

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Glucagon-like peptide-1 (GLP-1) secreted from intestinal enteroendocrine L-cells is an incretin that can augment glucose-dependent insulin secretion [1]. GLP-1 is known to stimulate proinsulin gene expression and pancreatic β-cell proliferation [2,3]. Promoting insulin secretin is a crucial goal when treating type 2 diabetes mellitus. Thus, stimulating endogenous release of GLP-1 has become one of therapeutic targets for diabetes [4,5]. A number of cellular models, such as isolated primary intestinal L-cell and enteroendocrine cell lines STC-1, GLUTag, and NCI-H716 cells, have been used to study the regulation of GLP-1 secretion [6,7]. These cellular models provide useful information concerning signaling pathways that regulate GLP-1 secretion. NCI-H716 cells derived from poorly differentiated adenocarcinoma of human cecum have potent GLP-1 secreting properties. Nutrients, hormones, and neurotransmitters are known to control the release of GLP-1 in physiological conditions [8,9]. It has been reported that muscarinic agonist can stimulate GLP-1 secretion in NCI-H716 cells, and that such secretion is closely linked to G-protein coupled receptor activations and elevated cytosolic Ca2+ signals [10-12]. Although GLP-1 secretion plays an important role in blood glucose control as mentioned above, the detail stimulus-secretion pathway in NCI-H716 cells is not completely understood yet.
Mammalian Na+/Ca2+ exchanger (NCX) is a bi-directional Ca2+ transporter that contributes to Ca2+ homeostasis of multiple tissues such as cardiac muscle, skeletal muscle, brain, and kidney [13]. Mammalian cells are known to express three types of NCX isoforms [14,15]. NCX1 is broadly expressed in various cells. It is particularly abundant in the heart, brain, and kidney. However, NCX2 is locally expressed in the brain and NCX3 is limitedly expressed in the brain and skeletal muscles. The direction of Ca2+ movement depends on electrical and chemical gradients across the membrane. The forward mode of NCX generates an inward Na+ current and an outward Ca2+ current while the reverse-mode NCX promotes Na+ efflux and Ca2+ influx in plasma membrane. It is usually accepted that the forward NCX is favored at resting membrane potential and that reverse NCX (rNCX) may operate when the cell is physiologically depolarized [13-16]. Na+ dependency of GLP-1 secretion has been suggested because glucose-induced GLP-1 secretion can be blocked by replacing extracellular Na+ with N-methyl-d-glucamine (NMG+) in GLUTag and intestinal L-cells [7,17]. Since the secretion of GLP-1 in response to nutrient or neural stimulation can be triggered by elevation of cytosolic Ca2+, extracellular Na+ may be involved in the stimulus-secretion pathway of NCI-H716 cells. Therefore, the purpose of this study was to investigate the existence and the role of reverse Na+/Ca2+ exchanger on Ca2+ mobilization under cholinergic stimulation in NCI-H716 cells. Here we report that NCX1 is present on the plasma membrane and that rNCX contributes to calcium entry for calcium oscillation under cholinergic stimulation in NCI-H716, an enteroendocrine GLP-1 secreting cell line.
Go to : 
METHODS
Culture of NCI-H716 cells
Human enteroendocrine NCI-H716 cells were obtained from the American Type Culture Collection (Manassas, VA, USA). Cells were maintained as a suspension culture in RPMI 1640 at 37°C in a humidified atmosphere containing 5% CO2 as described previously [18]. Two days before experiments, endocrine differentiation and cell adhesion cells were initiated by seeding cells onto cover glass in 6-well culture plates coated with Matrigel (Becton Dickinson Co., Bedford, MA, USA) in high-glucose DMEM. On the day of experiments, to stabilize cultured cells and measure cytosolic Ca2+, cells were resuspended with HEPES-buffered physiological saline containing 137 mM NaCl, 4.7 mM KCl, 0.56 mM MgCl2, 1 mM Na2HPO4, 1.28 mM CaCl2, 10 mM HEPES, and 5.5 mM glucose (pH 7.4).
