

 PDF
PDF Citation
Citation Print
Print



Introduction
The gross locomotor activities in mice show circadian patterns according to a 12/12-hour dark/light cycle [1]. Nocturnal mice are likely to be less active during the light phase and are more active during the dark phase [1]. Locomotor activities have been widely used to predict sleep-wake states [1-4]. Previous studies have reported that the total sleep duration of mice ranges from 6.2 to 14.9 hours and the awake state occupies 37.93%–74.2% of a 24-hour period [1,2,4].
However, when the rodents were observed more closely, the locomotor activities showed a polyphasic pattern during the active phase beyond their circadian rhythms under the 12/12-hour dark/light entrained condition as well as the free running state [5,6]. The locomotor activity of mice can be expressed depending on their adaptation to the external environment [7,8]. Some studies suggest that the ultradian rhythm of locomotor activities can be related to feeding rhythms [9,10]. Other studies have reported that biological signals, such as body temperature, insulin, and glucocorticoid might be related to the ultradian rhythm of locomotor activities [11,12]. Considering the previous findings, locomotor activities might reflect the adaptation response to the environment or feeding conditions. Furthermore, these ultradian rest-activity rhythms can be reflected by dopaminergic neurotransmission [13,14]. Dopaminergic oscillation is closely related to a hedonic drive and novel seeking [15,16]. Movements to satisfy internal drive or navigate to changing environments might be expressed into gross locomotor activities.
In previous studies, the ultradian rhythm of locomotor activity was analyzed by the observation of peaks and troughs using graphical representations [12,17]. Recently, the ultradian rhythm was analyzed by autocorrelations, spectral analysis, periodograms, or wavelets [5,13,18]. However, there is a lack of studies on the extent to which locomotor activities can be explained statistically by fitted cosine curves based on ultradian rest-activity rhythms. Therefore, this study explored whether the real locomotor activity data in mice could be statistically fitted to the cosine curve within the range of ultradian rhythms, and investigate the amount of locomotor activity that can be explained by the ultradian rest-activity rhythms. In addition, based on the cosine fitting analysis, this study attempted to find and obtain the ultradian rest-activity rhythm of each mouse and estimate their ranges using cosine curves.
Go to : 
Methods
Ethical statements: This study was approved by the Institutional Animal Care and Use Committee of the Pusan National University Hospital (No. PNUH-2017-118).
1. Subjects
Male mice from the Institute of Cancer Research (n=20, weight 34–38 g, age 5–6 weeks) were used in this study. The subjects were acclimated to the new environment for 7 days at a constant room temperature (20°C±1°C) and humidity (40%–60%). The subjects were housed individually in cages. Food and water were provided ad libitum. The dark/light cycle was entrained by a 12/12-hour cycle using a light control system (Philips, Burlington, MA, USA).
2. Experimental procedure and measurement
After the acclimation period, free locomotor activities of each mouse in the cage were observed for 2 days under the 12/12-hour dark/light cycle to compare each ultradian rest-activity rhythm by light entrainment. The locomotor activity of each mouse was measured using infrared motion detectors and the Mlog system (Biobserve Inc., Bonn, Germany). Each detector was placed at the top of the cage. The raw data measured by the Mlog system reflect the intensity of locomotor activity, and a larger value indicates that the animal is more active. This study comprised two experiments that were performed at different times, and the mice were divided into two groups of 10 mice (groups 1 and 2) for each experiment. The data from each experiment were analyzed using the cosine curve fitting. To discover the ultradian rest-activity rhythms, we compared the values analyzed using the data of each mouse and the integrated data of all mice.
