

 PDF
PDF Citation
Citation Print
Print



서 론
알레르기비염과 천식으로 대표되는 호흡기 알레르기 질환은 많은 면역세포와 사이토카인, 케모카인이 병태생리에 관여하며 표현형이 다양하게 나타난다[1] 질환의 면역학적 기전을 이해하고 치료제의 효능을 평가하기 위한 실험에 다양한 동물 모델이 활용되고 있는데, 그 중 마우스가 가장 널리 이용되고 있다. 마우스는 초보자도 다루기 편하고 구입 및 사육 비용이 저렴한 편이며, 번식이 쉽기 때문이다[2]. 무엇보다도 유전적인 정보가 잘 밝혀져 있어서 형질전환(transgenic), 유사 유전자(congenic), 특정 유전자가 결핍된(knockout) 마우스를 얻기가 용이할 뿐만 아니라 많은 근교 계통(inbred strain)을 사용할 수 있다는 장점이 있다[3]. 또한 면역학적 실험 방법이 잘 확립되어 있고, 단클론항체와 같은 세포 및 매개체 반응 분석에 사용할 수 있는 시약의 투여가 다른 동물에 비해 비교적 쉽다[4].
알레르기비염과 천식은 서로 동반되는 경우가 흔하고 알레르기비염 조절 정도가 천식의 발병 및 예후를 결정한다고 알려져 있는 등 서로 밀접한 연관이 있다[5]. 하지만 상기도와 하기도는 상피세포의 구성이 일치하지 않고 평활근세포가 기관지 점막하조직에서만 발견되는 등 해부학적인 차이가 있으며, 집먼지진드기(house dust mite, HDM) 항원에 노출되었을 때 선천면역반응이 상기도에서는 Toll-like receptor 2 (TLR2)가, 하기도에서는 TLR4가 활성화되는 등 면역학적인 차이점도 많다[6].
이에 본 논문에서는 마우스를 사용한 호흡기 알레르기 질환 문헌들을 정리하고, 알레르기비염 마우스 모델과 천식 마우스 모델의 유사한 점과 차이점을 살펴보고자 한다. 또한 마우스 모델을 제작하는 방법과 고려할 점 및 한계점에 대해 기술하여 연구에 적용할 수 있는 방법에 대해 고찰하고자 한다.
Go to : 
마우스 종에 따른 호흡기 알레르기 질환의 발현
현재 C57BL/6, BALB/c, A/J, C3H/HeJ, DBA/2 등 다양한 종(strain)의 마우스들이 실험에 사용되고 있으며, 마우스 종에 따라서 알레르기 질환의 유도 정도에 많은 차이가 있어서 실험에 주의가 필요하다.
천식 마우스에서 많은 연구가 이루어졌는데, A/J 마우스는 가장 심한 호흡기 알레르기가 잘 유도되는 종이며 C3H/HeJ 마우스는 호흡기 알레르기에 저항성이 있는 것으로 알려져 있고, A/J 마우스는 호산구 침윤이 많고 보체(complement) 중에 C5a가 결핍되어 알레르기 천식이 심하게 나타난다[7,8]. 또한 메타콜린에 대한 기도과민성 정도가 A/J에서 높아서, 종의 유전적인 특성도 중요한 영향을 미친다[9].
형질전환 혹은 특정 유전자가 결핍된 마우스는 대부분 C57BL/6종을 기반으로 생성되며 일부 BALB/c종에서도 생성되므로, C57BL/6종과 BALB/c종을 실험에서 많이 사용하게 된다. BALB/c는 Th2 면역반응, C57BL/6는 Th1 면역반응으로 좀더 편향되어 있는 것으로 알려져 있다. HDM extract를 사용한 천식 모델에서는 기도저항성과 Th2 사이토카인의 발현, 폐의 염증세포 침윤이 BALB/c에서 C57BL/6보다 증가하였으며, 2형 면역반응에 관여하는 인자인 tyrosine kinase interleukin-2 inducible T cell kinase가 결핍시 C57BL/6에서는 HDM 유도 기도염증에 저항성(resistance)이 생기고, BALB/c에서는 감수성(susceptibility)이 줄어든다고 보고되었다[10]. 이는 BALB/c에서 기도 과민성이 더 높고 폐 조직에서 비만 세포의 수와 Th2 사이토카인 중에 interleukin (IL)-13의 발현이 더 높게 나타나는 것으로 보고되었으며[11], 불변 자연살해 T세포(invariant natural killer T[iNKT] cells)의 아형 중에 NKT2 세포가 BALB/c 마우스의 장간막 림프절에 더 많이 분포하여 IL-4가 더 많이 생성되기 때문으로 설명되고 있다[12].
