

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Neutrophils are the first-line defense against invading pathogens [1,2]. Upon exposure to pathogens, neutrophil granules fuse with the plasma membrane and secrete their contents into the phagosome or extracellular space. However, the release of azurophil granules, which are rich in highly cytotoxic antimicrobial mediators (i.e., myeloperoxidase, cathepsin G, elastase, protease 3, defensin, and azurocidin), is tightly controlled [3,4]. Thus, azurophil granules are the least ready to be secreted among the various neutrophil granules [5]. For example, N-formyl-L-methionyl-L-leucyl-L-phenylalanine (fMLP), a major stimulator for neutrophils, alone cannot induce azurophil granule release. However, in the presence of cytochalasin B (CB), an inhibitor of actin polymerization, fMLP markedly increases azurophil granule release [6,7].
In various cell types, small G proteins are known to be key regulators of cytoskeletal remodeling [8]. Among them, the Rho-subfamily of GTPases such as Rho, Rac, and Cdc42, are important regulators of cytoskeletal dynamics, which are required for cell shape change, chemotaxis, phagocytosis, and granule translocation in leukocytes [9-14]. Importantly, Rac2 is critical for fMLP-induced enhancement of azurophil granule translocation in CB-treated neutrophils [15-17]. Recently, another small GTPase Rab27a was reported to regulate exocytosis of azurophil granules in neutrophils [18].
Lysophosphatidylcholine (LPC), an endogenous lipid, has various stimulating or modulating activities in neutrophils [19-26]. Previously, we showed that LPC increases bactericidal activity in human neutrophils by enhancement of azurophil granule-phagosome fusion via calcium/p38 MAPK dependent azurophil granule translocation [27]. However, the involvement of a small GTPase in the LPC-induced azurophil granule translocation has not yet been investigated. In the present study, we found that Rho, but not Rac2, is critical for LPC-induced enhancement azurophil granule translocation via Rho kinase (ROCK)/fibrous (F)-actin polymerization, elucidating a novel signaling pathway for azurophil granule translocation.
METHODS
Reagents
LPC (18:0) was obtained from Avanti Polar Lipids (Alabaster, AL, USA), and 3 mM stock solution in phosphate-buffered saline (PBS) was prepared with sonication. NSC23766, SB203580, and Y27632 were from Tocris Bioscience (Ellisville, MO, USA). Cytofix, Cytofix/Cytoperm, and FITC anti-CD63 Ab were from BD Bioscience (San Diego, CA, USA). FITC-conjugated phalloidin and CB were from Sigma-Aldrich Chemical (St. Louis, MO, USA). Fluo-3 AM was from Molecular Probes (San Diego, CA, USA). Rho activation assay kit was from Upstate (Billerica, MA, USA). Tat-C3 (a fusion protein between Tat-peptide and C3 toxin) was purified from bacteria containing the Tat-C3 cDNA plasmid as previously described [28]. Anti-G2A Ab (N-20) was from Santa Cruz Biotechnology (Santa Cruz, CA, USA).
Preparation of neutrophils
Neutrophils were purified from healthy volunteer donors. In brief, whole blood was layered onto a discontinuous Ficoll-Hypaque density gradient (Histopaque 1077; Sigma-Aldrich) and centrifuged at 2,500 rpm for 30 min. After dextran sedimentation, residual erythrocytes were eliminated with hypotonic lysis. The purity of neutrophils measured by Diff-Quick staining was > 95% on average. The viability of neutrophils stained with tryphan blue was > 99% as determined by morphology.
Flow cytometry
For the analysis of azurophil granule translocation, CD63, an azurophil granule marker, was used. Isolated neutrophils were stabilized for 30 min and stimulated with LPC (3 μM) for 15 min. Cells were fixed with Cytofix and washed. Then cells were stained with FITC-conjugated anti-CD63 Ab for 1 h on ice. For analysis of F-actin polymerization, neutrophils were fixed and permeabilized with Cytofix/Cytoperm for 30 min. After washing, neutrophils were stained with FITC-conjugated phalloidin, an F-actin marker, for 1 h on ice. Neutrophils were acquired on FACSCalibur (BD Bioscience) and were analyzed using Cell Quest Pro 6.0 (BD Bioscience) and FCS express V3 (De Novo Software, Los Angeles, CA, USA).