3. Individual data analysis
Individual locomotor activity data collected by the Mlog system for 2 days were preprocessed for cosine curve fitting. To reduce data variability with the neighboring data of locomotor activities, data were converted to minutes using an average. Outliers of the data beyond the range of 1.5 × interquartile range (IQR, Q3–Q1) were removed. The data were normalized within the range to obtain values between 0 and 1 (0–1 transformation). Considering that the locomotor activity data were displayed as 0 when there was no movement, the value of the cosine function was also transformed to a value between 0 and 1 (0–1 transformation). The cosine function with the highest fit was calculated repeatedly by changing the period and phase of the cosine function to obtain the best fit, as shown in Fig. 1. The following formula was used for the cosine function in this model:
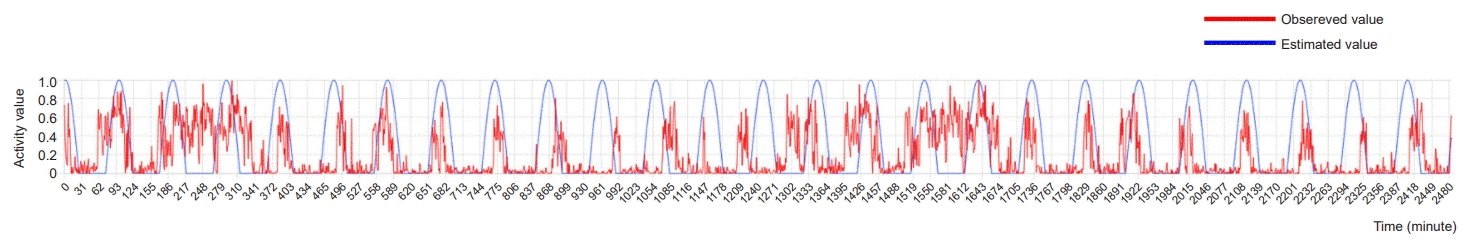 | Fig. 1.An example of cosine fitting analysis using locomotor activities. Individual data analysis was performed with Animal 9 in experimental group 1: y=COS ([x–1.6]/15.3). The locomotor activities of each mouse were fitted into the cosine curve (y=COS [(x-a)/b] +1). The values of a and b were calculated by repeated simulations until the best statistics were obtained. The red line indicates the amount of locomotor activities for 2 days. The blue line shows the cosine curves fitted using the locomotor activities of each mouse.
|
y=COS ([x–a]/b) + 1
where “a” and “b” are arbitrary real numbers for statistical simulation; a and b reflect the phase and amplitude of the cosine curve, respectively. The values of a and b were calculated using repeated simulations, until the best statistics were obtained. For intuitive judgment, it was judged based on the mean absolute error (MAE), which has a relatively small influence on outliers, and MAE values were used to determine the explanatory power of the calculated cosine model in explaining the measured locomotor activities.
4. Statistical analysis of measured data
If it is assumed that individual data can have similar patterns compared with each other, the data measured at the same time are integrated according to each period and converted into one data point for analysis of similarity on the fitted cosine curves. For data integration, mean, median, and max were used as group functions. In the process of integrating the data from the experimental group, the first quartile values sufficiently increased, and the process of transforming values using 0–1 transformation to change the negative value of the cosine function to 0 was unnecessary. Thus, the following formula was used for cosine function:
y=COS ([x–a]/b).
Furthermore, as the data were merged, the y range limit of the cosine function increased from 40 to 100. The rest of the process was the same as that for individual data analysis. Mathematical simulations for cosine curve fitting were performed using R (version 4.1.1.; Foundation for Statistical Computing, Vienna, Austria).
Go to :
Results
When the coefficients were fitted using individual data, as shown in Table 1, the mean MAE was 0.2944, variance was 0.000176, and standard deviation was 0.0132. Although the cosine graph of the fitted function showed a variance from the actual locomotor activities, the cosine functions were well fitted for each data based on the results that the variances of MAE were quite low. The mean MAE value (0.2944) in the data showed that approximately 70% of the fluctuations in locomotor activity were explained by a single cosine function.
Table 1.