HDM 항원인 Dermatophagoides farinae (Der f1)를 이용한 알레르기비염 모델에서 호산구와 호중구의 침윤 정도에는 두 종간의 유의한 차이가 없었으나, 혈청 총 immunoglobulin E (IgE)와 Der f1 특이 IgE, IgG1, IgG2의 발현이 BALB/c에서 유의하게 증가하고 IL-4, eotaxin-1, eotaxin-2, CXCCL- 1, CXCCL-2의 발현 역시 BALB/c에서 C57BL/6보다 더 증가하였다[13].
Go to :
마우스 성별에 따른 호흡기 알레르기 질환의 발현
일반적으로 암컷이 수컷보다 알레르기 염증 반응이 더 잘 일어나는 것으로 알려져 있는데, 암컷 마우스에서 수컷 마우스보다 기관지폐포세척액(bronchoalveolar lavage fluid, BALF)에서 호산구의 발현이 증가하고, 폐에서 Treg이 더 감소하여 Th2 면역반응이 수컷보다 더 증가하였다[14]. BALB/c 마우스 실험에서 암컷의 폐와 BALF에서 수컷보다 난알부민(ovalbumin, OVA)과 IL-33을 투여 시 2형 면역반응(type 2 inflammation)이 더 심하게 나타났으며, 사이토카인 및 케모카인의 생성, 호산구의 활성화 및 2형 선천성 림프구세포(group 2 innate lymphoid cells, ILC2s)의 발현이 더 증가함이 확인되었는데[15], 이는 안드로젠이 ILC2s의 기능을 억제하며, ILC2s의 유전적 발현이 본질적으로 암컷 마우스에서 수컷에서 보다 더 활성화된다는 보고와 일치한다[16]. C57BL/6 실험에서 나노입자(nanoparticle)를 투여하여 기도 호산구증 가증을 유도 시에 암컷에서 기도저항성이 더 증가하였고, 대식세포(macrophage)에서 IL-5와 IL-1β의 발현이 더 증가하며 대체 활성화된 2형 대식세포(alternatively activated macrophages)의 아형인 M2a와 연관된 유전자의 발현이 증가하는 것이 보고되었다[17].
Go to :
호흡기 알레르기 질환 모델의 항원
일반적으로 동일한 항원을 이용하여 알레르기비염 모델과 천식 모델을 제작할 수 있다. 고전적으로 달걀에서 유래한 OVA를 이용한 마우스 모델이 널리 사용되었는데, 비용적으로 저렴하고 순도가 높아서 질환이 잘 유도되며, OVA 특이적인 T세포 수용체를 발현하는 형질전환 마우스(OT-I와 OT-II 마우스)를 실험에 사용할 수 있는 등 OVA를 활용한 면역학적인 기전 분석에 도움이 되기 때문이다[18]. 하지만 OVA를 단독으로 투여할 때는 Th2 면역반응이 아닌 면역관용(immune tolerance)이 생겨서 면역증강제(adjuvant)를 같이 투여해줘야 하며, OVA는 인간에서 호흡기 알레르기 질환을 일으키는 항원이 아니라는 한계점이 있다[19].
면역증강제는 aluminum hydroxide (alum), lipopolysaccharide (LPS), tumor necrosis factor 등이 활용되며 alum이 가장 흔히 사용된다. Alum은 염증성 수지상세포의 요산 활성화를 통해 면역반응을 유도하여 알레르기비염이나 천식 모델을 쉽게 유도할 수 있게 해주나[20], alum을 투여 시에는 pathogen associated molecular patterns의 필요성이 없어서 선천면역경로 역시 우회할 수 있어서, 항원에 대한 즉각적인 면역 반응이 변경되어 나타날 수 있다는 점을 고려해야 한다[21]. 투여하는 면역증강제의 용량이나 종류에 따라 질환의 면역 반응을 조절할 수도 있는데, OVA를 고용량의 LPS와 같이 투여 시에는 Th1 반응, OVA를 저용량의 LPS와 같이 투여 시에는 Th2에 가까운 면역반응이 유도되는 것으로 알려져 있고[22], OVA를 alum과 같이 투여하면 호산구성 천식이 생기는 반면에 OVA를 complete Freud’s adjuvant와 같이 투여하면 호중구성 천식이 유도됨이 보고되었다[23].
