liquid scintillator. (B) Neutrophils were pretreated with 2 μg/ml PMA for 20 min, and glycine (500 μM)-induced reactive oxygen species (ROS) generation was measured. (C) Effect of glycine (1 mM) on [Ca2+]i in vehicle- or PMA-pretreated neutrophils. Neutrophils were pretreated with diphenylene iodonium (DPI) (1 μM) for 15 min before PMA exposure. (A–C) The average of three experiments is shown (mean ± SE). *p < 0.05; **p < 0.01;***p < 0.001 by Student’s t-test. DCF-DA, 2’,7’-dichlorodihydrofluorescein diacetate.
|
Effects of TRPM2-specific shRNA on glycine (1 mM)-induced increase in [Ca2+]i in neutrophils, and on glycine (500 μM)-induced enhancement of bactericidal activity
The Ca2+-permeable cation channel TRPM2 is abundantly expressed in neutrophils [22,23]. TRPM2 channel activity is well known to be increased by ROS [24,25]. Therefore, we examined the involvement of TRPM2 in the effects of glycine. The treatment of neutrophils with TRPM2-specific shRNA effectively decreased the glycine-induced increases in [Ca2+]i (Fig. 3A), as well as the bactericidal activity in neutrophils (Fig. 3B). Taken together, these results indicate the presence of a GlyR-ROS-Ca2+ signaling pathway in neutrophils that have phagocytosed E. coli.
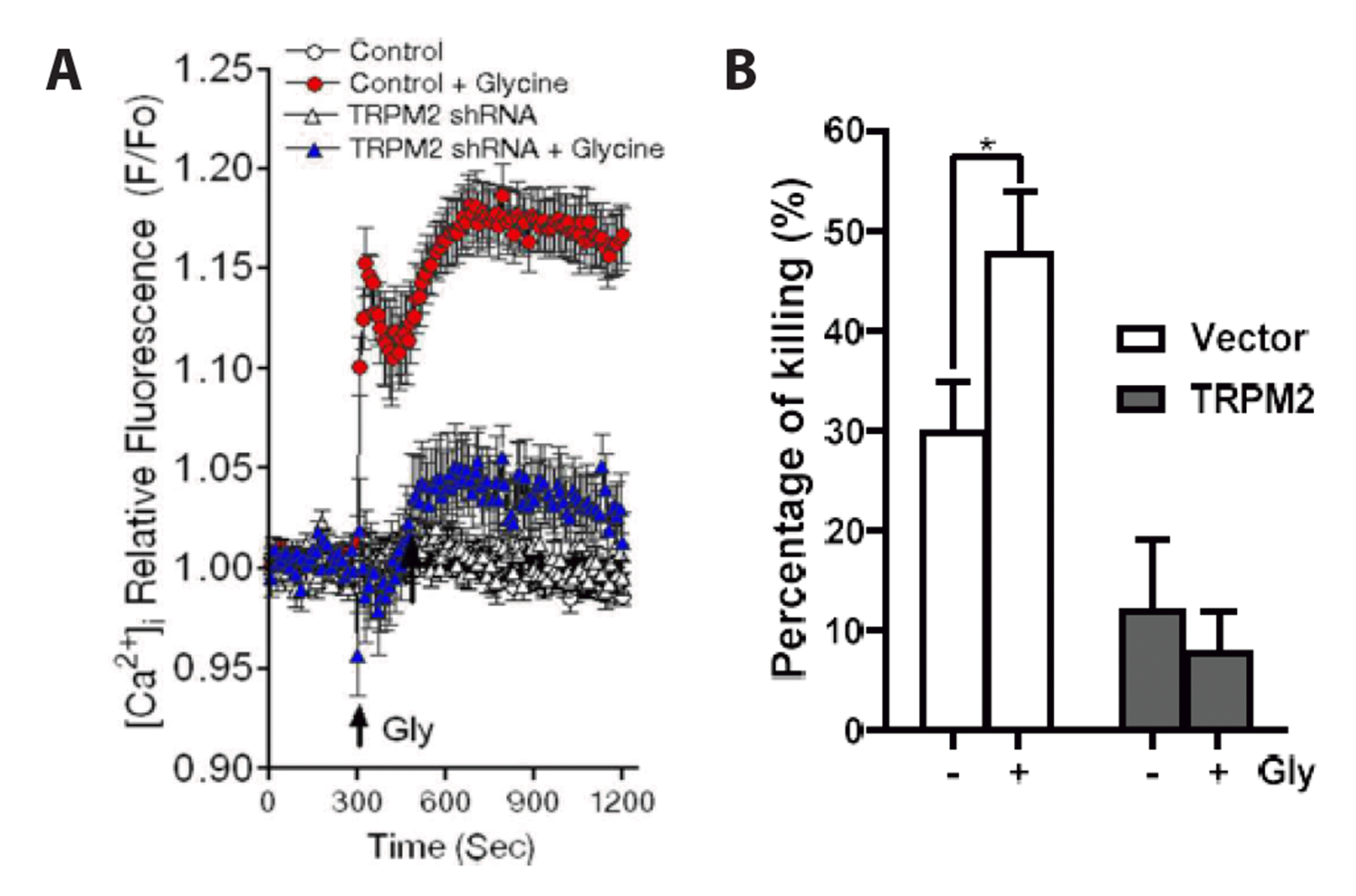 | Fig. 3Effects of TRPM2-specific shRNA on glycine (1 mM)-induced increase in [Ca2+]i in neutrophils (A), and on glycine (500μM)-induced enhancement of bactericidal activity (B).Neutrophils were transfected with shRNA against TRPM2. After 24 h, the effects of glycine on [Ca2+]i. in neutrophils (A) and bactericidal activity (B) were assayed. The average of three experiments is shown (mean ± SE). *p < 0.05 by Student’s t-test.
|
Subcellular localization of GlyRα2 in human neutrophils
