

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
The main pathobiology of diabetes involves the decreased number and dysfunction of islet cells [1,2]. Inflammatory damage mediated by cytokines affects insulin secretion and the survival of islet cells [3,4]. Abnormal activation of Nod-like receptor family pyrin domain containing 3 (NLRP3) induces cell pyroptosis [5].
Pyroptosis is a programmed cell death mediated by various injury stimuli via inflammasome complexes. As a receptor receiving injury stimuli, NLRP3 plays an important role. The NLRP3 inflammasome is composed of NLRP3, apoptosis-associated speck-like protein containing a CARD (ASC) and cysteine aspartic acid-specific proteinase-1 (caspase-1). Activated inflammasomes stimulate caspase-1, which cleaves interleukin (IL)-1 and IL-18 precursor molecules, releasing IL-1 and IL-18, which participate in the inflammatory response [5,6]. Gasdermin D (Gsdmd) protein, the direct executor of cell pyroptosis, is cleaved into Gsdmd-N and Gsdmd-C by activated caspases. Gsdmd-N has lipophilicity and pore-forming activity, which combines cell membrane lipids, induces cell membrane perforation, and releases inflammatory mediators [7].
Studies have found that high glucose, high fat, reactive oxygen species (ROS), and nuclear factor kappa B (NF-κB) can activate NLRP3 inflammasome and impair islet function [8-10]. ROS can damage islet β cells and inhibit insulin synthesis and secretion [11]. Nuclear factor (erythroid-derived 2)-like 2 protein (Nrf2) is a redox-sensitive transcription factor. Activated Nrf2 regulates downstream gene transcription to enhance the production of proteins, such as NAD (P)H: quinone oxidoreductase 1, glutamate-cysteine ligase, heme oxygenase-1 (HO-1), and superoxide dismutase, which reduce ROS production and ameliorate cell function disorders [12]. Nrf2 mainly mediates Keleh-like ECH-associated protein (Keap1)-Nrf2/antioxidant response element (ARE) signaling pathway. Under normal physiological conditions, Nrf2 is primarily negatively regulated by Keap1. Nrf2 bound to Keap1 remains in the cytoplasm, maintaining low transcriptional activity [13]. Under oxidative stress, Nrf2 is released from Keap1, translocates to the nucleus, and binds to the ARE to activate target gene transcription and produce antioxidant enzymes [14]. The number and activity of β cells have been found to be significantly lower in Nrf2 knockout mice than in wild-type mice [15]. Yagishita et al. [16] have also demonstrated that Nrf2 expression can inhibit ROS accumulation in pancreatic β cells.
Oxymatrine is the main component of a traditional Chinese herb, Sophora flavescens Ait. Recent studies have demonstrated that oxymatrine is helpful for the treatment of diabetes. In umbilical vein endothelial cells, oxymatrine alleviates high glucose-induced endothelial toxicity by inhibiting ROS production [17,18]. Oxymatrine can reduce the levels of advanced glycation end products, ROS, and inflammatory cytokines in the kidney of diabetic rats [19]. We had previously shown that oxymatrine stimulated insulin secretion in isolated rat islets, increased rat insulinoma (INS-1) cell vitality and cell proliferation, and inhibited apoptosis in INS-1 cells [20]. However, as the role of oxymatrine in pyroptosis in INS-1 cells is unclear, we investigated the effects of oxymatrine on pyroptosis in INS-1 cells as well as the NF-κB pathway, ROS production, and levels of Nrf2 and HO-1.
Go to : 
METHODS
Cell cultures
The INS-1 cell line was obtained from AiYan Biological technology Co., Ltd. (Shanghai, China). The cells were cultured in RPMI-1640 medium (Gibco, Grand Island, NY, USA) supplemented with streptomycin (100 μg/ml; Sigma-Aldrich, St. Louis, MO, USA), penicillin (100 U/ml; Sigma-Aldrich), β-mercaptoethanol (50 μM; Gibco), sodium pyruvate (0.11 g/L; Sangon Biotech Co., Ltd., Shanghai, China), and fetal bovine serum (10%; Gibco) at 37°C in a humidified atmosphere containing 5% CO2. The INS-1 cells were treated as follows: a control group: no treatment; HG + PA group: high glucose (30 mM glucose [Sangon Biotech Co., Ltd.]) + high fat (400 μM palmitic acid sodium [Sigma-Aldrich]); HG + PA + oxymatrine (1 μM) group: high glucose (30 mM glucose) + high fat (400 μM palmitic acid sodium) + oxymatrine (1 μM [Sigma-Aldrich]); HG + PA + oxymatrine (10 μM) group: high glucose (30 mM glucose) + high fat (400 μM palmitic acid sodium) + oxymatrine (10 μM). The concentrations of oxymatrine were based on the results of our previous work [20].