연구자들은 실제 사람에서 알레르기 질환을 유발하는 집먼지진드기, 곰팡이, 바퀴벌레, 꽃가루 등의 항원의 추출물 혹은 정제된 단백질을 이용한 호흡기 알레르기 질환 마우스 모델을 보고하였다. 가장 흔한 알레르기 항원인 집먼지진드기의 추출물(HDM extract)을 이용하거나[24] 미가공(crude) 상태의 HDM extract로부터 동정 및 정제된 Derp[25] 혹은 Derf13)를 사용하면 효과적으로 질환이 유도되는데, 이는 고유한 효소 활성, 면역원성 및 알레르기 염증반응을 촉진하는 면역세포의 Dectin-2 수용체를 통한 직접 활성화 등의 기전에 의한다[26]. 알레르기비염과 천식의 주요 항원인 Aspergillus fumigatus [27]나 Alternaria alternata [28] 같은 곰팡이도 마우스에 투여 시 IgE의 증가와 호산구의 발현 및 염증반응을 유발한다. 꽃가루는 계절성으로 주요한 알레르기의 원인인데 ragweed를 사용한 모델이 보고되었으며[29], 바퀴벌레 항원으로는 Bla g2가 주로 사용된다[30]. 이러한 항원들은 단독 혹은 혼합해서 마우스에게 투여된다.
Go to :
항원의 투여 방법 및 마우스 모델 프로토콜
호흡기 알레르기 질환의 마우스 모델 프로토콜은 연구자에 따라 매우 다양하나, 일반적으로는 항원을 감작(sensitization) 시킨 후에 항원을 유발(provocation)하는 과정을 거쳐 완성된다.
첫 번째 단계인 항원을 감작시키는 과정은 전신적인 면역반응을 유도하기 위해 필요하며, 알레르기비염 모델과 천식 모델 모두에서 일반적으로 시행된다. 저자는 항원의 감작 없이 유발 과정만 거칠 경우 호흡기에 염증세포는 어느정도 침윤이 되나, 혈청에서 항원 특이 IgE와 IgG가 증가하지 않는 것을 확인하였다[31]. 항원의 감작은 복강을 통해 투여하는(intraperitoneal) 방법이 고전적으로 가장 많이 사용되고 있는데, 주로 OVA [23,32,33]나 HDM [13,24,25]과 같은 항원을 alum과 같은 면역증강제와 같이 복강 내로 투여하며, 횟수는 1회에서 3회까지 연구자에 따라 다양하나, 감작과 감작 사이의 간격은 1주일이나 2주일 정도의 간격을 두는 것이 일반적이다. 복강 내 투여가 실제 사람에서는 면역반응을 일으키는 기전이 생리적이지 않고 사람에서 면역치료를 하는 루트로 피하조직(subcutaneous) 경로가 많이 사용되고 있다는 점을 근거로 마우스에서도 Derp 항원을 피하로 감작시키는 천식 모델이 보고되었다[34]. 또한 사람에서 실제 항원을 흡입하는 통로인 코를 통해서(intranasal) HDM 항원을 감작시키는 방법도 보고되었다[35].
두 번째 단계인 항원을 유발시키는 과정은 코나 폐에 국소적으로 염증 반응을 유도하기 위해 상기도 혹은 하기도에 항원을 투여하게 된다. 보통 항원 감작을 시행한 마지막 날로부터 1주일 정도 후에 시행하며, 항원 유발의 횟수는 3회에서 7회까지, 날마다 연속해서 시행하는 것이 일반적이다.
