

 PDF
PDF Citation
Citation Print
Print



Gliomas are responsible for 70% of brain tumors [1]. Although rare (correspond to less than 2% of tumors), these are the second leading cause of death of cancer in patients of pediatric age [2] making this cancer an important public health problem. Glioblastomas correspond to almost half of the cases of malignant brain tumors [1]. They usually affect adults, with a peak incidence between 45 and 75 years [1,3]. Glioblastomas (GBMs) are lethal tumors, and less than 10% of patients live 2 years after diagnosis [4,5]. They are highly infiltrative; thus, the complete surgical resection usually is not possible [3]. In addition to surgery, standard therapy was proposed by Stupp and collaborators, through radiotherapy with concurrent chemotherapy using temozolomide, however the average life span of patients is 14 months [6].
Epidermal growth factor-like domain multiple 7 (EGFL7) is a pro-angiogenic factor, largely restricted to the endothelium [7]. This protein plays a specific role in vascular tubulogenesis, regulates angiogenesis [8] and acts in vascular development of the placenta and embryonic growth in mammals [9]. Evidence suggests that EGFL7 expression plays a role in tumor progression through mediation of metastasis, proliferation, and angiogenesis [10,11]. Moreover, EGFL7 overexpression is associated with poor prognosis events on different tumor types such as gliomas [12–14] and colorectal cancer [10,15]. Also, interaction of EGFL7 with epidermal growth factor receptor (EGFR), a key oncogene, is directly related to the development, growth, and metastasis of several tumor types, such as gliomas [13].
In our previous studies, we found recurrent chr7q34 gain in gliomas, encompassing EGFL7 gene [16], and high EGFL7 expression associated with an unfavorable outcome of pilocytic astrocytoma patients [14]. Therefore, we extended the investigation of the potential prognostic of EGFL7 expression in GBM isocitrate dehydrogenase (IDH)-wildtype through in silico and immunohistochemistry approaches, and also proposed signaling pathways that EGFL7 can be contributing to gliomagenesis.
MATERIALS AND METHODS
In silico analysis
We performed in silico analysis in R software, using the TCGA2STAT package [17] and the tools of cBio [18] and DAVID [19] portals.
In order to evaluate potential pathways related to EGFL7 expression, correlation analysis was performed in cBio portal. RNA sequencing (RNASeq) data of GBM was obtained from The Cancer Genome Atlas (TCGA) (n = 152 patients), and the genes with Spearman’s correlation ≥ |0.6| were considered strongly correlated to EGFL7 expression. Those were submitted to gene ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis in DAVID portal, and GO/KEGG terms with p ≤ 0.05 were considered as statistically significant.
Additionally, RSEM-normalized data of expression (RNASeq) of GBM IDH-wildtype of the TCGA was obtained, together with the clinical data (n = 138). The number of reads of all genes were log10-transformed. Further, Z score was calculated for EGFL7, and patients with Z score ≥ 1.0 were classified with high EGFL7 expression. In order to evaluate the prognostic value of EGFL7, overall survival of the patients was analyzed in Kaplan Meier curves using log-rank statistical test.
Patients
Glioblastoma IDH-wildtype tissues were obtained from a series of 74 patients who had undergone surgery at Barretos Cancer Hospital. Formalin-fixed, paraffin embedded slides were reviewed by expert neuropathologists (A.P.B., L.N., and G.R.T.), who confirmed glioblastoma IDH-wildtype diagnosis according to 2021 World Health Organization central nervous system classification [1,20] and selected areas for tissue microarray construction. Overall, the age ranged from 26 to 75 years (median of 57 years), who were stratified in 20–59 years old (45 patients) and > 59 years old (29 patients). The average overall survival was 7.61 months (ranging from 0.13 to 45.50 months). All patients were deceased.