Intracellular Ca2+ measurements
Cultured cells were loaded with 5 μM Fura-2/AM, a low affinity Ca2+ sensitive dye, for 30 min at room temperature to measure intracellular Ca2+. Fura-2/AM loaded cells were continuously perfused with HEPES-buffered physiological saline. The perfusion rate was consistently controlled to be 1 ml/min using an electronically controlled perfusion system (Warner Instrument, Hamden, CT, USA). Intracellular Ca2+ levels were monitored using a TILL Photonics imaging system (Till photonics, Pleasanton, CA, USA). Fura-2/AM loaded cells were excited alternately with light at 340 nm and 380 nm using a Polychrome V monochrometer (TILL Photonics). Fluorescence images emitted at 510 nm through 40× fluorescence objective lens were captured using a Cool-SNAP HQ2 camera (Photometrics, Tuscon, AZ, USA) attached to an Olympus IX71 inverted microscope (Olympus, Tokyo, Japan) [19].
Western blots and immunocytochemistry
Protein samples were prepared from cultured NCI-H716 cells. Briefly, cells were pelleted and homogenized in an ice-cold lysis buffer containing 50 mM Tris/HCl, 2 mM EDTA, 10 mM EGTA, 5 mM DTT, 250 mM sucrose, 1% Triton X–100, and protease inhibitors (pH 7.5). Protein concentrations were measured using Bradford’s protein assay reagents (Bio-Rad Laboratories, Hercules, CA, USA). Samples were separated by 10% SDS-PAGE and transferred to nitrocellulose membrane at room temperature. After overnight incubation with polyclonal NCX1 primary antibody (Santa Cruz Biotechnology, Santa Cruz, CA, USA) at 4°C, membranes were then incubated with goat anti-rabbit IgG horseradish peroxidase secondary antibodies for 1 h at 37°C. Bands were visualized using enhanced chemiluminescence. To determine distributions of NCX1, prepared NCI-H716 cells were placed on a poly-L-lysine coated glass coverslip for 10 min at room temperature prior to fixation with 2% formaldehyde in PBS solution. After fixation, nonspecific bindings were blocked by 1 h incubation in 3% BSA medium. The immunocytochemistry was performed using a primary NCX1 antibody and Cy-2 goat anti-rabbit secondary antibody. Immunofluorescence images for NCX1 were collected with an Olympus IX71 inverted microscope.
Data analysis
Change of cytosolic Ca2+ was expressed as a representative trace in individual cells. Summated results were presented as mean ± SE. Differences were considered significant when the p-value was less than 0.05 using Student’s t-test.
Go to :
RESULTS
Cholinergic stimulation induced cytosolic Ca2+ oscillation
Initial experiments were performed to investigate if cholinergic stimuli could generate oscillatory cytosolic Ca2+ signals in NCI-H716 cells. Changes of intracellular Ca2+ concentration were monitored using various concentrations of carbamylcholine (CCh, 0.3 μM–1 mM), a long-acting acetylcholine analog in the presence of 1.28 mM extracellular Ca2+. As shown in Fig. 1A, the perfusion above 1 μM of CCh for 100 sec evoked an initial Ca2+ peak. Oscillatory Ca2+ signals then continued. The maximum effect of initial Ca2+ peak was observed at 100 μM of CCh. Half maximum dosage of initial Ca2+ peak was 0.85 ± 0.09 μM of CCh. The maximum frequency of Ca2+ oscillation was 12.02 ± 0.89 peaks/min. Half maximum effect was observed at 2.98 ± 0.47 μM of CCh (Fig. 1B). These results suggest that cholinergic stimulation could sufficiently generate oscillatory Ca2+ signals in NCI-H716 cells. Therefore, we used CCh at a concentration of 10 μM in the following experiments to generate repetitive Ca2+ oscillation. The following experiment was performed to determine whether voltage-operated Ca2+ channel (VOC) was involved in CCh-induced calcium oscillation in NCI-H716 cells. As shown in Fig. 2A and 2B, Ca2+ oscillation signal induced by CCh was not suppressed by nifedipine, a VOC blocker. These results indicate that VOC does not participate in CCh-stimulated calcium influx in NCI-H716 cells.