Pull-down assay
Neutrophils were resuspended in RPMI1640 and stimulated with LPC (3 μM) for 15 min. Cells were lysed in NP40 lysis buffer to detect Rho activity with Rho activation assay kit according to the manufacturer’s protocol. In brief, cells were lysed in NP-40 lysis buffer and the target proteins were loaded to 30 μl of the Rho-assay reagent slurry. The reaction mixtures were incubated for 45 min at 4°C with gentle agitation. Agarose beads were pelleted by brief centrifugation and 20 μl of the mixture per lane was loaded on an appropriate polyacrylamide gel. Membrane was incubated overnight at 4°C with agitation. The proteins were detected by enhanced chemiluminescence detection system.
Immunofluorescence microscopy
Neutrophils were transferred to 12-well plate and stimulated with LPC (3 μM). Then cells were fixed using 2% paraformaldehyde for 15 min and permeabilized using 0.05% Triton X-100 for 10 min at room temperature. Next, cells were blocked with 3% bovine serum albumin and incubated for 1 h. After incubation, neutrophils were washed three times and stained with FITC-conjugated phalloidin. After three washes in PBS, cells were mounted using mounting medium. Visual inspection and recording of images were performed using a Zeiss confocal microscope (Carl Zeiss, Thornwood, NY, USA).
Measurement of [Ca2+]i
[Ca2+]i was measured using Fluo-3 AM as previously described [25]. Neutrophils were loaded with Fluo-3 AM (4 μM) in 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES) physiologic salt solution (HEPES-PSS) (in mM) (HEPES 10, NaCl 140, KCl 5, CaCl2 1, D-glucose 10, MgCl2 1) for 1 h at 37°C. Neutrophils were washed twice with HEPES-PSS and resuspended in HEPES-PSS at 3 × 106 cells/ml. Traces of [Ca2+]i in Fluo-3 AM-loaded PMNs were measured with 490 nm/526 nm using Spectramax M2/e microplate reader (Molecular Device, Sunnyvale, CA, USA) every 10 sec. Raw fluorescence was subtracted with the average fluorescence during 5 min before the addition of LPC (3 μM). Cells were pretreated with Y27632 (10 μM) for 30 min before the addition of LPC.
Statistical analysis