알레르기비염 모델의 경우, 항원 투여 경로로 비강 내로 점적하는 방법이 널리 사용된다. 폐로 항원이 흡인되지 않도록 마취를 하지 않은 상태에서 진행하며[13,32], 마우스의 목과 등을 손가락으로 잡아서 움직이지 않게 한 후에 파이펫을 통해 마우스의 양쪽 콧구멍 입구에 번갈아 가면서 항원을 투여하는데, 삼키거나 폐로 흡인되지 않게 주의한다. 마우스가 자연스럽게 숨을 들이마시면서 항원을 잘 흡입하는지 관찰하면서 투여하는 것이 중요하다. 과다하게 많은 항원을 단시간에 투여할 경우 흡인의 위험성이 있으며, 마우스의 목 주위를 너무 세게 잡을 경우 질식할 위험이 있어서 주의가 필요하다 (Fig. 1A).
 | Fig. 1.Experimental method for provocation of allergen. A: Intranasal challenge. By dripping the allergen little by little into the nostrils, care should be taken so that the mouse does not swallow or aspirate allergen into the lungs. B: Aerosol challenge. Under the supply of electric power (red arrow, power supply and compressor of nebulizer), dissolved allergen in the nebulizer kit (purple arrow) is distributed into the chamber through mouthpiece (green arrow) in the form of a mist.
|
천식 모델의 경우, 비강 내로 점적 시 마취를 시행한 상태에서 진행하면 폐로 항원이 쉽게 흡인되어 천식을 유도할 수 있다[24,33]. 비강 내 투여는 일반적으로 항원을 여러 차례 반복해서 투여해야 염증반응이 충분히 유도되는 특성이 있어서, 천식 모델을 제작하기 위해서는 마취하에 기관지 내로 직접 항원을 투여하는 방법을 쓰기도 한다[21,36]. 이상의 방법은 마취를 해야 할 뿐 아니라 마우스에게 침습적인 편이기에, 가습기 챔버에 마우스를 위치시키고 항원을 분무시켜서 에어로졸 형태로 흡입하게 하여 투여하는 방법도 널리 사용된다[37-39]. 저자는 27×20×20 cm 크기의 챔버에 마우스 5-6마리를 같이 넣고 nebulizer kit를 연결하여, 에어로졸 형태의 항원을 균일하게 마우스들이 흡입할 수 있도록 하고 있다(Fig. 1B).
이와 같은 항원 투여 과정이 끝나면 보통 마지막 항원 투여 후 24시간 이내에 부검을 하여 분석을 한다. 이는 시간이 경과함에 따라 기도 염증의 표현형과 조직에서의 면역반응이 회복되어 버리기 때문이며, 천식 모델의 경우 마지막 항원 유발 후 2주가 지나면 기도저항성과 기도 점액질 발현 및 BALF에서 염증세포 침윤 정도가 대조군과 차이가 없을 정도로 회복되는 것으로 알려져 있다[40].
저자가 OVA 및 HDM을 사용하여 알레르기비염 및 천식 모델을 만드는 프로토콜을 Fig. 2에 정리하였다. 암컷 4-6주 령의 마우스를 주로 사용하며, BALB/c종이 알레르기의 표현형이 잘 발현되기는 하나, 관찰을 하고자 하는 형질전환 혹은 특정 유전자가 결핍된 마우스의 유전적 배경에 따라 C57BL/6N 혹은 C57BL/6J종을 이용할 수 있다. OVA 혹은 HDM을 alum과 함께 복강 내로 투여하는 감작 과정을 2-3회 거친 후에 항원을 비강 내 혹은 에어로졸로 4-5회 투여하는 유발 과정을 사용하고 있다.
