

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Disulfiram (DSF) has been used as a major therapeutic agent of alcohol dependence by inhibiting aldehyde dehydrogenase (ALDH) [1,2]. In the liver, alcohol is metabolized to acetaldehyde and can then be converted to acetic acid by ALDH. DSF blocks the activity of ALDH, resulting in the accumulation of acetaldehyde [3]. This causes serious hangover symptoms such as flushing of the skin, accelerated heart rate, shortness of breath, nausea, vomiting, and throbbing headache. DSF is a potent anti-cancer agent against various cancers [4]. DSF blocks the activity of ALDH, a marker of cancer stem cells, and inhibits proteasome activity by forming complexes with metal ions [5].
Most anti-cancer agents generate serious side effects including vomiting, hair loss, myelosuppression, and hypertension. In particular, myelosuppression is a life-threatening side effect in patients treated with anti-cancer agents [6], as bone marrow cells (BMs) provide hematopoietic and immune cells. Although there are many studies regarding the anti-cancer effects of DSF, useful information on the effects of DSF on BMs is lacking.
In this study, we determined whether DSF may influence the viability and function of BMs and the underlying mechanisms. To determine the toxicity of DSF in BMs, we measured the metabolic activity, mitochondrial function, and subset ratio of DSF-treated BMs. We investigated the effects of DSF on BMs treated with granulocyte-macrophage colony-stimulating factor (GM-CSF), an essential growth and differentiation factor for BMs. In addition, the BM toxicity of the DSF dosage used for anti-cancer effect was evaluated in vivo, based on the cellularity and subset analyses of BMs.
METHODS
Animals and reagents
C57BL/6 mice were purchased from ORIENT BIO (Seongnam, Korea) and maintained in our animal facility. 8- to 12-week-old mice were used in this study. For in vivo experiments, 10-week-old Balb/c mice were used. The weight range of mice was 22–26 g. All animal experiments were performed in accordance with the Institutional Guideline for Animal Use and Care of Jeju National University (approval No. 2019-0002, 2019-0034). DSF was dissolved in dimethyl sulfoxide to appropriate concentrations. Lipopolysaccharide (LPS) purified from Escherichia coli O55 was purchased from Sigma (St. Louis, MO, USA) and dissolved in sterile phosphate buffered saline.
Preparation of BMs
BMs were prepared from femur and tibia of mice by flushing as established in our lab [7]. BMs were treated with Ammonium-Chloride-Potassium Lysing Buffer (Thermo Fisher Scientific, Waltham, MA, USA) to remove red blood cells. The cells were then passed through a 70 µm cell strainer to obtain single cells. To culture BMs, 5% complete medium (RPMI 1640 medium containing 5% fetal bovine serum and 100 IU/ml penicillin/streptomycin, 2 mM L-glutamine) was used.
Measurement of cellular metabolic activity
For measurement of BM metabolic activity, BMs were cultured in 96-well culture plates at a concentration of 1 × 106 cells/ml (200 µl/well) and treated with DSF. After 3 days of culture, Cell Counting Kit-8 (CCK-8; Dojindo Molecular Technologies, Kumamoto, Japan) solution was added 10 µl/well for 4 h. The viable cells generate orange-colored products in proportion to their metabolic activity. The optical density of samples was measured at 450 nm using a microplate reader (Multiskan FC; Thermo Fisher Scientific) [8].
Flow cytometry analysis
BMs were cultivated at a concentration of 1 × 106 cells/ml in 6-well culture plates and treated with DSF for 3 days. The treated BMs were harvested and used for flow cytometry analysis. To measure apoptosis, the cells were stained with annexin V-fluorescein isothiocyanate (FITC) and 0.25 µg/ml propidium iodide (PI). To check mitochondrial membrane potential (MMP) in the BMs, the cells were incubated with 10 µg/mL rhodamine 123 (Sigma) for 30 min at room temperature. Additionally, to detect the proportion of granulocytes and B cells, allophycocyanin-labeled anti-Gr-1 antibody and biotin-labeled anti-B220 antibody, streptavidin-FITC were used. To analyze dendritic cell (DC)-specific marker expression, the cells were stained with FITC-labeled anti-MHC II antibody, phycoerythrin-labeled anti-CD11c antibody. All stained cells were analyzed with CytoFLEX flow cytometer and CytExpert software, or BD LSRFortessa Cell Analyzer and FlowJo software (all from BD Biosciences, San Diego, CA, USA).