Flow cytometry analysis
The FAM-FLICA Caspase-1 Assay Kit (Immuno Chemistry Technologies, Bloomington, MN, USA) was employed for cell pyroptosis detection. The INS-1 cells were seeded in 6-well plates at a density of 4 × 105 cells/ml. After treatment for 24 h, the cells were collected, washed, and centrifuged (1,000 rpm, 5 min). The supernatant was discarded, and the pellets were mixed with FAM-FLICA caspase-1 (10 μl:290 μl) and incubated at 37°C for 1 h. After the medium was removed by centrifugation and the samples washed three times with 1 × Apoptosis wash buffer, a working solution of propidium iodide (100 μg/ml) was added to the cell suspension (1 μl:100 μl). The cells were incubated at room temperature for 15 min and detected using a Beckman DxFLEX flow cytometer (Beckman Coulter, Inc., Indianapolis, IN, USA).
Reactive oxygen species assay
A Reactive Oxygen Species Assay Kit (Beyotime Biotechnology, Shanghai, China) was used to measure ROS production. After treatment for 24 h, the INS-1 cells were digested with trypsin without EDTA (Gibco), centrifuged (1,500 rpm, 5 min) at 4°C, and collected. Each sample was incubated with 1.5 ml of a working solution of dichlorodihydrofluorescein diacetate (5 μM) at 37°C in the dark for 20 min. Subsequently, the cells were centrifuged (1,500 rpm, 10 min), and the staining solution was carefully discarded. After washing two times with phosphate-buffered saline (PBS), the cells were resuspended with PBS (200 μl/well). Beckman DxFLEX flow cytometer was used to detect the fluorescence signals of 10,000 cells within half an hour to obtain a curve.
Lactate dehydrogenase (LDH) release assay
The release of LDH in the supernatants was measured by using a commercially available LDH assay kit (Jiancheng Bioengineering Institute, Nanjing, China) according to the manufacturer’s instructions. The INS-1 cells were treated as follows: control; HG + PA; HG + PA + oxymatrine (1 μM); HG + PA + oxymatrine (10 μM). The cells were cultured in a 24-well plate for 24 h. The absorbance was determined using a multifunctional microplate reader (Berthold Technologies, Bad Wildbad, Germany) at 450 nm.
Transmission electron microscopy (TEM)
The INS-1 cells were treated as follows: control; HG + PA; HG + PA + oxymatrine (10 μM). After the cells were digested and centrifuged twice (1,000 rpm, 5 min, and 1,600 rpm, 5 min), the supernatant was removed and the cell masses collected. The samples were fixed (2.5% glutaraldehyde and 1% osmic acid), dehydrated (ethanol and acetone), embedded in epoxy resin, sectioned using a microtome to a thickness of 50 nm, and stained (uranium acetate and lead citrate). Finally, the specimens were observed and images acquired using a transmission electron microscope (TEM; Hitachi, Tokyo, Japan).
Enzyme-linked immunosorbent assay (ELISA) to measure IL-1β and IL-18 secretion
IL-1β and IL-18 concentrations were measured using an IL-1β ELISA kit (R&D Systems, Minneapolis, MN, USA) and an IL-18 ELISA kit (Huijia Biological Technology Co., Ltd., Xiamen, China), respectively. The INS-1 cells were treated as follows: control; HG + PA; HG + PA + oxymatrine (10 μM). The cells were seeded in 96-well plates (5 × 103 cells/well) for 24 h. After the cells were digested and centrifuged (2,000 rpm, 20 min), the supernatant was collected, and the assay performed according to the manufacturer’s instructions. The optical density was determined at 450 nm by using the multifunctional microplate reader.