Previously, we showed the presence of GlyR in human neutrophils [13]. However, the intracellular localization of GlyR in human neutrophils is unknown. To examine the localization of GlyRα2 in human neutrophils, cellular fractionation was carried out using nitrogen cavitation and subcellular fractionation methods [21,26-28]. Purified neutrophils were pressurized with N2 for 20 min at 3350 psi and the supernatant was obtained. Density gradients were prepared over 9 ml of Percoll (1.120 g/ml, 1.090 g/ml, and 1.050 g/ml). The Percoll density gradient following centrifugation revealed four distinct bands. The particulate fractions are referred to as α, β1, β2 and γ (less dense), which represent azurophil granules, specific granules, gelatinase granules, and plasma membrane with secretory vesicles, respectively. Fig. 4A clearly shows the localization of GlyRα2 in the γ-band, the fraction for secretory vesicles and plasma membranes.
 | Fig. 4Subcellular localization of glycine receptor (GlyR)α2 in human neutrophils.(A) Isolated neutrophils were pressurized with N2 gas for 20 min at 350 psi, and cavitated neutrophils were loaded onto three separate Percoll density gradients. After centrifugation, four distinct bands were observed from the bottom, designated as α-band (azurophil granules), β1-band (specific granules), β2-band (tertiary granules), and γ-band (secretory vesicles and plasma membranes). For negative controls, non-transfected HEK 293 cells were used. For positive controls, neutrophils and GlyRα2-transfected HEK 293 cells (HEK+α2) were used. Western blot revealed the localization of GlyRα2 in the γ-band (secretory vesicles and plasma membranes), with or without blocking peptide against GlyRα2. Exposure of neutrophils to E. coli or phorbol myristate acetate (PMA) effectively translocated GlyR to the plasma membrane. Representative fluorescein isothiocyanate (FITC) histograms on membrane expression of GlyRα2 in response to E. coli (B) or PMA (C). Neutrophils were treated with E. coli or PMA for 5 min. After fixation, neutrophils were stained with FITC-conjugated anti-GlyRα2 antibody for 30 min. (D) A representative FACS histogram on membrane expression of flavocytochrome b588 in response to E. coli. Neutrophils were exposed to E. coli for 20 min.