All the statistical data were analyzed with Graphpad Prism 5.0 (Graphpad software). Data were expressed as mean ± SEM and statistical significance was evaluated by two-tailed Student’s t-test or ANOVA. Bonferroni test was used for post-hoc comparison. Value of p < 0.05 was considered to indicate statistical significance.
RESULTS
LPC induces azurophil granule translocation in human neutrophils
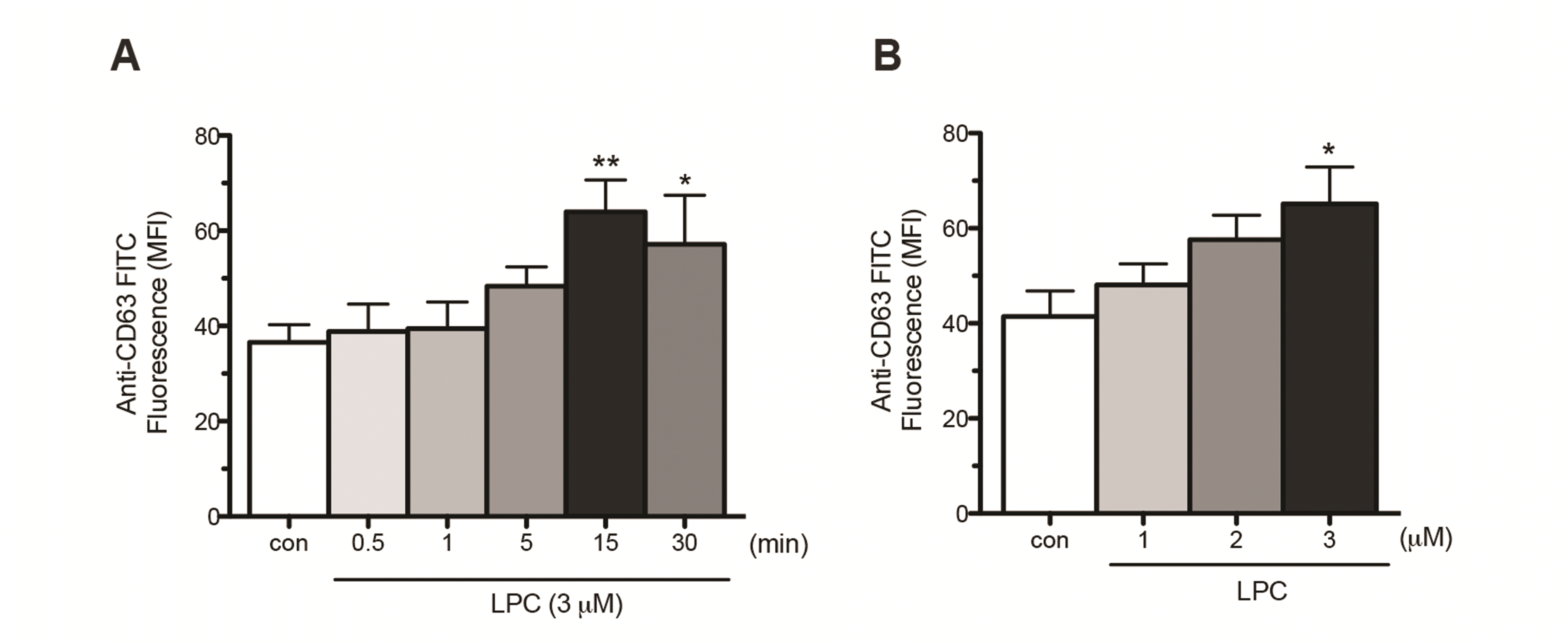
CD63 is a representative marker for azurophil granules, which translocates into the plasma membrane as azurophil granules degranulate [29]. Previously, we showed that LPC induces translocation of CD63 in human neutrophils in RPMI medium supplemented with 5% fetal bovine serum [27]. Although this condition mimics the in vivo condition, the various constituents of fetal bovine serum may affect LPC-induced signaling cascades. Thus, in this study we examined the effects of LPC in the serum-free condition. LPC increased surface expression of CD63 in a time- (Fig. 1A) and concentration- (Fig. 1B) dependent manner in serum-free condition; the LPC-induced CD63 surface expression peaked at 15 min and at 3 μM. This LPC-induced CD63 surface expression in the serum-free condition is comparable to that observed in serum-containing condition [27].
Contrarily, fMLP (1 μM) alone expectedly could not induce surface expression of CD63 (Fig. 2A). However, in the presence of CB (2.5 μg/ml, i.e., 5.2 μM) that by itself did not affect CD63 surface expression, fMLP markedly increased surface expression of CD63 (Fig. 2A, B). Intriguingly, however, CB pretreatment blocked LPC-induced CD63 surface expression (Fig. 2A, B). This result suggests that LPC enhances CD63 surface expression by induction of F-actin polymerization in human neutrophils. LPC increased F-actin polymerization as shown by increased staining of FITC-conjugated phalloidin, a marker of F-actin (Fig. 2C). These results indicate that LPC induces CD63 translocation via F-actin polymerization in human neutrophils. It is to be noted that while fMLP in the presence of CB increases mean fluorescence intensity (MFI) by a shift of the whole distribution to the right, LPC increases MFI with making the distribution more bimodal by increasing the proportion of cells with the highest values (Fig. 2A).