Measurement of caspase-1 activity
Caspase-1 activity was assessed by using a caspase-1 activity assay kit (Beyotime Biotechnology) according to the manufacturer’s instructions. The INS-1 cells were treated as follows: control; HG + PA; HG + PA + oxymatrine (10 μM). The cells were incubated in 96-well plates (5 × 103 cells/well) for 24 h. After the cells were digested and centrifuged (700 rpm, 5 min, 4°C), the supernatant was discarded. Subsequently, the cells were resuspended with pyrolysis buffer (100 μl pyrolysis buffer for 2 × 106 cells), lysed in an ice bath for 15 min, and centrifuged (3,800 rpm, 10 min, 4°C). The supernatant was mixed with precooled Ac-YVAD-pNA (2 mM) and incubated at 37°C for 60 min. The absorbance was measured at a wavelength of 405 nm using the multifunctional microplate reader.
Western blotting
The protein levels were analyzed by western blotting. The INS-1 cells were treated as follows: control; HG + PA; HG + PA + oxymatrine (1 μM); HG + PA + oxymatrine (10 μM). The cells were cultured in 6-well plates (4 × 105 cells/ml) for 24 h. In brief, the cell lysates were electrophoresed and transferred onto polyvinylidene fluoride membranes. Then, the membranes were incubated with the following primary antibodies (diluted in Tris-buffered saline with 0.1% Tween-20 [TBST] buffer): anti-NLRP3 antibody (ab214185, 1:1,000), anti-IL-1β antibody (ab205924, 1:1,000), anti-NF-κB p65 antibody (ab16502, 1:2,000), anti-Nrf2 antibody (ab89443, 1:500), anti-HO-1 antibody (ab13243, 1:2,000), anti-Keap1 antibody (ab119403, 1:1,000), anti-ASC antibody (ab180799, 1:2,000), and β-actin antibody (ab8226, 1:1,000) from Abcam (Cambridge, UK); anti-Gsdmd antibody (93709, 1:1,000) and anti-Histone H3 antibody (4499, 1:2,000) from Cell Signaling Technology (Boston, MA, USA); and anti-caspase-1 antibody (NBP1-45433, 1:1,000) from Novus Biologicals (Littleton, CO, USA). After washing, the membranes were incubated with the appropriate secondary antibody (diluted in TBST buffer) from Abcam (1:5,000, Cambridge, UK). Image Pro Plus 6.0 software (Media Cybernetics, Houston, TX, USA) was used to determine the intensity of the protein bands.
Luciferase reporter gene assay for DNA-binding activity of NF-κB and Nrf2
A firefly luciferase reporter gene assay kit was obtained from Beyotime Biotechnology. The NF-κB-luc reporter plasmid and Nrf2-luc reporter plasmid were purchased from Genomeditech (Shanghai, China). The INS-1 cells were treated as follows: control; HG + PA; HG + PA + oxymatrine (10 μM). After 36‒48 h of co-transfection with the plasmids, the cells in each well were treated with 100 μl of pyrolysis buffer and centrifuged (3,700 rpm, 3 min). Then, the supernatant was collected and mixed with luciferase test reagent (100 μl sample:100 μl luciferase test reagent) to measure the relative luciferase activity using the multifunctional microplate reader. The measurement time was 10 sec and the interval was 2 sec.
Statistical analysis
The data were expressed as mean ± standard deviation (SD). Sigmaplot (Systat Software, San Jose, CA, USA) was used to statistical analysis. Statistical differences were tested by one-way analysis of variance (ANOVA) and Tukey’s test. p < 0.05 was considered significant.
Go to :
RESULTS
Oxymatrine protected INS-1 cells from pyroptosis
Flow cytometry, the LDH release assay, and TEM were employed to determine the effects of oxymatrine on pyroptosis in INS-1 cells. As shown in Fig. 1, mitochondrial cristae and abundant organelles were visible in the control cells; no mitochondrial swelling or vacuolar degeneration was observed. The number of autophagosomal vesicles was higher in the HG + PA cells, which also showed mitochondrial swelling, vacuolar degeneration, irregular nuclear morphology, local nuclear membrane depression, decreased organelles, and a large number of intracellular vacuoles (Fig. 1A). The HG + PA cells showed an increase in caspase-1 activity to 18.70% ± 1.48% at 24 h (p < 0.01 vs. control, Fig. 1B, C) and the LDH content (p < 0.01 vs. control, Fig. 1D). Incubation of the HG + PA cells with oxymatrine (10 μM) decreased the number of autophagosomal vesicles in the cytoplasm (Fig. 1A) and caspase-1 activity (11.00% ± 1.08% at 24 h; p < 0.01 vs. HG + PA, Fig. 1B, C). Oxymatrine (1 μM and 10 μM) also suppressed LDH release (p < 0.01 vs. HG + PA, Fig. 1D).