|
Exposure of neutrophils to PMA or E. coli effectively translocates GlyR to the plasma membrane (Fig. 4B, C). The NADPH oxidase comprises cytosolic components and membrane-associated cytochrome b558 (gp91 and phox22 subunits of NADPH oxidase) [29]. In resting neutrophils, 15% of cytochrome b558 is localized in the plasma membrane and the remaining 85% in the intracellular granular membrane, chiefly specific granules, and upon activation, translocates to the plasma membrane [29]. As expected, the exposure of neutrophils to E. coli induced the translocation of cytochrome b538 to the plasma membrane (Fig. 4D), conferring the basis for cooperation of GlyR and NADPH oxidase.
Glycine enhances azurophil granule-phagosome fusion via p38 MAPK
Next, we attempted to clarify the cellular events occurring after [Ca2+]i increase, regarding enhancement of microbicidal activity. Neutrophil azurophil granules contain myeloperoxidase and proteases, which play a critical role in killing microbes [30]. Therefore, azurophil granule-phagosome fusion [31,32] and neutrophil proteases (such as neutrophil elastase and cathepsin G contained mostly in azurophil granules) [33-36] are critical for neutrophil bactericidal activity. Furthermore, it was previously demonstrated that azurophil granule release in neutrophils is linked to fluid-phase pinocytosis [17]. Thus, we examined whether glycine enhances fluid-phase pinocytosis (by measuring uptake of Lucifer yellow, a water-soluble, membrane-impermeable fluorescent dye) and azurophil granule-phagosome fusion (by measuring the area of co-localization between CD63 [an azurophil granule marker] and zymosan particles, which was divided by the total intracellular zymosan area in confocal images of neutrophils). As shown in Fig. 5A and B, glycine effectively increased Lucifer yellow uptake and azurophil granule-phagosome fusion, which were blocked by strychnine and DPI. These results indicate that glycine-induced enhancement of ROS production (Fig. 1D) is critically involved in the enhancement of Lucifer yellow uptake and azurophil granule-phagosome fusion. Protease inhibitors completely blocked the enhancement of neutrophil bactericidal activity induced by glycine (Fig. 5C).