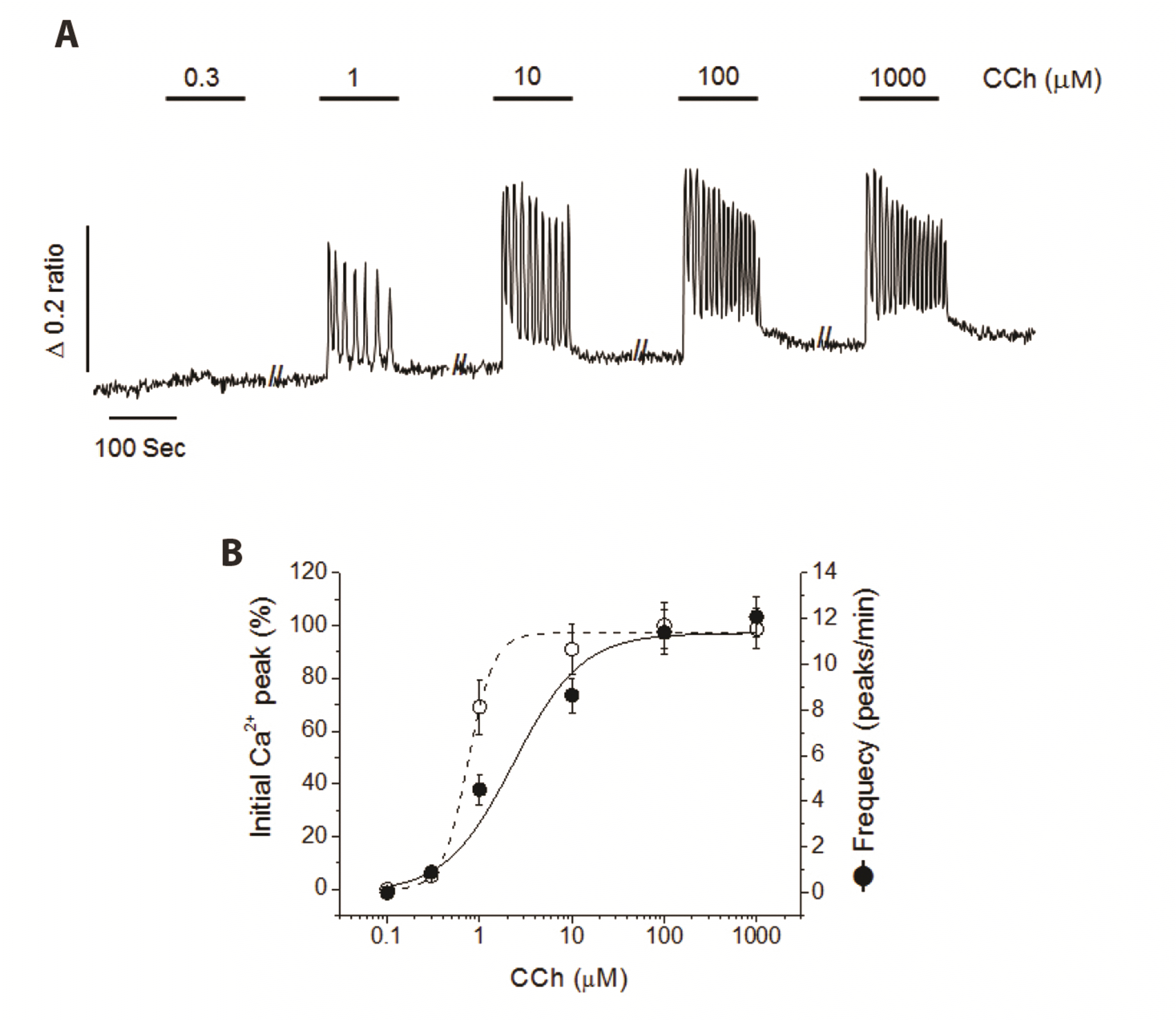 | Fig. 1Dose-dependent Ca2+ oscillation induced by carbamylcholine (CCh) in NCI-H716 cells.(A) Representative cytosolic Ca2+ oscillation obtained from stepwise increase of various concentration of CCh (0.3 μM–1 mM) every 100 sec. (B) CCh-induced initial Ca2+ peak (% of maximum) and frequency of Ca2+ oscillation (peaks/min). Cytosolic Ca2+ measurement was obtained from seven separate experiments in fura-2 loaded NCI-H716 cells. CCh significantly stimulated initial Ca2+ peak and frequency of Ca2+ oscillation, dose-dependently.