 | Fig. 1Oxymatrine protected INS-1 cells from pyroptosis.(A) The electron microscopic analysis of different cells. Scale bar: 2 μm; Oxymatrine: 10 μM. (B) Representative graphics from flow cytometry analysis were showed; Oxymatrine: 10 μM. (C) The caspase-1 activity analysis of different cells; Oxymatrine: 10 μM. (D) The LDH release analysis of different cells. control: no treatment; HG: high glucose (30 mM glucose); PA: palmitic acid sodium (400 μM). Arrows: the autophagosomal vesicles. Data are presented as mean ± SD and represent an average of three experiments. LDH, lactate dehydrogenase. **p < 0.01 vs. control, ##p < 0.01 vs. HG + PA; the experiments were repeated three times.
|
Oxymatrine decreased the levels of NLRP3, Gsdmd-N, caspase-1, ASC, IL-1β, and IL-18 in INS-1 cells under high glucose and high fat conditions
After the cells were treated under different conditions, the levels of NLRP3, Gsdmd-N, caspase-1, ASC, and IL-1β were detected by Western blotting, while IL-1β and IL-18 secretion was measured using ELISA. Caspase-1 activity was examined by spectrophotometry. The levels of NLRP3, Gsdmd-N, caspase-1, IL-1β, and ASC (p < 0.01 vs. control, Fig. 2A‒G), the secretion of IL-1β and IL-18 (p < 0.01 vs. control, Fig. 2H), and caspase-1 activity (p < 0.01 vs. control, Fig. 2I) were increased in HG + PA cells. As expected, treatment with oxymatrine decreased these protein levels (p < 0.01 vs. HG + PA, Fig. 2B–E; p < 0.05 vs. HG + PA, Fig. 2D, G), the secretion of IL-1β and IL-18 (p < 0.01 vs. HG + PA, Fig. 2H), and caspase-1 activity (p < 0.01 vs. HG + PA, Fig. 2I).
 | Fig. 2Oxymatrine decreased the levels of NLRP3, Gsdmd-N, caspase-1, ASC, IL-1β, and IL-18 in INS-1 cells under high glucose and high fat conditions.(A) Representative Western blotting bands of NLRP3, Gsdmd-N, caspase-1, and IL-1β proteins were showed. (B) Analysis of the amount of NLRP3 proteins. (C) Analysis of the amount of caspase-1 proteins. (D) Analysis of the amount of IL-1β proteins. (E) Analysis of the amount of Gsdmd-N proteins. (F) Representative western blotting bands of ASC proteins were showed; Oxymatrine: 10 μM. (G) Analysis of the amount of ASC proteins; Oxymatrine: 10 μM. (H) Analysis of the secretion of IL-1β and IL-18; Oxymatrine: 10 μM. (I) Analysis of the activity of caspase-1; Oxymatrine: 10 μM. control: no treatment; HG: high glucose (30 mM glucose); PA: palmitic acid sodium (400 μM). Data are presented as mean ± SD and represent an average of three experiments. NLRP3, Nod-like receptor family pyrin domain containing 3; Gsdmd, Gasdermin D; caspase-1, cysteine aspartic acid-specific proteinase-1; ASC, apoptosis-associated speck-like protein containing a CARD; IL, interleukin. Data from Western blotting are normalized to control. **p < 0.01 vs. control, #p < 0.05, ##p < 0.01 vs. HG + PA; the experiments were repeated three times.
|
Oxymatrine inhibited high glucose and high fat-induced ROS production in INS-1 cells
As indicated in Fig. 3, ROS production was higher in HG + PA cells than in the control cells (p < 0.01 vs. control). Treatment with oxymatrine suppressed ROS production in HG + PA cells (p < 0.05 vs. HG + PA, Fig. 3A, B).