Involvement of Rho signaling in LPC-induced azurophil granule translocation
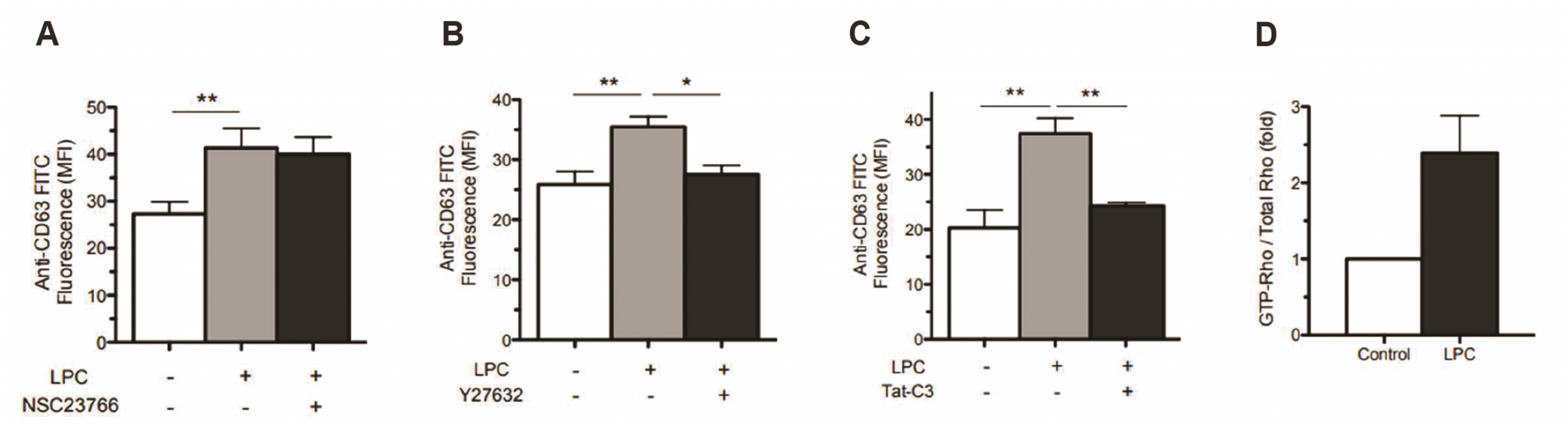
To examine whether Rac2 is involved in LPC-induced azurophil granule translocation, we investigated the effect of NSC23766, an inhibitor of Rac, on LPC-induced CD63 surface expression. Interestingly, NSC23766 (50 μM) did not affect LPC-induced translocation of azurophil granules (Fig. 3A), however, Y27632 (10 μM), a ROCK inhibitor, effectively blocked the LPC-induced CD63 surface expression (Fig. 3B). Tat-C3 exoenzyme, a specific inhibitor of Rho, also effectively inhibited LPC-induced CD63 surface expression (Fig. 3C). LPC (3 μM) markedly increased Rho-GTP activity in human neutrophils at 15 min (Fig. 3D). These results indicate that LPC induces CD63 surface expression via Rho/ROCK signaling.
To examine whether inhibition of ROCK blocks LPC-induced F-actin polymerization, neutrophils were pretreated with Y27632 (10 μM) before LPC treatment and stained with FITC-conjugated phalloidin. Y27632 blocked LPC-induced F-actin polymerization (Fig. 4A). Next, we performed confocal microscopy analysis for the inhibitory effect of Y27632 on LPC-induced F-actin polymerization. Resting neutrophils have cortical F-actin staining in a ring-like structure [30], as confirmed in Fig. 4B, left. Treatment with LPC (3 μM) for 15 min enhanced F-actin staining in the cytosol of neutrophils (Fig. 4B, middle). Remarkably, Y27632 blocked the LPC-induced enhancement of cytosolic actin polymerization (Fig. 4B, right). These results indicate that Rho/ROCK signaling is involved in LPC-induced F-actin polymerization, resulting in increased CD63 surface expression.
Two distinct signaling pathways in LPC-induced azurophil granule translocation
G2A, a G protein-coupled receptor abundantly expressed in immune cells, is reportedly involved in various actions of LPC in neutrophils [20,21,24,27]. To examine the involvement of G2A in LPC-induced CD63 surface expression, neutrophils were pretreated with anti-G2A Ab (1 μg/ml) for 1 h and further stimulated with LPC (3 μM) for 15 min. Anti-G2A Ab blocked the LPC-induced CD63 surface expression (Fig. 5A), suggesting that LPC induces azurophil granule translocation via G2A.