|
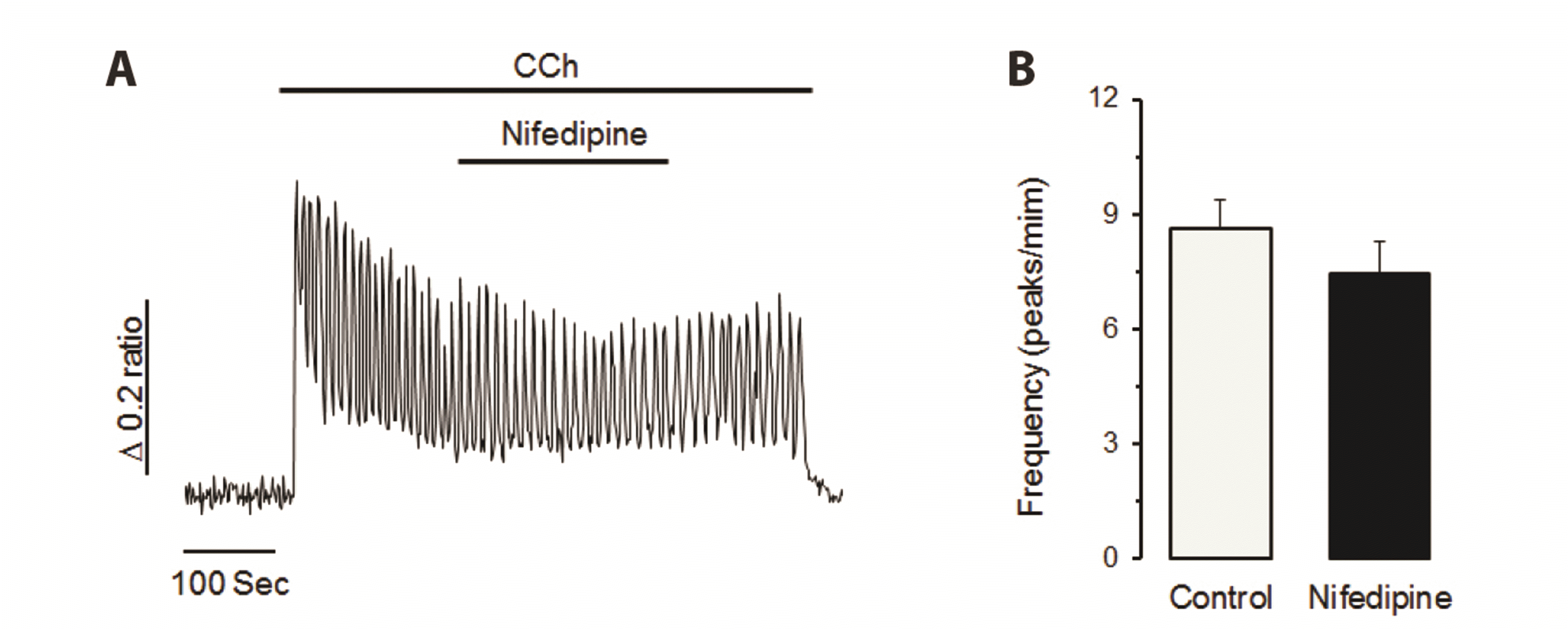 | Fig. 2Voltage-operated Ca2+ channel (VOC) does not contribute to carbamylcholine (CCh)-induced Ca2+ entry process in NCI-H716 cells.(A) Nifedipine, a VOC antagonist, failed to change CCh-induced Ca2+ oscillation in NCI-H716 cells. (B) CCh-induced frequency of Ca2+ oscillation was not changed by nifedipine treatment. Similar results were obtained from six separate experiments in NCI-H716 cells. VOC might not be involved in the calcium influx in CCh-stimulated NCI-H716 cells.
|
Elimination of extracellular Na+ terminates CCh-induced Ca2+ oscillation
Next, we determined whether Na+/Ca2+ exchanger could contribute to CCh-induced oscillatory Ca2+ signals in NCI-H716 cells. Preferentially, we eliminated extracellular Na+ by replacing Na+ with NMG+ or Li+. As shown in Fig. 3A, the replacement of Na+ with NMG+ resulted in a slight enhancement of Ca2+ oscillation pattern in the initial state, followed continuous decreases. Cytosolic Ca2+ oscillation was complete terminated at about 120 sec after Na+ elimination. Reintroduction of extracellular Na+ resulted in a slow restoration of oscillatory Ca2+ signals. When Na+ was replaced with Li+, cytosolic Ca2+ oscillation was slowly terminated similar to the replacement of Na+ with NMG+. Such termination was then also partially recovered by Na+ reperfusion (Fig. 3B). these data indicate that extracellular Na+ may be essential to Ca2+ influx for the generation of oscillatory Ca2+ signaling induced by muscarinic stimulation in the stimulus-secretion mechanism of NCI-H716 cells. Thus, the following experiment was planned to evaluate whether KB-R7943, a specific reverse-mode NCX blocker, could affect CCh-induced oscillatory Ca2+ signaling in NCI-H716 cells.