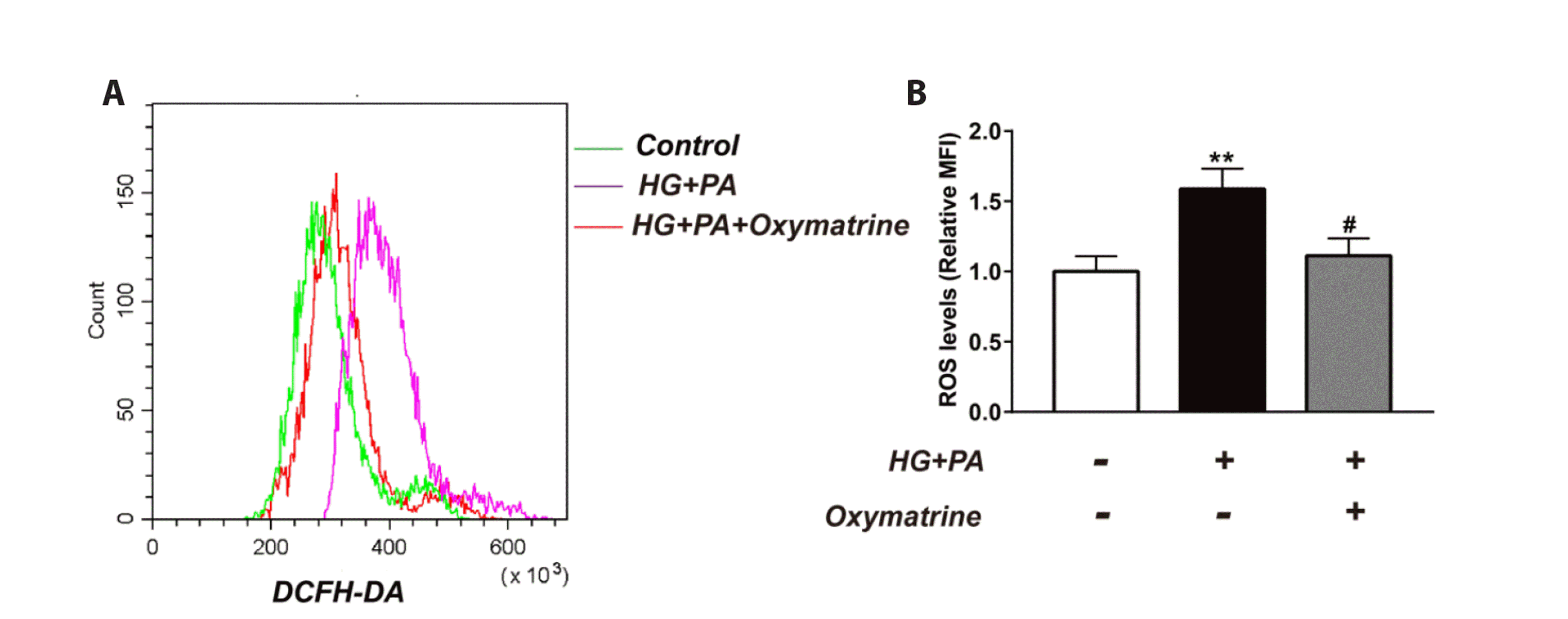 | Fig. 3Oxymatrine inhibited high glucose and high fat-induced ROS production in INS-1 cells.(A) Representative graphics of ROS production were showed. (B) The ROS production from different cells. control: no treatment; HG: high glucose (30 mM glucose); PA: palmitic acid sodium (400 μM); Oxymatrine: 10 μM. Data are presented as mean ± SD and represent an average of three experiments. ROS, reactive oxygen species; DCFH-DA, Dichloro-dihydro-fluorescein diacetate; MFI, mean fluorescence intensity. Data are normalized to control. **p < 0.01 vs. control, #p < 0.05 vs. HG + PA; the experiments were repeated three times.