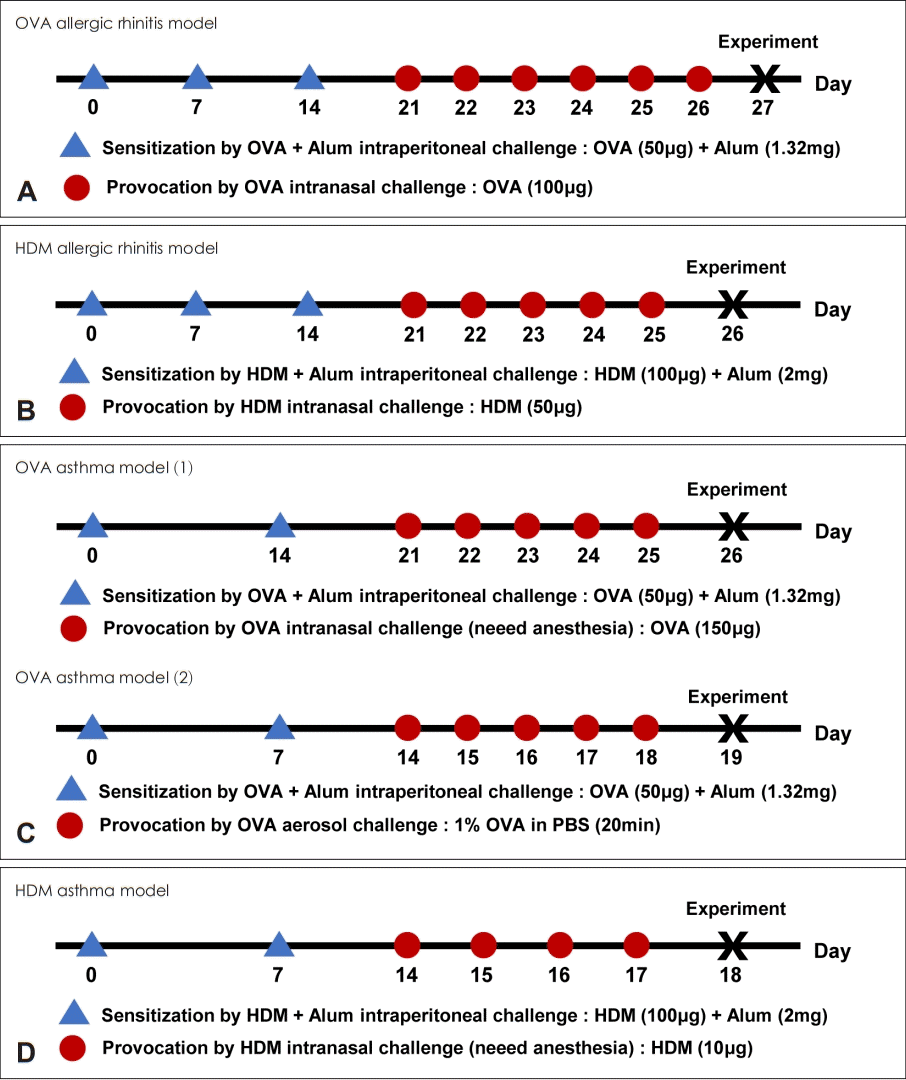 | Fig. 2.Experimental protocol for the development of allergic rhinitis and asthma in mouse. A: OVA-induced allergic rhinitis model. B: HDM-induced allergic rhinitis model. C: OVA-induced asthma model, either by intranasal challenge or aerosol challenge. D: HDM-induced asthma model. OVA, ovalbumin; HDM, house dust mite; Alum, aluminum hydroxide.
|
Go to :
만성 호흡기 알레르기 질환 마우스 모델
대부분 연구자들이 수행하는 마우스 모델은 1개월 이내로 제작이 끝나는 급성 모델이지만, 실제로 알레르기비염과 천식은 만성적인 질환이기 때문에 항원을 장기간 투여하는 만성 모델에 대한 연구도 필요하다.
알레르기비염 모델의 경우, OVA를 3개월 동안 에어로졸로 투여받은 마우스의 코에서 상피하 섬유화, 배상세포(goblet cell) 증식, matrix metalloproteinase 9 및 tissue inhibitors of metalloproteinase 1의 증가가 1개월 동안 투여한 마우스보다 월등히 증가하였으며, 폐에서의 기관지 주변 섬유화 역시 더 증가하여 기도의 재형성(remodeling)이 일어남을 확인하였다[41]. OVA를 비강 내로 총 6주 혹은 11주 동안 투여하여 생성된 알레르기비염 모델에서 호산구 침윤과 상피세포 증식이 호흡상피에서는 증가하지만 후각상피에서는 증가 정도가 미미하여 상피세포의 종류에 따라 알레르기 염증이 미치는 정도가 다름을 보고한 문헌이 있고[42], OVA와 alum을 복강 내로 3차례 투여하여 감작시킨 후, 비강 내로 OVA를 일주일에 3번씩, 3주간 투여한 후에 에어로졸로 OVA를 일주일에 3번씩 총 6주간 투여하여 총 78일 동안 모델을 제작하여 알레르기비염과 천식의 표현형을 동시에 관찰한 문헌이 있다[43].