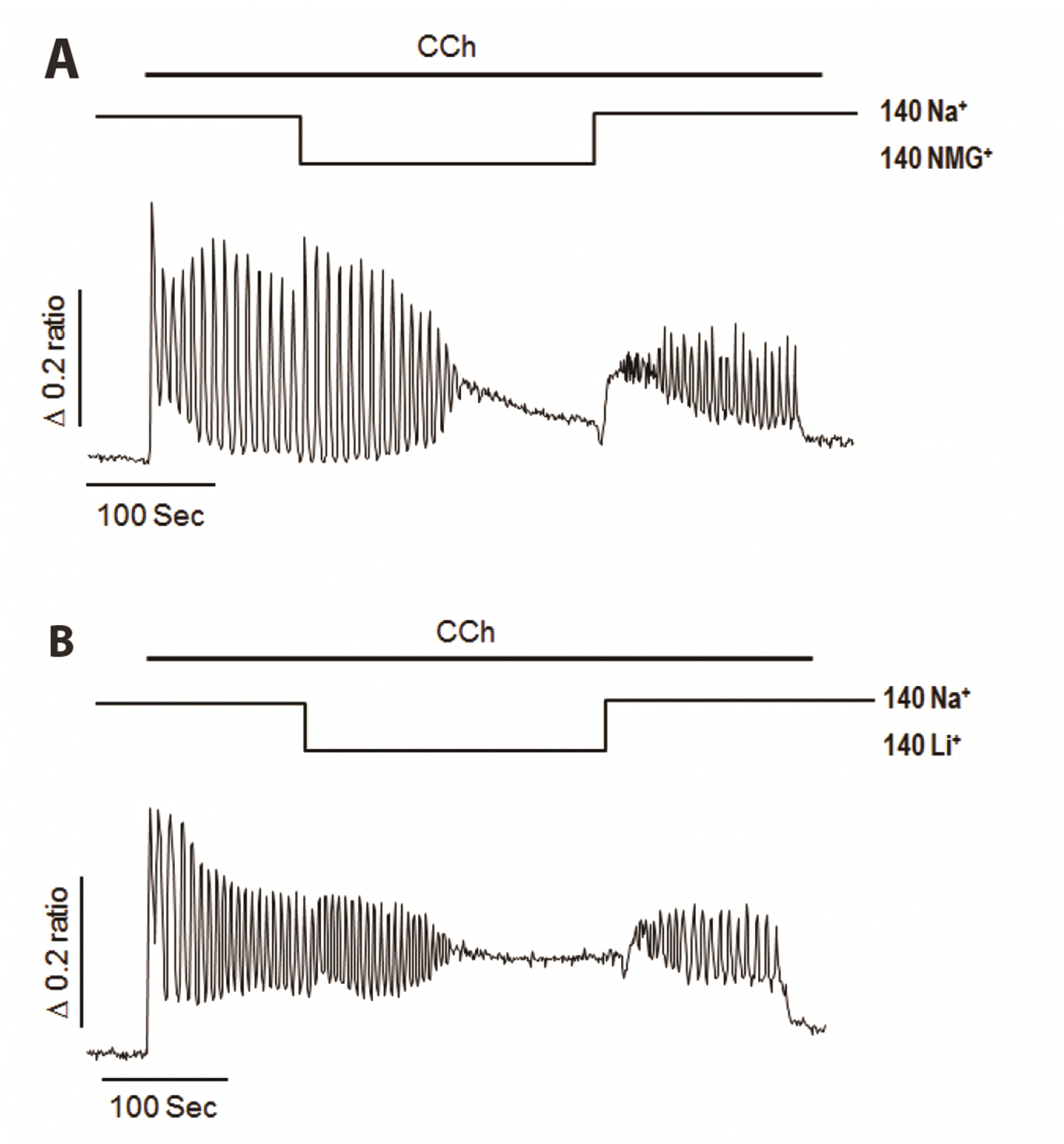 | Fig. 3Effects of extracellular free Na+ on carbamylcholine (CCh)-induced Ca2+ oscillation in NCI-H716 cells.(A) Substitution of Na+ with N-methyl-d-glucamine (NMG+) resulted in cessation of Ca2+ oscillation, which was restored by Na+ reperfusion. (B) Replacing Na+ with Li+ resulted in termination of cytosolic Ca2+ oscillation and restoration of oscillatory Ca2+ signals by Na+ reperfusion. Na+-dependent Ca2+ influx might be important for the generation of oscillatory Ca2+ signaling induced by muscarinic stimulation in stimulus-secretion mechanism of NCI-H716 cells.