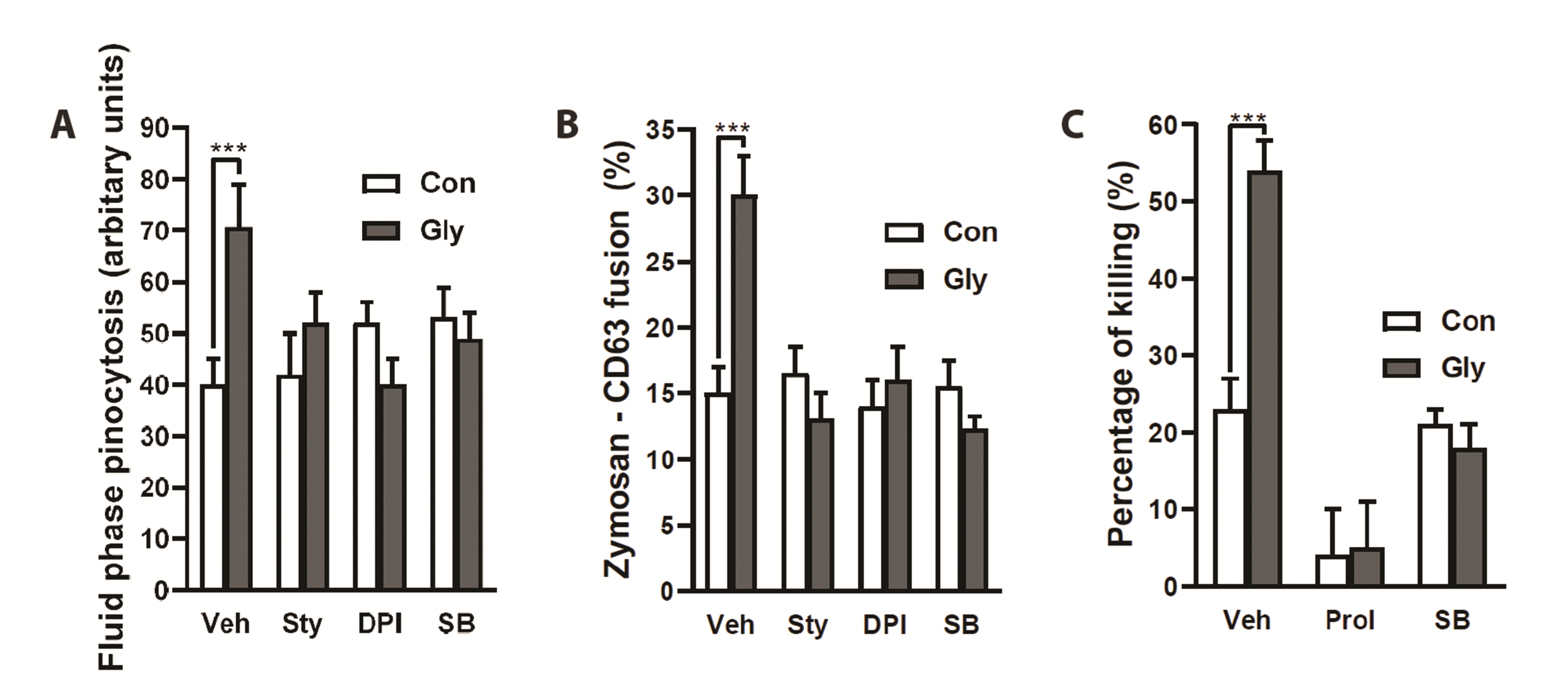 | Fig. 5Glycine enhances azurophil granule-phagosome fusion via p38 mitogen-activated protein kinase (MAPK).(A) Effects of strychnine, diphenylene iodonium (DPI) and SB203580 on glycine-induced enhancement of fluid-phase pinocytosis. Uptake of Lucifer yellow in neutrophils was measured as fluid-phase pinocytosis. Neutrophils were exposed to E. coli for 20 min. Unphagocytosed E. coli was washed away and neutrophils were further incubated with glycine (500 μM) for 15 min in the presence of 0.5 mg/ml Lucifer yellow. Strychnine (1 μM), DPI (1 μM) or SB203580 (10 μM) was added during 15 min of incubation. Neutrophils were lysed with ice-cold distilled water for 1 h and fluorescence was measured. (B) Effects of strychnine, DPI, and SB203580 on zymosan-CD63 fusion. Neutrophils were allowed to ingest Texas Red-conjugated zymosan for 20 min. Unphagocytosed zymosan particles were washed away and neutrophils were further incubated with glycine in the presence of strychnine, DPI, or SB203580. Neutrophils were fixed and stained with anti-CD63 antibody. Stacks of 12–16 confocal images were collected with an LSM 510 laser-scanning confocal microscope (Carl Zeiss, Jena, Germany) and analyzed by LSM Image Examiner software (Carl Zeiss). The fusion of CD63 and zymosan particles was analyzed by measuring the area of co-localization between CD63 and zymosan particles, which was divided by total intracellular zymosan area in confocal images of neutrophils at each slice of 0.52 μm in thickness. (C) Effects of protease inhibitor cocktail and SB203580 on glycine-induced enhancement of bactericidal activity. Neutrophils were pretreated with a protease inhibitor cocktail or SB203580 for 1 h before E. coli exposure. (A–C) The average of three experiments is shown (mean ± SE). ***p < 0.001 by Student’s t-test.
|
p38 MAPK is known to be involved in phagosomal maturation in macrophages [37], fluid-phase pinocytosis [38], and translocation of the intracellular cytochrome b558 (a component of NADPH oxidase) to the plasma membrane in neutrophils [39]. Thus, we examined the effect of the p38 MAPK inhibitor SB203580 on glycine-induced increases in Lucifer yellow uptake, azurophil granule-phagosome fusion, and bactericidal activity. As shown in Fig. 5A–C, SB203580 completely blocked glycine-induced enhancement of all of these parameters. The lack of effect of p38 MAPK inhibition by SB203580 on basal microbicidal activity (Fig. 5C) is in agreement with previous reports [40,41].