EGFL7 immunohistochemistry
The EGFL7 immunohistochemistry reactions were performed according to Brunhara et al. (2021) [14]. Briefly, slides with sections of 4 μm were deparaffinized and rehydrated. Then, antigenic retrieval was performed in tris(hydroxymethyl)aminomethane (Tris)–ethylenediaminetetraacetic acid buffer for 20 minutes at 98ºC. Inhibition of endogenous peroxidases and non-specific proteins was performed with the Novolink Polymer Detection System immunohistochemistry kit (Leica Biosystems, Wetzlar, Germany). The rabbit anti-EGFL7 antibody (cat. No. ab115786, Abcam, Cambridge, MA, USA) was applied at a 1:100 dilution on the slide, for 90 minutes at room temperature. Further, immunoreaction product complexes were treated by Novolink Polymer Detection System (30 minutes of post-primary antibody and 30 minutes of polymer at room temperature). Staining was performed using 3,3′-diaminobenzidine and counterstaining was performed with Gill-2 hematoxylin.
The reactions were analyzed by an expert neuropathologist (L.N.) using intensity (0, negative; 1, weak; 2, medium; and 3, strong) and extension (0, negative; 1, ~25% of mark; 2, between 25% and 50%; and 3, more than 50% of mark) scores. The scores were summed, and the expression was considered low (sum between 0 and 3) or high (sum between 4 and 6) [14]. Endothelial cells were used as positive internal control.
Statistical analysis