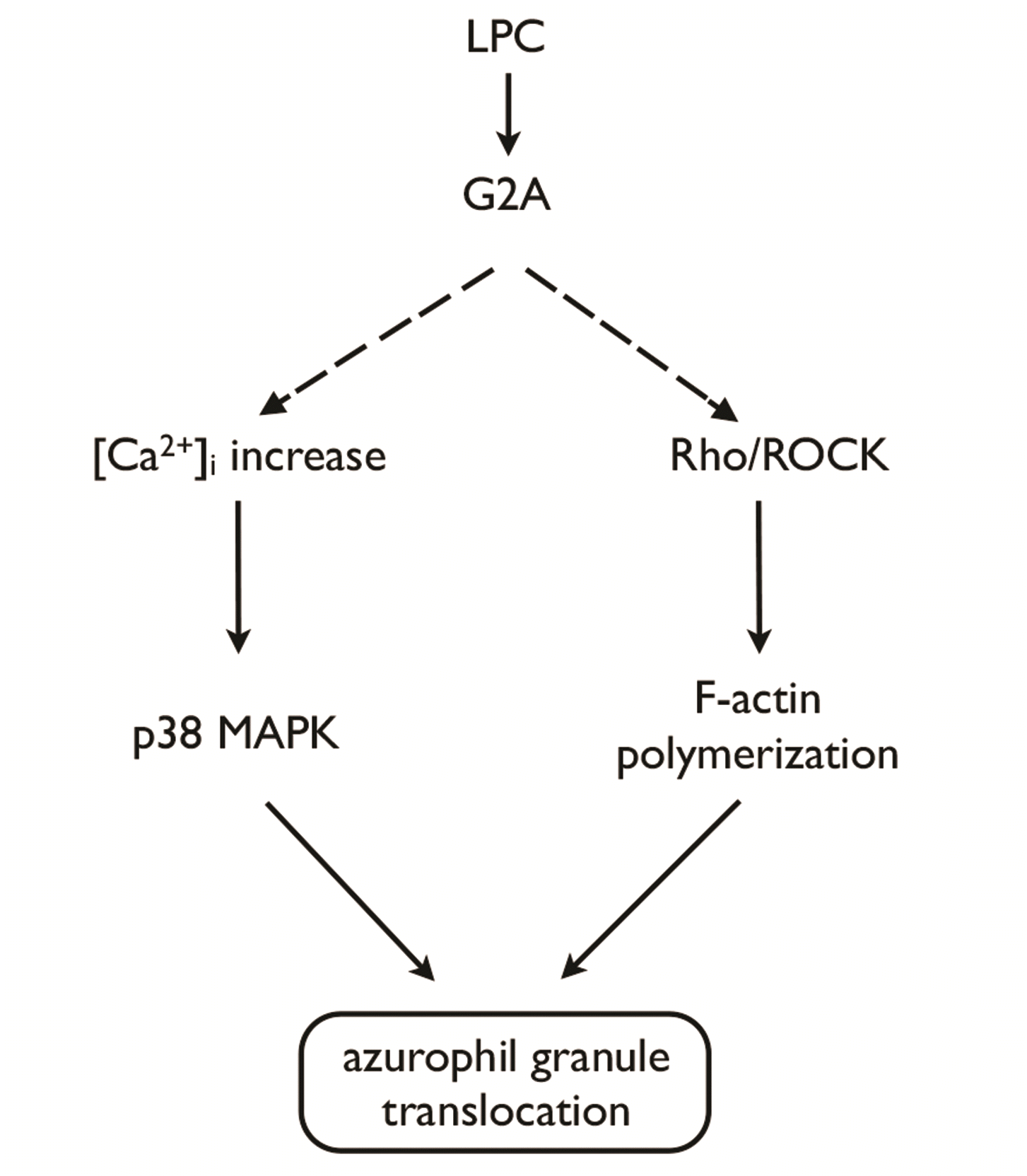
Previously we reported that Ca2+/p38 MAPK signaling pathway is involved in LPC-induced enhancement of azurophil granule-phagosome fusion in human neutrophils [27]. Similarly, to our previous observation in serum-containing condition [27], pretreatment with SB203580 (10 μM), an inhibitor of p38 MAPK, completely inhibited LPC-induced CD63 surface expression (Fig. 5B). However, it did not affect the LPC-induced increase in F-actin polymerization found in this study (Fig. 5C). Reciprocally, Y27632 (10 μM) did not affect LPC-induced [Ca2+]i increase (Fig. 5D). These results suggest that LPC induces azurophil granule translocation via 2 complementary, but distinct signaling pathways, i.e., Rho/ROCK/F-actin polymerization and Ca2+/p38 MAPK pathways (Fig. 6).