천식에서도 많은 연구들이 있는데, 에어로졸로 OVA를 투여하는 유발 과정을 한차례 시행한 뒤 2주 후에 OVA 유발 과정을 한 번 더 시행하여 제작된 중증 천식 모델[38], HDM을 비강 내로 투여하는 유발 과정이 끝나고 6주후에 HDM을 다시 투여하는 재노출 천식 모델 및 HDM 감작 과정 처음부터 일주일에 3번씩 HDM을 총 6주간 투여하는 만성 천식 모델에 대한 연구가 보고되었다[44]. 또한 OVA와 HDM, 바퀴벌레 항원(cockroach allergen, CRA)를 혼합하여 복강 내로 투여하여 감작시킨 후에 OVA, HDM, CRA를 번갈아가며 7주 동안 투여하여 유발시켜서 Th2 면역반응 및 기도저항성이 증폭되며 기도평활근의 두께와 폐섬유화가 증가하고 dexamethasone에도 저항성을 보이는 중증 천식 마우스 모델이 보고되었다[45].
Go to :
면역세포 투여를 통한 호흡기 알레르기 질환 마우스 모델
그외에, 알레르기 면역 반응의 특정 측면을 분석하거나 증폭된 면역 반응을 유도하기 위해 T세포나 수지상세포(dendritic cells)를 직접 마우스에게 투여하는 방법도 있다. T세포의 다양한 아형의 역할을 분석하기 위해 OVA 특이적인 T세포 수용체를 발현하는 형질전환 마우스에서 분리한 T세포 클론을 마우스에게 투여하여 감작시키는 방법이 고안되었다[46]. 또한 T 세포에게 항원을 전달하는 기능을 하며 알레르기 면역반응을 개시하고 유지하는데 중요한 수지상세포를 투여하기도 하는데, OVA 혹은 Aspergillus fumigatus 등의 항원으로 활성화된 마우스 골수 유래 수지상세포를 비강 내 혹은 기관지로 투여하는 감작 과정을 통해 각각 알레르기비염과[47] 천식 모델을[48,49] 제작한 연구들이 보고되었다. 저자는 항원 유발과정에서 OVA로 활성화된 마우스 골수 유래 수지상세포를 정맥으로 투여하여, 기존 프로토콜보다 더 증폭된 Th2/Th17 면역반응을 나타내고 호산구와 호중구가 동시에 침윤되며 스테로이드 투여에도 저항성을 보이는 천식 마우스 모델을 제작하였고, 이는 사람에서 이질적인 형태로 나타나는 천식의 특성을 잘 구현하고 있다[31]. 이상의 항원의 감작 및 유발시키는 방법에 따른 장단점을 Table 1에 정리하였다.
Table 1.
Comparison of several sensitization or provocation methods to induce allergic rhinitis and asthma in mice
| Routes | Technical and experimental aspects | Immunological and pathophysiological aspects |
|---|---|---|
| Intraperitoneal [13,24,25,30] | • Most traditional and conventional method | • Usually requires an adjuvant |
| • Few doses required | • Not physiologic route of human sensitization | |
| • Sedation not required | ||
| Subcutaneous [23,34] | • Few doses required | • Usually requires an adjuvant |
| • Sedation not required | • Not physiologic route of human sensitization | |
| Intranasal [15,28,29,32,33] | • Many instillations required | • Mimics human sensitization |
| • Risk of aspiration | ||
| • Variation can occur between individuals | ||
| • Sedation required for asthma model | ||
| Intratracheal [21,36] | • Invasive | • Mimics human sensitization |
| • Few doses required | ||
| • Sedation required | ||
| Aerosol [37-39] | • Requirement of high allergen dose | • Mimics human sensitization |
| • Requirement of considerable time in aerosol chamber | • Possibility of skin allergic inflammation by allergen exposure | |
| • Sedation not required | ||
| Immune cell transfer [31,46-49] | • Need effort for preparation of cells | • Induce robust immune response |
| • High productivity | • Possibility of systemic allergic inflammation | |
| • Not physiologic route of human sensitization |
![]()
Go to :
알레르기비염 마우스 모델의 지표
마우스 모델을 통해 궁극적으로 사람에서 나타나는 질환의 특성과 지표를 확인하는 것이 중요하다. 사람에서 알레르기 비염의 특징인 맑은 콧물, 재채기, 가려움증, 코막힘 중에 재채기를 하는 횟수를 측정하며 가려움의 정도는 간접적으로 코를 비비는 횟수를 관찰한다. 일반적으로 2명 이상의 관찰자가 마지막 항원 투여를 한 후 20분 정도 관찰하여 재채기와 코를 비비는 횟수를 합하여 결과로 기록하는데 주관적인 지표이기 때문에 해석에 주의가 필요하다[13].