|
Effects of KB-R7943 on CCh-induced Ca2+ oscillation
Addition of KB-R7943 remarkably abolished 10 μM CCh-induced Ca2+ oscillation. Such abolishment was recovered by withdrawing KB-R7943 (Fig. 4A). Similar results were observed when extracellular medium was changed to free Ca2+ buffer (Fig. 4B). In contrast, pretreatment of KB-R7943 had no obvious effect on the initial Ca2+ peak (Fig. 4C, D). Indeed, cholinergic stimuli known to generate initial cytosolic Ca2+ peaks with sustained oscillatory Ca2+ signals. Initial Ca2+ peaks are originated by inositol 1,4,5trisphospate-dependent calcium release from intracellular calcium stores and sustained Ca2+ oscillation in need of calcium influx from extracellular medium in secretory cells [20,21]. In the present study, KB-R7943 reduced only maintenance of oscillatory Ca2+ signaling without affecting initial Ca2+ transients. These results lead us to conclude that rNCX might contribute to the generation of CCh-induced cytosolic Ca2+ oscillation by modulating Ca2+ influx pathways from extracellular fluid in NCI-H716 cells.
 | Fig. 4Effects of KB-R7943 on carbamylcholine (CCh)-induced Ca2+ oscillation in NCI-H716 cells.(A) KB-R7943 significantly blocked CCh-induced Ca2+ oscillation in NCI-H716 cells. Ca2+ oscillation was restored after cessation of KB-R7943 perfusion. (B) Elimination of extracellular Ca2+ resulted in complete inhibition of CCh-induced Ca2+ oscillation. (C) Pretreatment of KB-R7943 failed to inhibit initial Ca2+ peak induced by CCh. (D) Effect KB-R7943 on CCh-induced initial Ca2+ peak obtained from seven separate experiments. rNCX might contribute to the generation of CCh-induced cytosolic Ca2+ oscillation by modulating Ca2+ influx pathway from extracellular fluid in NCI-H716 cells.
|
Expression of NCX1 proteins in NCI-H716 cells
In order to determine whether NCI-H716 cells could express NCX1 at the protein level, we performed Western blot analysis from prepared cell lysates using commercial primary antibodies to NCX1. NCX1 protein was detected in NCI-H716 cells at a band of about 120 kDa (Fig. 5A). Additional band of a smaller size at 67 kDa probably represent proteolytic product of NCX protein. Such degradation product of NCX has well described in other tissues [22]. In the immunocytochemical experiment on NCX1 protein localization, as shown in Fig. 5B, NCX1 protein was expressed on NCI-H716 cells. There was no non-specific background staining in the negative experiment without primary antibody.