In vivo BM toxicity of DSF
DSF was injected at a dosage of 50 mg/kg by intraperitoneal injection three times every other day (day 0, day 2, day 4). To measure the effects of DSF in vivo, the mice were divided into four groups including control. Mice were sacrificed on day 3, day 5, day 7. At indicated days, BMs were harvested from the femur and tibia of the mice. The cell number was counted by a trypan blue exclusion test and marker analysis was performed.
Statistical analysis
Data in graphs were presented as mean ± standard deviation (SD). Flow cytometry data were obtained from more than 3 independent experiments. Statistical significance was analyzed by Student’s t-test or one-way ANOVA, followed by Tukey-Kramer multiple comparison test using GraphPad Prism (GraphPad Software, San Diego, CA, USA). A p-value of < 0.05 was considered as significant. *, **, *** indicate p < 0.05, 0.01, 0.001 compared to the control, respectively.
RESULTS
Effects of DSF on the metabolic activity of BMs
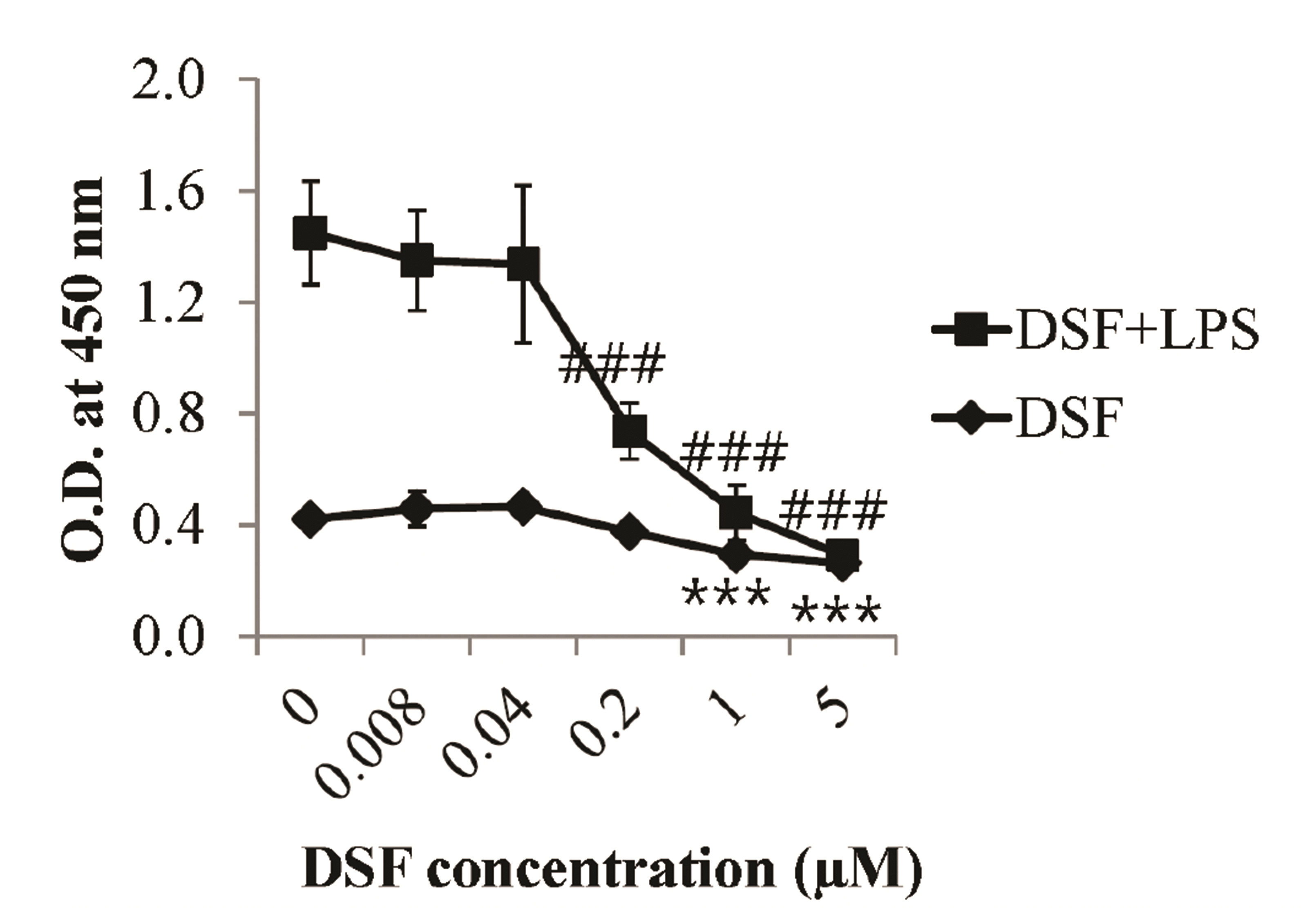
To assess the effects of DSF on BMs, they were treated in the absence or presence of 1 µg/ml lipopolysaccharide (LPS, a representative inflammatory agent) and DSF over a range of concentrations (0–5 µM). The CCK-8 assay demonstrated that LPS significantly increased the metabolic activity of BMs compared to control BMs (Fig. 1). DSF significantly decreased the cellular activity of BMs in the absence or presence of LPS. DSF suppressed basal and LPS-induced metabolic activity in BMs.
DSF decreases MMP of BMs and causes the cell death