기도에 염증세포가 침윤하는 것을 조직병리 사진을 통해 관찰하는 것이 필수적이다. 마우스의 머리 부분을 채취하여 파라핀 블록으로 제작한 후, 내장된 조직을 절단하여 단면을 확인하며, 코의 비중격 및 비강벽을 따라 호흡상피를 관찰한다. Hematoxylin and eosin 염색을 통해 조직의 전체적인 염증세포 침윤 정도를 확인하고 periodic acid-Schiff 염색을 통해 배상세포의 과증식 정도를 확인하며, Sirius red 염색을 통해 호산구 침윤을 관찰하고, Masson’s trichrome 염색으로 상피세포 층의 두께와 콜라젠 침윤 정도를 확인할 수 있다. 각 조직 절단면에서 무작위로 여러 곳을 선정하여 고배율(high power field)당 보고자 하는 세포의 개수의 합을 결과로 제시하거나[32], 염증세포의 분포 정도를 정량화된 scoring 방법이나 ImageJ와 같은 소프트웨어를 통한 면적 계산방법을 통해 확인할 수 있다[33]. 그 외에 보고자 하는 단백질에 붙는 특이 항체를 이용하여 면역조직화학(immunohistochemistry) 및 면역형광(Immunofluorescence) 염색을 하면 조직에 특이하게 나타나는 단백질을 세밀하게 분석할 수 있다[25].
혈청에서 면역글로불린(Ig)을 측정하는 것도 유용한데, total IgE 및 OVA나 HDM과 같은 항원에 특이적인 IgE 및 IgG1, IgG2a, IgG2b, IgG2c 등을 확인한다. 혈청은 마우스의 안와 혹은 심장에서 채혈을 하여 얻게 되며 enzyme-linked immunosorbent assay (ELISA)를 통해 면역글로불린의 발현 정도를 확인할 수 있다[33].
면역학적인 분석을 위해 사이토카인의 발현 정도를 ELISA 혹은 polymerase chain reaction을 이용하여 측정하며, 면역세포의 수와 발현 정도 및 전사인자(transcription factor), 세포 내 사이토카인의 발현을 유세포분석(flow cytometry)을 통해 측정한다. 이는 코 점막과 코세척액(nasal lavage fluid) 혹은 경부 림프절을 채취하여 결과를 확인할 수 있다. 코세척액은 카테터를 인두 개구부위로 삽입하여 콧구멍으로 나오게 하여 phosphate-buffered saline (PBS)을 주입하여 흘러나오는 용액을 모아서 얻을 수 있다(Fig. 3A) [50].
 | Fig. 3.Parameter for mouse model of allergic rhinitis and asthma. A: Method to obtain nasal lavage fluid. A catheter is inserted through the pharyngeal opening into the choana, then lavage fluid from nasal cavity is collected in the tube. B: Method to obtain airway hyperresponsiveness. After tracheostomy, mouse is connected to a computer-controlled small animal ventilator and ventilated using flexiVent system (SCIREQ Montreal, Canada). Lung resistance is measured in response to increasing doses of aerosolized methacholine at the nebulizer (yellow arrow). C: Method to obtain bronchoalveolar lavage fluid. It is obtained by lavaging the lower airways of mouse with phosphate-buffered saline via a tracheal cannula.
|
Go to :
천식 마우스 모델의 지표
천식 모델의 경우 사람에서 천식의 중요한 특징인 가역적 기도폐쇄와 기도과민성을 관찰하는 것이 필수적이다. 사람에서 콜린작용제(cholinergic agonist)와 같은 약제의 투여 후에 1초간 강제호기량(forced expiratory volume in 1 second)과 강제폐활량(forced vital capacity)을 확인하는 것처럼, 마우스에서도 콜린작용제에 대한 과민반응을 확인하는데 2가지 방법을 사용한다. 첫 번째로, 마취하에 기관 내 삽관을 하여 기도 내의 압력을 측정하는 방법이 있는데, 메타콜린과 같은 콜린작용제의 투여 용량을 늘려가면서 기도 저항성 및 탄성을 측정한다(Fig. 3B) [51]. 이는 침습적이고 측정 후에는 동물을 희생시켜야 하는 단점이 있다. 두 번째로, 마취하지 않는 자연 운동 상태에서 체적변동 측정기(whole body plethysmography)를 통해 기도의 폐색을 반영하는 수치인 enhanced pause (Penh)를 측정하는 방법이다[52]. 이는 장시간 혹은 반복적으로도 측정이 가능한 장점이 있다.