Individual statistical data analysis of measured locomotor activities using cosine curve fitting
| Subject | Mean | Max | Min | Q1 | Q3 | SD | VAR | MAEa) | Estimated a | Estimated b | Estimated period (hr) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Experimental group 1 | |||||||||||
| Animal 1 | 1.1879 | 10 | 0 | 0 | 2 | 2.3628 | 5.5831 | 0.3005 | 9.3 | 21.9 | 2.293 |
| Animal 2 | 2.2189 | 17 | 0 | 0 | 3 | 4.0803 | 16.6494 | 0.2741 | 39.9 | 26.8 | 2.806 |
| Animal 3 | 0.8924 | 10 | 0 | 0 | 0 | 2.2141 | 4.9024 | 0.2916 | 0 | 39.8 | 4.168 |
| Animal 4 | 1.0802 | 10 | 0 | 0 | 0 | 2.3513 | 5.5289 | 0.2810 | 0 | 23.7 | 2.482 |
| Animal 5 | 2.7746 | 20 | 0 | 0 | 4 | 4.9152 | 24.1596 | 0.2764 | 29.0 | 19.7 | 2.063 |
| Animal 6 | 0.4770 | 5 | 0 | 0 | 0 | 1.2229 | 1.4955 | 0.2655 | 24.4 | 29.2 | 3.058 |
| Animal 7 | 1.5964 | 12 | 0 | 0 | 2 | 2.9273 | 8.5694 | 0.3031 | 24.5 | 15.4 | 1.613 |
| Animal 8 | 1.2225 | 10 | 0 | 0 | 2 | 2.3943 | 5.7326 | 0.3025 | 0 | 23.4 | 2.450 |
| Animal 9 | 1.2444 | 10 | 0 | 0 | 2 | 2.4576 | 6.0398 | 0.2913 | 1.6 | 15.3 | 1.602 |
| Animal 10 | 1.2014 | 10 | 0 | 0 | 1 | 2.4175 | 5.8445 | 0.2834 | 0 | 30.3 | 3.173 |
| Experimental group 2 | |||||||||||
| Animal 11 | 1.1983 | 10 | 0 | 0 | 2 | 2.3813 | 5.6707 | 0.2934 | 39.9 | 33.6 | 3.519 |
| Animal 12 | 2.1716 | 15 | 0 | 0 | 4 | 3.6793 | 13.5377 | 0.3017 | 39.9 | 16.6 | 1.738 |
| Animal 13 | 1.5758 | 15 | 0 | 0 | 1 | 3.3122 | 10.9710 | 0.3026 | 0 | 39.4 | 4.126 |
| Animal 14 | 1.0208 | 10 | 0 | 0 | 0 | 2.3011 | 5.2954 | 0.2924 | 39.9 | 25.9 | 2.712 |
| Animal 15 | 0.8971 | 10 | 0 | 0 | 0 | 2.1892 | 4.7928 | 0.2871 | 39.9 | 38.7 | 4.053 |
| Animal 16 | 2.3471 | 15 | 0 | 0 | 4 | 3.8717 | 14.9901 | 0.3148 | 0 | 19.9 | 2.084 |
| Animal 17 | 1.2127 | 10 | 0 | 0 | 0 | 2.4627 | 6.0653 | 0.3023 | 0 | 22.9 | 2.398 |
| Animal 18 | 1.2015 | 10 | 0 | 0 | 1 | 2.4119 | 5.8177 | 0.3131 | 0 | 16.8 | 1.759 |
| Animal 19 | 1.1246 | 10 | 0 | 0 | 1 | 2.3053 | 5.3147 | 0.3151 | 7.2 | 16.7 | 1.749 |
| Animal 20 | 1.3212 | 10 | 0 | 0 | 2 | 2.5055 | 6.2778 | 0.2973 | 8.3 | 39.9 | 4.178 |
![]()
Meanwhile, as shown in Table 2, the mean MAE for integrating each individual analysis in the two experimental groups was 0.3284, variance was 0.000566, and standard deviation was 0.0237. The results of the integrated data were inferior to those of individual data analysis. The similarity between the behavior patterns fitted by the cosine curves in the subjects was not high.
Table 2.
Statistics of cosine curve fitting analysis using statistics of measured data
| Integrated data | Mean | Max | Min | Q1 | Q3 | SD | VAR | MAEa) | Estimated a | Estimated b | Estimated period (hr) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Experimental group 1 | |||||||||||
| Mean | 4.7175 | 17 | 0 | 1 | 7 | 3.9671 | 15.7382 | 0.3298 | 17 | 49 | 5.131 |
| Median | 2.1515 | 13 | 0 | 0 | 4 | 3.1019 | 9.6222 | 0.3581 | 48 | 32 | 3.351 |
| Max | 24.6241 | 83 | 0 | 8 | 37 | 19.0927 | 364.5324 | 0.3140 | 24 | 49 | 5.131 |
| Experimental group 2 | |||||||||||
| Mean | 4.7747 | 18 | 0 | 1 | 8 | 4.3435 | 18.8661 | 0.3049 | 5 | 56 | 5.864 |
| Median | 2.7803 | 15 | 0 | 0 | 5 | 3.8083 | 14.5036 | 0.3614 | 91 | 42 | 4.398 |
| Max | 23.9667 | 84 | 0 | 7 | 37 | 19.8799 | 395.2106 | 0.3027 | 10 | 56 | 5.864 |
![]()
The estimated ultradian rest-activity rhythm ranged from 1.602 to 4.168 hours and 1.738 to 4.126 hours in the experimental groups 1 and 2, respectively, as shown in Table 1. The periods of ultradian rest-activity rhythm estimated using integrated data ranged from 3.351 to 5.131 hours and 4.398 to 5.864 hours in the experimental groups 1 and 2, respectively, according to the data integration method in Table 2.