To investigate how DSF affects the metabolic activity of BMs, we measured MMP of the cells. DSF-treated BMs were stained with rhodamine 123 solution. DSF significantly decreased the MMP of BMs at a range of DSF concentrations (0.04–5 µM) (Fig. 2B), indicating that DSF can destabilize the double-membrane structure of the mitochondria. Destabilization of the mitochondrial membrane in cells is closely correlated with cell death [9]. To determine whether DSF induces the death of BMs, the cells were stained with annexin V-FITC and PI. This quantitative cell death analysis revealed that DSF significantly increased the numbers of late apoptotic cells (annexin V+/PI+) and necrotic cells (annexin V–/PI+) compared to the control at DSF concentration of 1 and 5 µM (Fig. 2D).
DSF does not affect the subsets of BMs
To investigate how DSF influences the population of BMs, we measured the expression of subset-specific markers, B220 and Gr-1, on DSF-treated BMs (Fig. 3). Flow cytometry analysis revealed that DSF did not significantly affect the percentage of Gr-1- or B220-positive cells. These results indicate that DSF does not damage the ratio of BM subsets.
DSF impairs the metabolic activity and differentiation of GM-CSF-treated BMs
GM-CSF is an essential growth and differentiation factor for BMs [10]. To investigate whether DSF affects BMs, we measured the metabolic activity and differentiation of BMs after GM-CSF and DSF treatment. The metabolic activity of GM-CSF-treated BMs was not affected by DSF at lower concentrations (0–0.06 µM), whereas it was significantly affected at higher concentrations (0.125–0.5 µM) (Fig. 4A). In addition, 1 and 5 µM DSF blocked the differentiation of GM-CSF-treated BMs to CD11c+ DCs (Fig. 4B). These results demonstrated that DSF markedly impaired the metabolic activity and differentiation of GM-CSF-treated BMs.