|
Oxymatrine affected NF-κB (p65) levels in INS-1 cells at different times
Treatment of INS-1 cells under different conditions for 0.5 h, 1 h, or 2 h altered NF-κB (p65) protein levels. In HG + PA cells, the nuclear p65 protein levels increased over time (0.5 h: 2.32 ± 0.22; 1 h: 3.64 ± 0.32; 2 h: 3.99 ± 0.35) (p < 0.01 vs. control, Fig. 4A, B, D, F) but decreased after treatment with oxymatrine (0.5 h: 1.17 ± 0.14; 1 h: 1.41 ± 0.14; 2 h: 1.82 ± 0.12) (p < 0.01 vs. HG + PA, Fig. 4A, B, D, F). Compared with the control cells, cytoplasmic p65 protein levels of HG + PA cells decreased over time (0.5 h: 0.54 ± 0.05; 1 h: 0.40 ± 0.05; 2 h: 0.34 ± 0.03) (p < 0.01 vs. control, Fig. 4A, C, E, G) but increased after treatment with oxymatrine (0.5 h: 0.91 ± 0.07; 1 h: 0.77 ± 0.08; 2 h: 0.66 ± 0.06) (p < 0.01 vs. HG + PA, Fig. 4A, C, E, G).
 | Fig. 4Oxymatrine affected NF-κB (p65) levels in INS-1 cells at different times.(A) Representative Western blotting bands of p65 proteins in the nucleus or cytoplasm were showed. (B) Analysis of the amount of p65 proteins in the nucleus when the cells were incubated for 0.5 h. (C) Analysis of the amount of p65 proteins in the cytoplasm when the cells were incubated for 0.5 h. (D) Analysis of the amount of p65 proteins in the nucleus when the cells were incubated for 1 h. (E) Analysis of the amount of p65 proteins in the cytoplasm when the cells were incubated for 1 h. (F) Analysis of the amount of p65 proteins in the nucleus when the cells were incubated for 2 h. (G) Analysis of the amount of p65 proteins in the cytoplasm when the cells were incubated for 2 h. Control: no treatment; HG: high glucose (30 mM glucose); PA: palmitic acid sodium (400 μM); Oxymatrine: 10 μM. Data are presented as mean ± SD and represent an average of three experiments. NF-κB, nuclear factor kappa B. Data are normalized to control. **p < 0.01 vs. control, ##p < 0.01 vs. HG + PA; the experiments were repeated three times.
|
Oxymatrine affected Nrf2, HO-1, and Keap1 levels in INS-1 cells
As shown in Fig. 5, the levels of HO-1 and nuclear Nrf2 proteins increased in HG + PA cells compared with those in the control cells (p < 0.01 vs. control, Fig. 5A, B; p < 0.05 vs. control, Fig. 5A, E). In contrast, levels of Keap1 and cytoplasmic Nrf2 proteins decreased in HG + PA cells (p < 0.01 vs. control, Fig. 5A, C, D). The treatment of HG + PA cells with oxymatrine increased nuclear Nrf2 and HO-1 levels (p < 0.01 vs. HG + PA, Fig. 5A, B, E) and decreased cytoplasmic Nrf2 and Keap1 levels (p < 0.05 vs. HG + PA, Fig. 5 A, C, D) to a greater extent.
 | Fig. 5Oxymatrine affected Nrf2, HO-1, and Keap1 levels in INS-1 cells.(A) Representative Western blotting bands of Nrf2, HO-1, and Keap1 proteins in the nucleus or cytoplasm were showed. (B) Analysis of the amount of Nrf2 proteins in the nucleus. (C) Analysis of the amount of Nrf2 proteins in the cytoplasm. (D) Analysis of the amount of Keap1 proteins. (E) Analysis of the amount of HO-1 proteins. Control: no treatment; HG: high glucose (30 mM glucose); PA: palmitic acid sodium (400 μM); Oxymatrine: 10 μM. Data are presented as mean ± SD and represent an average of three experiments. Nrf2, nuclear factor (erythroid-derived 2)-like 2 protein; HO-1, heme oxygenase-1; Keap1, Keleh-like ECH-associated protein. Data are normalized to control. *p < 0.05 vs. control, **p < 0.01 vs. control, #p < 0.05 vs. HG + PA. ##p < 0.01 vs. HG + PA; the experiments were repeated three times.
|
Oxymatrine affected DNA-binding activity of NF-κB (p65) and Nrf2 in INS-1 cells
The DNA-binding activity of NF-κB (p65) and Nrf2 were assessed in INS-1 cells after 24 h treatment under different conditions. Compared with the control cells, the transcriptional activity of p65 and Nrf2 was activated in HG + PA cells (p < 0.01 vs. control, Fig. 6A, B). Oxymatrine treatment inhibited the transcriptional activity of p65 (p < 0.05 vs. HG + PA, Fig. 6A) but enhanced that of Nrf2 (p < 0.01 vs. HG + PA, Fig. 6B).