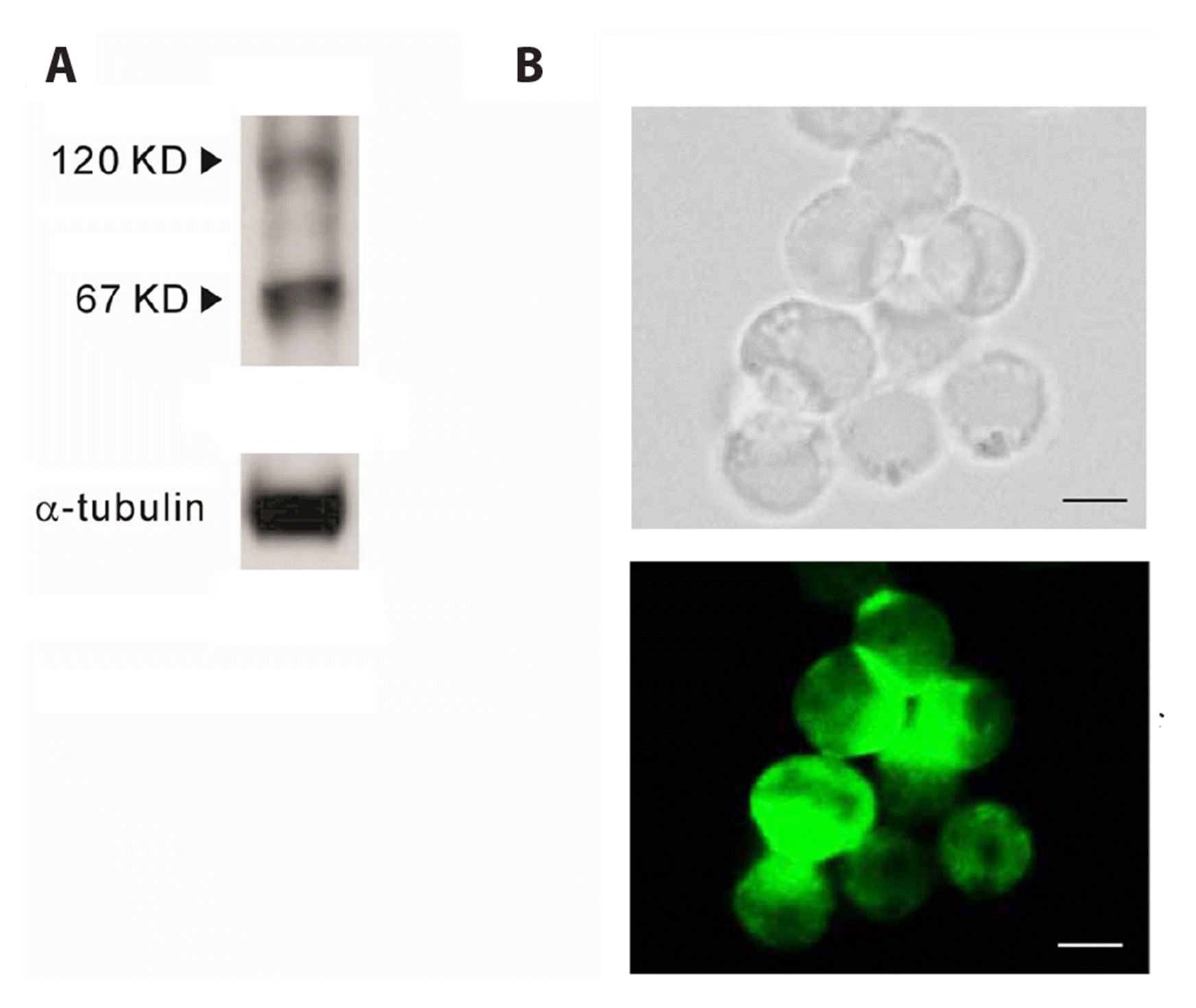 | Fig. 5Type 1 Na+/Ca2+ exchanger (NCX1) protein expression and distribution in NCI-H716 cells.(A) Western blot analysis of NCX1 expression. NCX1 protein was detected in NCI-H716 cells at band of about 120 kDa, and small sized proteolytic product band of 67 kDa was additionally detected. (B) Immunocytochemistry for NCX1 protein expression. Corresponding bright-field (top) and immunofluorescence (bottom) microscopy images showing the labelling of NCX1 protein. NCX1 protein was remarkably expressed on NCI-H716 cells, scale bars = 10 μm.