The myelosuppressive effects of DSF in mice are minimal
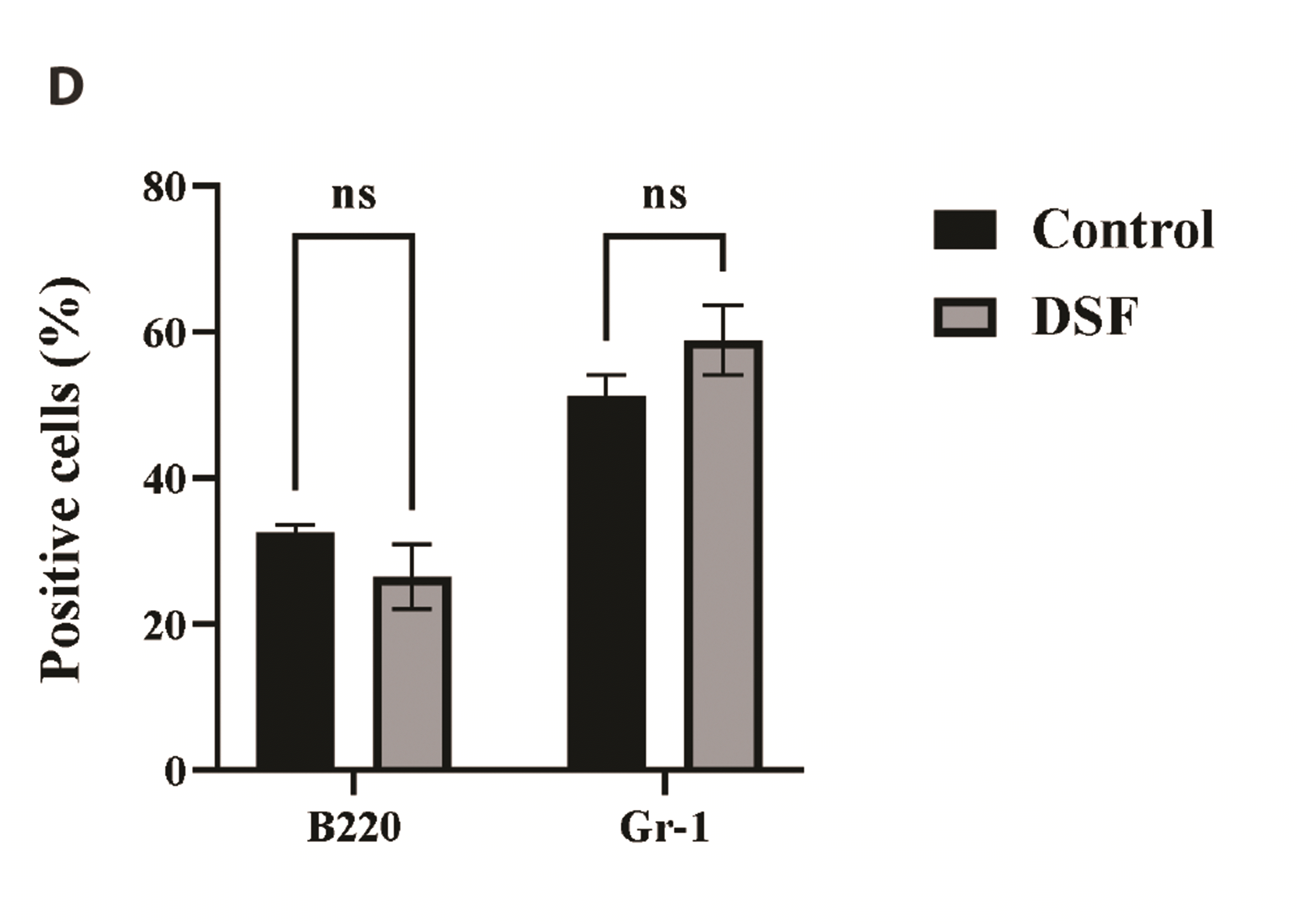
To evaluate the effects of DSF on the BMs in vivo, we injected mice with DSF (Fig. 5A). In this study, the mice were intraperitoneally injected with 50 mg/kg DSF, as described in the Methods. To measure the myelosuppressive effects of DSF, we counted the number of BMs. DSF marginally decreased the number of BMs in mice compared to control, but without statistical significance. A graph comparing the control and DSF (day 5) group is presented in Fig. 5B, which showed that DSF at the dosage does not significantly suppress the bone marrow in vivo. The major subsets of BMs were analyzed using surface markers, Gr-1 and B220 (Fig. 5C, D). The marker analysis revealed that DSF does not significantly change the proportion of granulocytes and B lymphocytes in BMs.
DISCUSSION
DSF has been used for the treatment of alcohol dependence by blocking ALDH [2]. Recent studies have demonstrated anti-cancer effects of DSF on solid and hematological malignancies. In breast cancer cells, DSF inhibits ALDH activity and modulate intracellular reactive oxygen species (ROS) generation [11]. In addition, DSF was identified as a novel cancer selective growth inhibitory compound for prostate cancer cells via high-throughput cell-based screening [12]. However, there have been few studies on the effects of DSF on BMs, although the mechanisms underlying the effects of DSF on cancer cells have been investigated. In this study, we demonstrated that DSF suppressed BMs, especially granulocytes, in vitro.