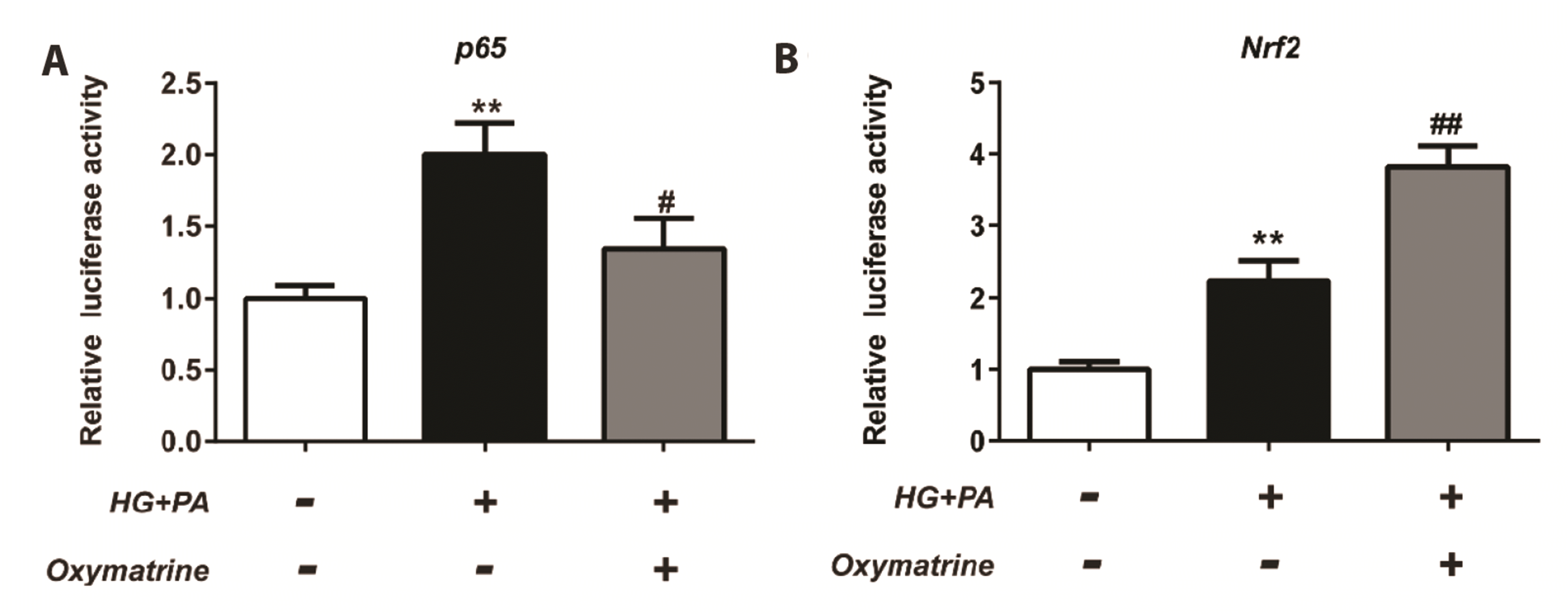 | Fig. 6Oxymatrine affected DNA-binding activity of NF-κB (p65) and Nrf2 in INS-1 cells.(A) Analysis of the DNA-binding activity of NF-κB (p65). (B) Analysis of the DNA-binding activity of Nrf2. Control: no treatment; HG: high glucose (30 mM glucose); PA: palmitic acid sodium (400 μM); Oxymatrine: 10 μM . Data are presented as mean ± SD and represent an average of three experiments. NF-κB, nuclear factor kappa B; Nrf2, nuclear factor (erythroid-derived 2)-like 2 protein. Data are normalized to control. **p < 0.01 vs. control, #p < 0.05, ## p < 0.01 vs. HG + PA; the experiments were repeated three times.