Immunohistochemistry staining was associated with clinicopathological data (location, age, sex, and Karnofsky Performance Status [KPS]). Finally, overall survival was analyzed using Kaplan Meier curves. Statistical analysis was performed using chi-square tests or Fisher exact test (clinicopathological), log-rank (overall survival), or univariate Cox proportional hazards. Contrasts with a p ≤ .05 were considered statistically significant.
RESULTS
Through in silico analysis, we found 78 genes significantly associated with EGFL7 expression profile in GBM patients of TCGA database (Supplementary Table S1). These genes were found in 40 enriched GO terms, most of them related to angiogenesis/vasculogenesis, Notch pathway, and cell adhesion (shown in Fig. 1, Supplementary Table S2). Furthermore, pathway analysis using KEGG showed eight enriched terms, mainly related to adhesion, blood cells or blood vessel events, and phosphoinositide 3-kinase (PI3K)–Akt, Notch, and Rap1 signaling pathways (shown in Fig. 1, Supplementary Table S3). Survival analysis of TCGA data showed no difference in overall survival regarding EGFL7 gene expression (p = .182, shown in Fig. 2A).
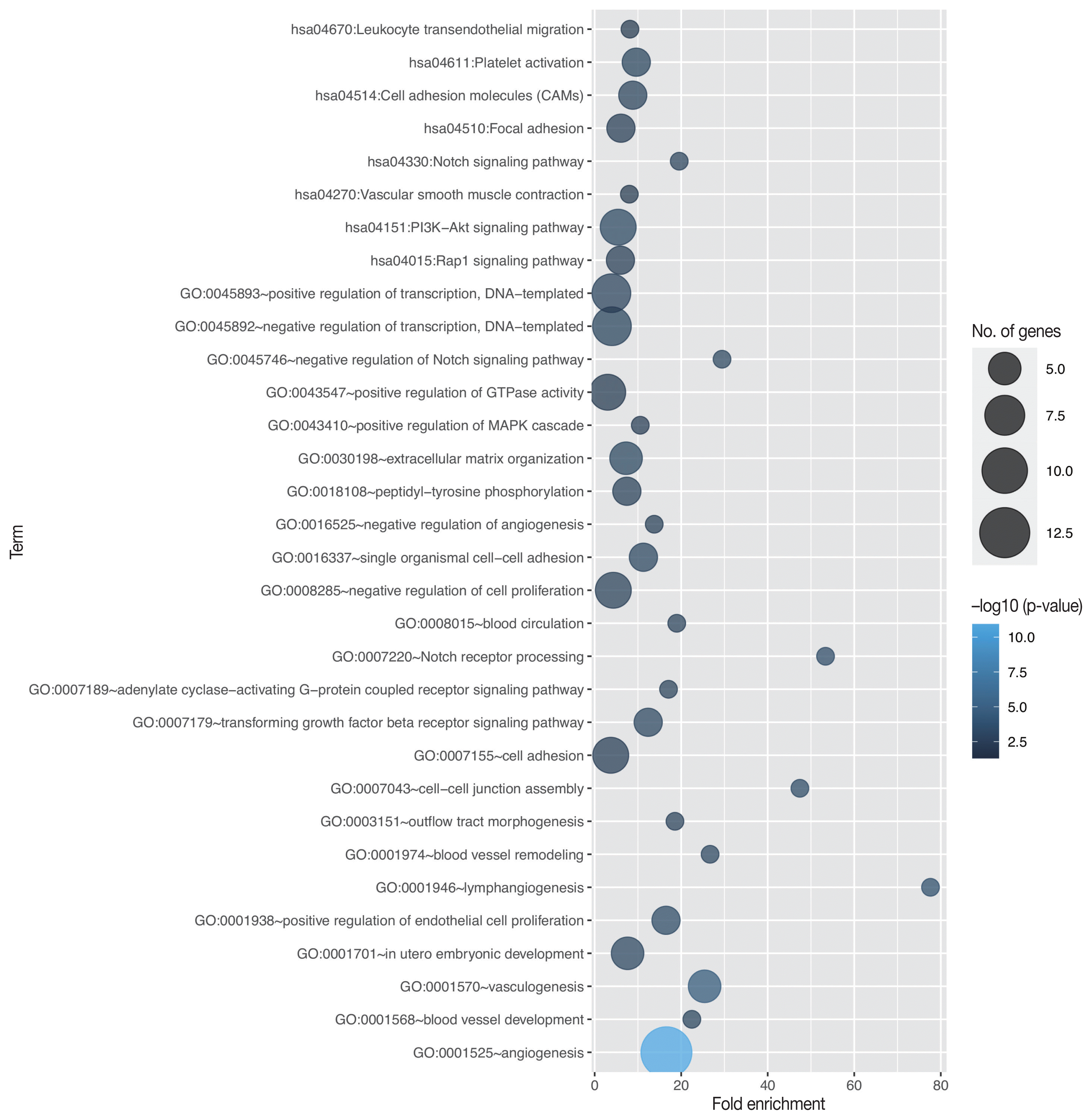
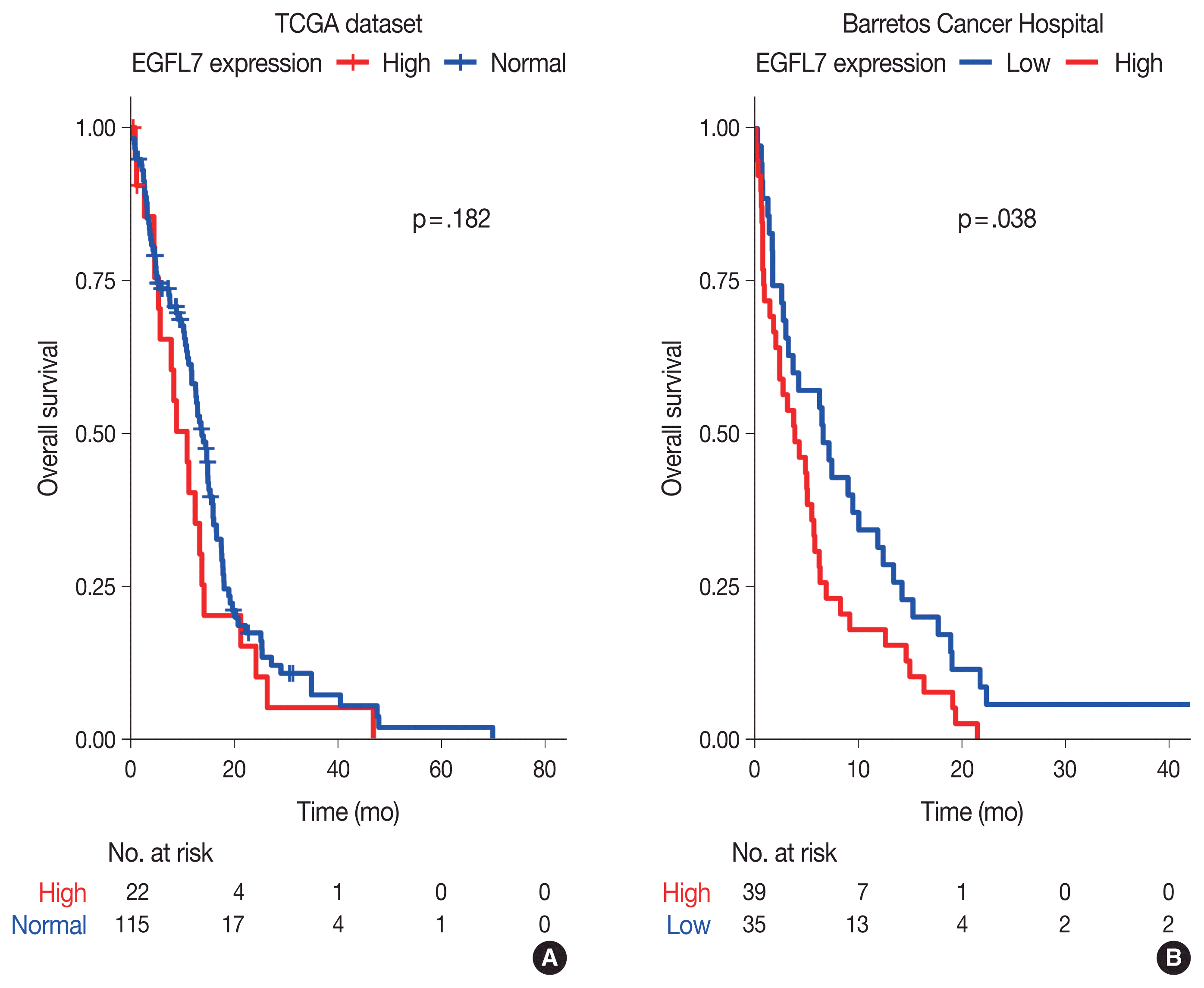
Fig. 1
Kyoto Encyclopedia of Genes and Genomes pathway and gene ontology (GO, biological processes) enrichment analysis of the genes highly correlated to EGFL7 expression. The sizes and color of the bubbles indicate the number of genes per term and significance level. EGFL7, epidermal growth factor-like domain multiple 7.