Go to :
Discussion
The results of this analysis suggested the possibility of using a single cosine curve-fitting analysis to explore the ultradian rest-activity rhythm using locomotor activity in mice. In addition, the fitted cosine curve explained approximately 70% of the fluctuating locomotor activities. To compare the estimated ultradian rest-activity rhythm, these analyses were performed for the locomotor activities of the mice that were acclimated under well-controlled light conditions, including an illumination system with the 12/12-hour dark/light cycle. Therefore, it should be cautiously considered that these influences may be different under irregular or disrupted conditions undergoing a major change. Nevertheless, it is expected that locomotor activity in mice can be predicted to a large extent under the influence of the internal ultradian rest-activity rhythms of the individual in well-controlled stable environments.
However, the graph of the fitted function differed from the actual locomotor activity. Based on the MAE, the errors ranged from 26.7% to 31.5%. These differences may be caused by irregular locomotor activities due to adaptive responses to external environments [8,17]. Based on these findings, approximately 70% of the mouse's movement information may be affected by the internal ultradian oscillating rest-activity rhythms and 30% may be affected by external events or environment. In addition, some studies have reported that ultradian rest-activity rhythms may be reflected by the state of dopaminergic neurotransmission [13,14]. Meanwhile, dopaminergic neurotransmission has been related to addiction and depression [19,20]. Research on the relationship between addiction and depression with ultradian rhythms using cosine fitting analysis might be facilitated.
Meanwhile, the dopaminergic state can be influenced by various factors, such as genetic components, dopaminergic medications, and substances [13,14,21]. Considering the changeability of the dopaminergic state, these intrinsic ultradian oscillators may also change during long observation periods. Because fitting was attempted under the assumption that the ultradian rhythm will have a constant cycle, there is the possibility that these results may not be accurate in longitudinal observations. In case of the analysis of ultradian rhythms, wavelet analysis that can evaluate the changeable period of ultradian rhythms according to time would be useful [22].
The fitted cosine curves for the locomotor activities of each mouse were different with a period range from 1.602 to 4.168 hours, yet some mice showed similar movement patterns. The inter-mice differences were also supported by the finding that the MAE values after the integration of data in the experimental groups increased compared to the values before integration. These findings suggest the possibility that each individual has its own rest-activity rhythm depending on their own biological states or genetic characteristics. In this study, the period for cosine analysis of locomotor activity was not determined, and any period was allowed to be fitted within 2 days. As such, if the data strongly suggested a circadian rhythm (24-hour period), the circadian rhythm would be estimated as the best rhythm. However, in this study, cosine curves with a period ranging from approximately 1.6 to 4.2 hours were optimally fitted. These results suggest that the locomotor activity data reflect the ultradian rhythm better than the circadian rhythm; that is, even though the locomotor activities are influenced by circadian rhythms, they reflect the ultradian rhythm better. Previous studies have implied that the patterns of locomotor activities show an ultradian rhythm of several hours reflecting dopaminergic oscillation [13,14]. In studies using spectral analysis, the ultradian rhythms ranged from 2 to 6 hours, similar to the results of this study [14]. Thus, considering the findings of this study, cosine fitting analysis using locomotor activities can be useful in evaluating ultradian rest-activity rhythms.
This study had some limitations. First, because this study analyzed the locomotor activities of mice under relatively stable conditions, the results of this study cannot be generalized to the analysis of locomotor activities under every condition, including major stressful events or disrupting conditions. Second, cosine fitting analysis cannot be used with data that were observed for long-term periods, which cannot be assumed to have constant ultradian rhythms. Third, this study did not examine other supporting biological conditions that may affect movement. Fourth, the results of this study cannot be extended to female mice, because only male mice were included. Considering the sex difference between male and female mice, it is necessary to investigate whether these results are reproduced in female mice in the future study. However, this study suggests an appropriate analytical method for exploring ultradian rest-activity rhythms using locomotor activity in mice.
In conclusion, this study suggests that locomotor activities can be statistically fitted into a single cosine curve under well-controlled conditions with light entrainment of the 12/12-hour dark/light cycle. Each mouse might have its own ultradian rest-activity rhythm related to the biological conditions, such as the state of dopaminergic ultradian oscillation, as reported in previous studies [13,14]. In addition, this statistical analysis model using single cosine fitting analysis can be effectively utilized in research on ultradian rest-activity rhythms. As such, this ultradian rest-activity model using locomotor activity may be used to investigate the effects of any pharmacological agent on ultradian rhythms. In the future, it is necessary to explore whether this cosine-fitting analysis can be used to analyze ultradian rhythms under different experimental conditions.
Go to :
 XML Download
XML Download