마우스 폐의 엽을 채취하여 조직병리 사진을 통해 폐 실질 조직과 기관지 및 기관지 주변 공간을 관찰한다. 사이토카인의 발현 정도와 면역세포의 분포, 전사인자의 발현을 폐 조직 및 기관지폐포세척액(BALF) 혹은 종격동 림프절을 채취하여 조사할 수 있다.
기관지폐포세척액은 기관지 절개술을 시행한 후 카테터를 폐 쪽으로 향하게 넣은 후 PBS를 주입하고 씻어낸 용액을 모으게 되는데, 실린지로 PBS를 주입하거나 용액을 빼낼 때 너무 속도를 빠르게 하게 되면 폐포 및 기관지 세포의 손상을 유발할 수 있으므로 주의가 필요하다. 차가운 PBS를 사용하고 1 mL의 PBS에 0.5 mM ethylenediaminetetraacetic acid (EDTA)를 혼합한 용액을 사용하면 세척액의 질을 높일 수 있다(Fig. 3C) [21].
세척액은 면역세포와 사이토카인의 양이 많지는 않기 때문에 림프절을 배양하여 실험하는 것도 유용하다. 호흡기 알레르기 질환에서 주요한 역할을 하는 호산구, T세포, 수지상세포, 대식세포 등의 숫자와 구성정도 등은 유세포분석을 통해 세밀하게 측정할 수 있다. 이와 같은 지표를 분석하여, 최소 3회 이상의 연속된 모델 제작에서 비슷한 소견이 재현되는지를 판단하는 것이 필요하다.
Go to :
호흡기 알레르기 질환 마우스 모델의 적용 시 고려할 점
알레르기비염 모델과 천식 모델은 앞서 기술한 바와 같이 마우스의 종에 따라 질환 발현의 차이가 있다는 점이 유사하고 모델 제작에 사용하는 항원이 비슷하다. 조직에서 호산구나 배상세포와 같은 염증세포의 침윤을 관찰하고 혈청에서 항원 특이 면역글로불린을 측정하며 Th2 면역반응을 확인하게 된다. 하지만 알레르기비염 모델이 천식 모델과 다른 특징들이 있어서 실험을 할 때 고려해야한다. 첫째, 모델을 제작하는 프로토콜 중에 항원을 기도에 투여하는 유발 과정에 차이가 있다. 둘째, 알레르기비염 모델은 천식 모델에서의 기도과민성과 같은 객관적인 생리학적인 검사가 없다. 셋째, 마우스의 비강은 실험 환경의 영향으로 기저 염증이 있을 수 있어서 코세척액에서도 호중구와 단핵구가 흔히 관찰될 수 있다[53]. 넷째, 마우스의 비강은 폐에 비해 호산구의 증가가 적게 일어난다[54].
마우스 모델에서 얻은 결과를 사람의 질환에 똑같이 적용하기에는 몇 가지 한계점이 존재한다. 우선 해부학적으로 사람은 비강 대부분이 호흡상피로 덮여있는 반면에 마우스는 비강의 거의 절반을 후각상피가 차지하며, 비강의 전방에는 호흡상피, 후방에는 후각상피가 분포한다. 따라서 마우스에서는 주로 전방의 비중격이나 외측벽을 주로 분석하게 된다[55]. 또한 마우스 모델에서는 침윤된 호산구의 탈과립 및 상피세포 내 축적 현상이 나타나지 않으며, 마우스의 기도평활근은 leukotriene, histamine, bradykinin 등의 매개물질에 반응하지 않는다[56]. 무엇보다도, 알레르기비염이나 천식은 사람에서만 일어나는 질환으로 마우스에서는 자연적으로 발생하지 않기에 인위적으로 항원을 투여하여 유발을 시키다보니 기도의 염증이 사람에서처럼 오래 지속되지 않는다. 이러한 마우스 모델을 활용하여 연구를 진행할 때, 연구자는 마우스 종과 성별, 투여 항원의 종류 및 투여방법 등에 따라 결과에 다양한 차이가 나타난다는 점을 유념하여 실험 목적에 맞게 디자인하는 것이 필요하고, 데이터 분석 시에도 주의를 기울여야 한다.
Go to :
 XML Download
XML Download