|
Go to :
DISCUSSION
The present study provides evidence that reverse-mode NCX might significantly contribute to acetylcholine-induced Ca2+ entry pathway of NCI-H716, a GLP-1 secreting cell line. Since GLP-1 is a well-known incretin hormone, many studies have been focused on stimulus-secretion mechanism of GLP-1 secreting cells to develop therapeutics for diabetes mellitus. In general, muscarinic receptors are closely linked to G-protein coupled receptor activation [10-12]. It is already known that muscarinic activation can enhance the secretion of GLP-1 in NCI-H716 cells [23]. Thus, muscarinic agonists can mobilize Ca2+ from internal stores through activation of InsP3 receptors, which subsequently activates store-operated Ca2+ entry from the external medium to refill depleted stores in NCI-H716 cells, similarly to other non-excitable epithelial cells [20,21]. However, these issues have not yet been addressed in detail. In this study, we confirmed that CCh could effectively generate oscillatory Ca2+ signals in NCI-H716 cells. The initial Ca2+ peak and the rhythmic Ca2+ frequency induced by CCh were also increased in a dose-dependent manner. These results support that muscarinic receptor-mediated Ca2+ signaling pathways which may act as a critical mechanism for regulating GLP-1 secretion are well operated in NCI-H716 cells.
In this study, for the first time, rhythmic calcium oscillation was observed in NCI-H716 cells by cholinergic stimulation. In general, repetitive and regular calcium oscillation can help intermittent message to include various information as the frequency, amplitude, and spatiotemporal characteristics of signals are different [24,25]. In part, calcium oscillation signals are known to be involved in various intracellular signaling processes including gene expression, exocytosis, and excitation-contraction coupling [25]. Intracellular Ca2+ oscillation signal is generated by a precise harmony between the mechanism of calcium mobilization and the mechanism of calcium extrusion [21]. Calcium mobilization can be achieved by the secretion of calcium from the intracellular calcium reservoir and the influx of calcium from the extracellular fluid. Calcium extrusion is achieved by refilling of calcium into the cell reservoir and excretion of calcium to the extracellular space. The calcium signal is precisely controlled by the balance of the outflow process. Therefore, calcium oscillation signal, which is well observed in NCI-H716 cells after cholinergic stimulation, is highly likely to act as an important regulator in the secretion of GLP-1. It is not yet clear which types of calcium channels are involved in changes in intracellular calcium concentration in NCI-H716 cells. Some studies have reported that depolarization of cell membrane in GLP-1 secreting cells, such as GLUTag cells or primary intestinal cells, can promote the secretion of GLP-1 [26,27]. However, the oscillatory calcium signal induced by CCh was unaffected by voltage-operated Ca2+ channel blocker nifedipine in this study. These results suggest that voltage-operated Ca2+ channels do not participate in CCh-induced oscillatory calcium signals in NCI-H716 cells. Recently, sodium dependency on GLP-1 secretion in intestinal L-cells has been suggested because glucose-induced GLP-1 secretion can be blocked by replacement of extracellular Na+ with NMG+ [7,17]. The present study also confirmed that when Na+ in the extracellular fluid was removed by substituting NMG+ or Li+, CCh-induced calcium oscillation disappeared. These results suggest that the calcium oscillation signal generated in NCI-H716 cells has a sodium dependence.
Na+/Ca2+ exchanger (NCX), a bi-directional Ca2+ transporter, is known to contribute to Ca2+ homeostasis of multiple tissues [13-16]. When cells are in an electrically or chemically activated state, reverse-mode NCX is known to promote Na+ efflux and Ca2+ influx through the plasma membrane. Muscarinic agonists not only can stimulate InsP3 production, but also can promote the opening of non-selective cation channels followed by elevation of membrane potentials in various cell types [28-30]. Thus, Na+ introduced through non-selective cation channels might play an important role in Ca2+ influx through rNCX under muscarinic stimulation in NCI-H716 cells. In this study, KB-R7943, a rNCX blocker, effectively attenuated CCh-induced cytosolic Ca2+ oscillation. Furthermore, depletion of cytosolic Na+ by replacement of Na+ with NMG+ or Li+ also slowly reduced and terminated CCh-induced cytosolic Ca2+ oscillation at about 120 sec after Na+ elimination. These results indicate that muscarinic agonists can activate Na+-dependent Ca2+ influx which might be closely linked to the generation of oscillatory Ca2+ signals in NCI-H716 cells. In addition, as a result of the experiment using cell immunostaining method, it was confirmed that the receptor of NCX1 was present on NCI-H716 cells.
Based on data presented here, it can be concluded that rNCX is very closely involved in generation of oscillatory Ca2+ signals promoted by CCh in NCI-H716 cells. Thus, rNCX might play a physiologically important role as a regulator of GLP-1 secretion in NCI-H716 cells.
Go to :
 XML Download
XML Download