Go to :
DISCUSSION
In this study, we found a novel mechanism for the enhancement of neutrophil bactericidal activity (Fig. 6), in which intracellular GlyRα2 translocates to the plasma membrane (Fig. 4B, C) and glycine acts via GlyRα2 to induce chloride influx (Fig. 2A). The subsequent signaling mechanism couples the chloride influx to the enhancement of NADPH oxidase activity (the cytochrome b558 component Nox also translocates to the plasma membrane in neutrophils that have phagocytosed E. coli), resulting in increased ROS production (Fig. 1D). This elevated ROS production increases TRPM2 channel activity, resulting in [Ca2+]i increase (Figs. 1E left panel, 2C, 3A). Subsequent cascading events include p38 MAPK activation, and enhancement of fluid-phase pinocytosis, azurophil granule-phagosome fusion, and bactericidal activity (Fig. 5). However, it is to be noted that the GlyRα2/TRPM2/p38 MAPK signal is not critically involved in basal bactericidal activity, but can be mobilized for boosting the bactericidal activity of neutrophils when neutrophils are stimulated by glycine.
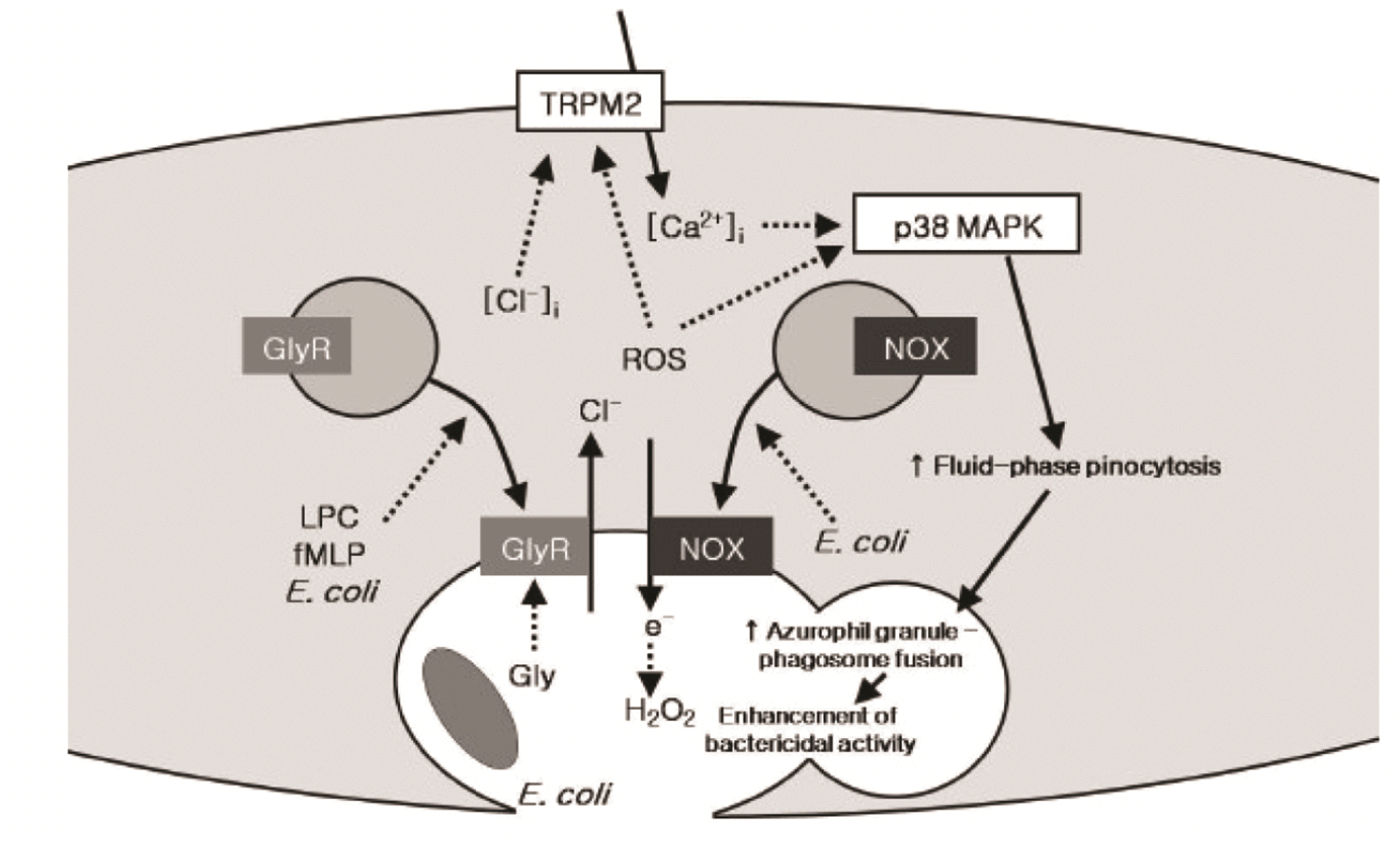
NADPH oxidase is essential for neutrophil microbicidal activity [29]. We detected glycine/GlyR-mediated enhancement of NADPH oxidase activity in E. coli-treated neutrophils, but not in resting neutrophils (Fig. 1D), indicating that GlyR is not involved in the basal phagocytosis-associated respiratory burst but only in the enhancement of it induced by supplemental glycine. Impaired NADPH oxidase activity is known to be accompanied by impairment in release of azurophil granules that contain various proteases, and protease activity; both chronic granulomatous disease neutrophils and DPI-treated neutrophils show impairment in azurophil granule release [42-44] and in the digestion of killed E. coli [36,45,46]. We found that ROS was essential in glycine-induced azurophil granule-phagosome fusion via the Ca2+/p38 MAPK/fluid-phase pinocytosis/azurophil