The metabolic activity of DSF-treated BMs was measured using the CCK-8 assay (Fig. 1). In vitro, DSF inhibits the metabolic activity of BMs. To assess the effects of DSF on inflammation, we stimulated the activity of BMs using LPS. DSF dramatically reduces the metabolic activity of BMs activated by LPS. While there was no significant change at low concentrations of DSF (0–0.04 µM), the metabolic activity of BMs was significantly decreased at DSF concentrations of 0.2–5 µM in the presence of LPS and 1–5 µM of DSF in the absence of LPS (Fig. 1). In addition, the MMP and death of DSF-treated BMs were investigated by flow cytometry. Similar to the results of the CCK-8 assay, the MMP of DSF-treated BMs was also decreased at DSF concentrations of 0.04–5 µM compared to the control (Fig. 2B). Considering that reduction of MMP in cells is related to cell death [9], DSF may cause the death of BMs. Annexin V-FITC/PI staining revealed that 1 and 5 µM DSF significantly induced the necrosis and late apoptosis of BMs (Fig. 2D).
To investigate which types of cells in BMs were affected by DSF, we performed flow cytometry; subset-specific marker analysis was performed using B220 and Gr-1 (Fig. 3). B220 is a B cell-specific marker [13] and Gr-1 is commonly used as a granulocyte marker [14]. Subset analysis demonstrated that DSF did not significantly affect the proportion of BMs.
A recent study reported that DSF/copper increased the level of ROS and the expression of superoxide dismutase (SOD)2, a closely associated enzyme [15]. Hydrogen peroxide produced by SOD in neutrophils has been considered the main mediator of ROS-induced neutrophil apoptosis [16]. These findings suggest that DSF may cause the death of BMs by increasing SOD and ROS production. The relationship between DSF, SOD, and damage to BMs need to be further studied.
To investigate whether DSF affects the metabolic activity and differentiation of BMs modulated by a growth factor or cytokine, we used GM-CSF, an essential growth/differentiation factor for BM-derived DCs [17]. The CCK-8 assay revealed that DSF affected the metabolic activity of GM-CSF-treated BMs in a concentration-dependent manner (Fig. 4A). The metabolic activity of GM-CSF-treated BMs was not significantly decreased by 0–0.06 µM DSF; however, it was markedly decreased by 0.125–0.5 µM DSF. Furthermore, flow cytometry using the DC-specific marker, CD11c, showed that 1–5 µM DSF significantly inhibited DC generation from BMs (Fig. 4B). These results demonstrated that DSF affects the metabolic activity of BMs modulated by GM-CSF and can block the differentiation of DCs from BMs over a certain concentration.
To evaluate the toxicity of DSF on BMs in vivo, we injected mice with DSF 50 mg/kg via the peritoneal cavity (Fig. 5A). The dose of DSF was determined based on several previous studies regarding the anti-cancer effects of DSF in vivo [18-20]. The results of BM cellularity showed that 50 mg/kg DSF did not significantly decrease the cell number of BMs in mice (Fig. 5B). On Day 5, BMs were harvested and marker analysis was performed; there was no significant proportional change in the subsets of BMs compared to the control (Fig. 5C, D). Therefore, we searched the potential defense mechanisms against DSF in vivo to explain the discrepancy between the in vitro and in vivo results. There are several antioxidant mechanisms in vivo including SOD that can reduce the oxidative stress caused by ROS [21,22]. In a previous in vitro study, DSF increased SOD and ROS [23]. It may work similarly in vivo; however, DSF 50 mg/kg is estimated to be tolerated in mice by antioxidant defense mechanisms. Although 50 mg/kg DSF did not induce in vivo toxicity in BMs in this study, it is likely that DSF doses over 50 mg/kg may induce myelosuppression in mice, given that 200, 400 or 800 mg/kg DSF have been shown to be genotoxic in BMs [24]. Future studies are needed to investigate DSF toxicity in vivo and action mechanisms.

 XML Download
XML Download