|
Go to :
DISCUSSION
Previous reports have indicated that high glucose and high fat concentrations can induce cell pyroptosis. Li et al. [21] have shown that high glucose-treated hippocampal neuronal cells undergo pyroptosis in vitro in an NLRP3-dependent manner. Gu et al. [22] demonstrated that high glucose levels trigger pyroptosis in human renal glomerular endothelial cells. In high fat diet-fed obese mice, hypertrophic adipocytes have been found to undergo pyroptosis [23]. In the mouse pre-osteoblast MC3T3-E1 cell line, high glucose inhibits the proliferation of osteoblasts by activating the pyroptosis pathway [24]. Pyroptosis is also implicated in the high glucose-induced cell death of H9c2 cardiomyocytes, human ventricular cardiomyocytes, and EA.hy926 endothelial cells [25-27]. Moreover, pyroptosis is closely related to diabetes. Type 2 diabetic db/db mice showed high levels of NLRP3 and IL-1β [28]. However, after the application of an NLRP3 inhibitor, these type 2 diabetic mice showed a decrease in NLRP3 and IL-1 β levels along with an amelioration of insulin resistance [28]. A 2013 report also confirms that type 2 diabetes mellitus (T2DM) is associated with the NLRP3-related inflammatory response, which is induced by activating the mitochondrial ROS pathway [29]. In T2DM rats, NLRP3 is activated by the NF-κB and thioredoxin interacting protein, which affects the progression of diabetic cardiomyopathy [25]. Pyroptosis is also commonly observed in various complications of T2DM, such as diabetic nephropathy and diabetic retinopathy [30,31]. Consistent with these findings, the present study data showed an increase in the number of autophagosomal vesicles in the cytoplasm of HG + PA INS-1 cells. High glucose and high fat also increased LDH release, caspase-1 activity, and the levels of related inflammatory factors, including NLRP3, IL-1β, IL-18, Gsdmd-N, caspase-1, and ASC, leading to pyroptosis in INS-1 cells. However, these changes were suppressed by oxymatrine treatment, demonstrating that oxymatrine can inhibit high glucose and high fat-induced pyroptotic cell death in INS-1 cells.
Hyperglycemia reduces the binding affinity of the NF-κB (p65) subunit to IκB alpha, causing increased nuclear translocation of p65 [32] and transcription of target genes involved in inflammatory responses [33]. Saturated fatty acids may also directly activate the toll-like-4 receptor, resulting in the activation of the downstream c-Jun NH 2-terminal kinase and inhibitor kappa B kinase β/NF-κB cascade [34,35]. Lipotoxicity, glucotoxicity, and glucolipotoxicity induce metabolic stress, which manifests as increased oxidative stress and ROS production [36-38]. As mentioned above, the activation of NLRP3 is affected by the NF-κB (p65) pathway and ROS production. Under oxidative stress, the transcriptional activity of Nrf2 was increased, and Nrf2 reduced ROS production by mediating the transcription and expression of a series of antioxidant stress proteins such as HO-1. Thus, both NF-κB and Nrf2 regulate NLRP3 by different mechanisms. Oxymatrine has been reported to regulate NF-κB pathway and Nrf2 expression. The NF-κB pathway is suppressed by oxymatrine in colon cancer cells and fibroblast-like synoviocytes [39,40]. By increasing the levels of Nrf2 and HO-1, oxymatrine reduces renal ischemia-reperfusion injury, cerebral ischemia-reperfusion injury, arsenic trioxide (As2O3)-affected liver injury, and lipopolysaccharide/D-galactosamine-induced acute liver failure [41-44]. Therefore, to explore the mechanism of oxymatrine-inhibited pyroptosis in INS-1 cells, we analyzed the effects of oxymatrine on these factors. The results showed that high glucose and high fat increased the DNA-binding activity of NF-κB (p65) and Nrf2. Treatment with oxymatrine suppressed the transcriptional activity of NF-κB and reduced the entry of P65 into the nucleus. In contrast, oxymatrine promoted Nrf2 transcription, which increased HO-1 expression and reduced intracellular ROS levels.
In summary, oxymatrine inhibits NLRP3-mediated pyroptosis in INS-1 cells, which may be related to its inhibition of the NF-κB pathway and activation of the Nrf2 pathway. These results contribute to understanding the mechanism of oxymatrine-regulated islet cell function and provide a theoretical basis for the clinical application of oxymatrine for the treatment of diabetes.
Go to :
 XML Download
XML Download