![]()
Fig. 2
Overall survival curves of glioblastoma patients. (A) In silico data of The Cancer Genome Atlas (TCGA) RNA sequencing. (B) Data of protein expression from 74 patients detected through immunohistochemistry. p-values were calculated using log-rank analysis. EGFL7, epidermal growth factor-like domain multiple 7.
![]()
We further performed analysis of EGFL7 expression by immunohistochemistry in a series of 74 glioblastomas IDH-wildtype. We found high cytoplasmatic EGFL7 expression in 39 cases (52.7%) (shown in Fig. 3). We further associated EGFL7 immunostaining with patients clinicopathological features (Table 1). High EGFL7 immunolabelling was significantly associated with patients presenting KPS lower or equal to 70 (69.2% of the patients with high EGFL7 expression presented KPS ≤ 70 vs. 45.7% of the patients with low EGFL7 expression, p = .041) (Table 1). EGFL7 expression was not associated with the other clinical features analyzed (sex, location, or age group) (Table 1). Importantly, there was found a poorer overall survival of the patients with high EGFL7 expression (mean of 5.69 months) compared to low EGFL7 expression (mean of 9.95 months, p = 0.038, in log-rank analysis, shown in Fig. 2B). Univariate Cox regression showed that patients with high EGFL7 expression have a 1.645-fold risk of death (95% confidence interval, 1.021 to 2.650; p = .041) compared to patients with low EGFL7 expression.