granule-phagosome fusion pathway. This finding reveals a clear mechanism for the inter-relationship between ROS and proteases in neutrophil bactericidal activity. Specifically, the blocking effect of DPI on glycine-induced enhancement of azurophil granule-phagosome fusion (Fig. 5B) provides a mechanism for the cooperation of ROS and protease action, two important microbicidal effectors of neutrophils [47].
Interestingly, LPC translocates GlyR to the plasma membrane, and induces glycine/GlyR-mediated enhancement of neutrophil bactericidal activity [13]. These results suggest that the translocation of intracellular GlyR to the neutrophil plasma membrane is a commonality of signaling for neutrophil stimulators. However, a difference between glycine (Fig. 1C) and LPC [13] was noted in the sensitivity of their bactericidal activity-enhancing effect on DPI. Further studies are needed to elucidate their shared as well as disparate intracellular signaling pathways.
The administration of glycine (30 mg/kg) to mice that had been intraperitoneally given E. coli enhanced bacterial clearance (Fig. 1B). The reported physiological plasma concentration of glycine (~ 250 μM) in mice [48] as well as in humans [49] is below 500 μM, the optimum glycine concentration for bactericidal activity (Fig. 1A). In fact, strychnine affected neither basal neutrophil bactericidal activity in vitro (where the glycine concentration in RPMI medium is 133 μM) (Fig. 1A), nor basal bacterial clearance in mice challenged with the intraperitoneal administration of E. coli (Fig. 1B), probably due to an insufficient concentration of glycine. Therefore, boosting GlyR activation in neutrophils by glycine supplementation (Fig. 1A, B) could be beneficial for treating of serious microbial infectious disorders, especially when direct-acting antimicrobials are not sufficiently effective. Together, our findings suggest neutrophil GlyRα2 as a novel target in the fight against microbes.
Go to :
 XML Download
XML Download