DISCUSSION
In this study, we found a novel mechanism for azurophil granule translocation in human neutrophils, wherein LPC induces azurophil granule translocation through G2A/Rho/ROCK/F-actin polymerization, complementing the previously identified LPC-induced [Ca2+]i increase/p38 MAPK activation pathway (Fig. 6). Interestingly, there were several differences between LPC and fMLP in their mode of action for increasing azurophil granule translocation. LPC alone (in the absence of CB) effectively increased azurophil granule translocation (Fig. 2A). In sharp contrast to the case of fMLP (Fig. 2A, B; [6,15]), CB effectively blocked LPC-induced azurophil granule translocation rather than enhancing it (Fig. 2A, B). LPC induced enhancement of F-actin polymerization in human neutrophils (Fig. 2C), which was cytosolic as shown in confocal microscopy (Fig. 4B). FMLP also reportedly increases azurophil granule translocation in the presence of CB via cytosolic F-actin polymerization in human neutrophils [30]. Importantly, Rac2 critically mediates F-actin polymerization for the translocation of azurophil granules to the plasma membrane in human neutrophils in response to stimulation with fMLP [30]. Intriguingly, in another sharp contrast to fMLP [15,17], LPC-induced azurophil granule translocation was critically dependent on Rho/ROCK, but not Rac2, signaling pathway (Fig. 3). To our knowledge, this is the first report to implicate the importance of Rho in azurophil granule translocation in human neutrophils.
G2A, activated by LPC, is linked to Gαs [22,31], Gαi [20,21], Gαq [21,32], and Gα13 [31-33]. For LPC-induced azurophil granule translocation, two distinct complementary pathways are essential (Fig. 6). One is the previously described calcium-dependent p38 MAPK pathway [27] and the other is Rho/ROCK/F-actin polymerization pathway identified in this study. These two pathways are dependent on G2A ([27]; Fig. 5A). Although we did not investigate the identity of specific heterotrimeric G-proteins involved in each pathway, two different heterotrimeric G- proteins could be involved downstream of G2A. For calcium-dependent p38 MAPK pathway, we previously showed that pertussis toxin, an inhibitor of Gαi, blocks LPC-induced increase in Cl– influx which subsequently leads to [Ca2+]i increase in human neutrophils [27], in line with the previous reports [20,23]. Further, both Gαi and Gαq was recently shown to be involved in LPC-induced increase in [Ca2+]i increase in human neutrophils [21]. On the other hand, because Gα13 was reported to be involved in G2A-mediated activation of RhoA-dependent actin remodeling in fibroblast cells [33], Gα13 could be involved in LPC-induced Rho/ROCK/F-actin polymerization.
Collectively, we found a novel signaling pathway for azurophil granule translocation in human neutrophils, i.e., G2A/RhoA/ROCK for LPC, in addition to the well-established Rac2 pathway for fMLP, IL-8 or leukotriene B4 [15], suggesting the presence of distinct small GTPase pathways for azurophil granule translocation depending on different stimulators. Further, the results of this study could contribute to better understanding of LPC-induced enhancement of neutrophil bactericidal activity [27] in relation to its anti-septic action [24].

 XML Download
XML Download