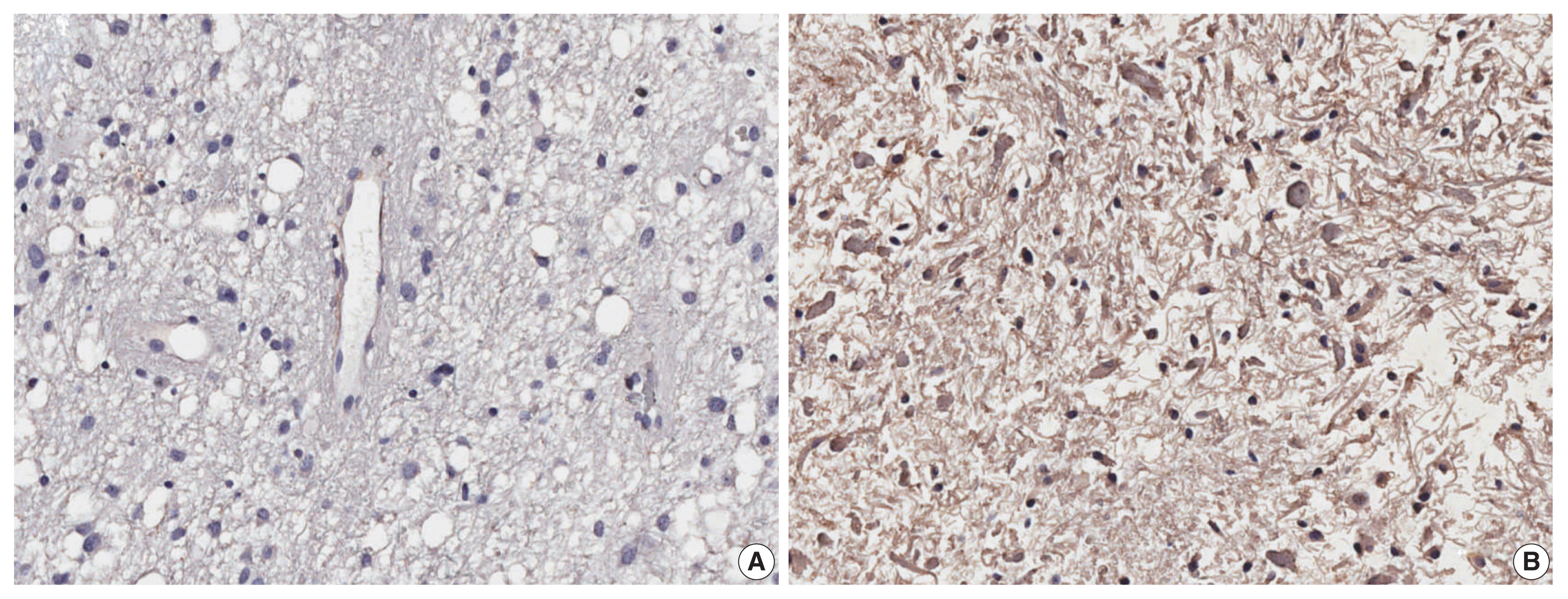
Fig. 3
Microphotographs of resected specimens of glioblastomas immunostained for EGFL7. (A) Absence of immunoreactivity, although vascular endothelial cells serve as a positive internal control for EGFL7. (B) Diffuse and strong EGFL7 immunoreactivity. EGFL7, epidermal growth factor-like domain multiple 7.
![]()
Table 1
EGFL7 expression and its association with clinicopathological data
![]()
DISCUSSION
In the present study, we analyzed the TCGA database in order to infer genes that may participate in the regulation of EGFL7 expression, as well as evaluated whether EGFL7 expression had a prognostic role in glioblastomas. Our analysis found that patients with high EGFL7 expression generally have low KPS and poor survival. Additionally, our findings give insights of the role and regulation of EGFL7 in this tumor type.
EGFL7, a downstream target of BMP9-SMAD1/5-mediated signaling, is a secreted angiogenic factor and promotes endothelial expansion through the interference of Notch signaling, ERK activation, and extracellular matrix remodeling [7]. Data of the literature points to the relation of high expression of EGFL7 and poor prognosis in acute myeloid leukemia (AML) [21,22], hepatocellular carcinoma [23], colorectal cancer [10], pancreatic carcinoma [11], and ovarian epithelial cancer [24]. Wang and colleagues additionally found a poor survival of patients with high EGFL7 expression who were EGFRwt [13] mainly by improving the activity of the β-catenin/TCF complex. Li et al. [25] described that EGFL7 expression promotes the proliferation of hepatocellular carcinoma by increasing CKS2. Corroborating these data, EGFR activation (p-EGFR) was found correlated with EGFL7 overexpression in hormone-producing pituitary adenomas [26].
Our in silico results showed the genes co-expressed with EGFL7 participating mainly in events such as angiogenesis, vasculogenesis, leukocyte transendothelial migration, corroborating the data present in the literature [7,27]. Data also shows enriched genes of Notch pathway (NOTCH4, DLL4, and JAG2). These genes may be subject of further studies to help elucidating the role of this gene in Notch pathway and angiogenesis. In fact, the relation between EGFL7 and Notch pathway remains with conflicting results in the literature. In AML, studies indicate that EGFL7 antagonizes Notch signaling through antagonism to the canonical Notch ligand [28]. On the other hand, silencing of EGFL7 in cutaneous melanoma inactivated Notch pathway, increasing apoptosis and suppressing cell proliferation [29]. Either way, EGFL7 provides its own microenvironment for the migration of endothelial cells, facilitating the budding that leads to angiogenesis [30]. It is known that angiogenesis is one of the hallmarks of cancer, and allows a better supply of blood to the tumor tissue, preventing its cells from having hypoxia/anoxia and/or nutrient deficiency [31].
Parsatuzumab, a humanized anti-EGFL7 antibody, selectively blocks the interaction between EGFL7 and endothelial cells. There is rationale of using this antibody in the inhibition of vascular growth and, used in combination to anti–vascular endothelial growth factor therapy, inhibition of vascular regrowth after vessel damage, which could reduce tumor vascularization [32]. Given the interesting results of the prognostic role of EGFL7, randomized phase II studies used parsatuzumab in combination with FOLFOX or bevacizumab in the treatment of metastatic colorectal cancer [33] and parsatuzumab in combination with carboplatin, paclitaxel, and bevacizumab for first-line nonsquamous non–small cell lung cancer [34] were performed. The authors found no evidence of the effectiveness of combining parsatuzumab with the proposed treatments, but stress the importance of future studies regarding the mechanisms of EGFL7 and development.
EGFL7 expression should be considered beyond vascular background since its expression was found in neuronal components in addition to endothelium [35]. Our immunohistochemistry analysis showed that the patients presenting high EGFL7 expression in tumor cells generally have lower KPS and poorer survival. In line, we have previously shown in pilocytic astrocytoma that high EGFL7 expression was associated with unfavorable outcome [14]. Our in silico analysis pointed to the enrichment of PI3K-Akt and Rap1 signaling pathways. In fact, PI3K-Akt pathway has been extensively described in carcinogenesis, including gliomagenesis. Together with mTOR, this pathway regulates several biological processes that are important in growth, metabolism, angiogenesis, autophagy, and chemotherapy resistance in GBM [36]. Importantly, these pathways have been considered for drug design [36]. On the other hand, Rap1 signaling pathway has been less studied in GBM. Rap1 belongs to Ras-related protein family and is involved in cell adhesion and cell junction formation [37] through ERK/mitogen-activated protein kinase signaling and integrin activation [38]. In vitro, Rap1 was activated by integrin through phospholipase D, enhancing cell adhesion and cell proliferation whereas in vivo Rap1 knockdown profoundly altered U373MG glioblastoma growth in xenograft model [39].
Altogether, our data point to biological functions and pathways related to the increase of tumor vasculature, tumor growth, metabolism, chemotherapy resistance, and changes in cell adhesion and/or cell junction formation. These alterations may be influencing tumor behavior, leading to lower KPS at diagnosis and consequent poor overall survival. However, more molecular studies are warranted in order to elucidate EGFL7 participation in glioblastoma biology.
 XML Download
XML Download