

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Parkinson’s disease (PD) is a neurodegenerative disorder characterized by the marked degeneration of substantia nigral dopamine neurons. High frequency stimulation of the subthalamic nucleus (STN) has become an established treatment of the motor symptoms of PD to reduce L-3,4-dihydroxyphenylalanine (L-DOPA) induced motor side effects13,24).
However, the exact mechanism of deep brain stimulation (DBS) in the STN is unknown, and it is a subject of ongoing research. At the cellular level, DBS has complex effects by driving or inhibiting neuronal elements depending upon the location and stimulation parameters2,15,23).
Glutamate, gamma-aminobutyric acid (GABA), and dopamine are the principal neurotransmitters in the basal ganglia circuit. Although there have been many reports about the changes in extracellular striatal dopamine from high frequency stimulation (HFS)-STN, there are only a few number of papers about extracellular striatal glutamate, GABA changes, and tyrosine hydroxylase (TH) immunoreactivity of HFS-STN1,12,17).
The study of the changes of extracellular striatal glutamate and GABA after HFS-STN may contribute to the understanding of the role of glutamate and GABA on striatal and nigral dopamine system.
Glutamate and GABA play an important role in basal ganglia activity and modulate the striatal dopaminergic functions. In this study, we compared the effects of stimulation of the subthalamic nucleus on extracellular glutamate and GABA levels in the striatum in sham group, 6-hydroxydopamine (6-OHDA) lesioned group, and 6-OHDA lesioned plus DBS group, respectively.
Go to : 
MATERIALS AND METHODS
Animals
The subjects of this study were 8 weeks-old adult male albino Sprague-Dawley rats (Orient Animal Corp., Gyungki-do, Korea) weighing 280–300 g each. They were group-housed with three per cage, under a reversed 12: 12 h light-dark cycle with light on from 08: 00–20: 00 h. All of the rats had free access to food and water in the barrier system with regulated temperature (23±1°C), humidity (50±1°C), and ventilation (9–12 times per hour). All animals were handled daily for at least 7 days prior to the experiment. The care and treatment of the animals were provided in accordance to the Guide for the Care and Use of Laboratory Animals (NIH). All animal experiments were approved by the Institutional Animal Care and Use Committee (IACUC) of the Catholic University. All experiments were carried out in accordance with the Guidelines for Animal Experimentation.
Experimental design
The rats were divided into three groups (n=6 per group): 1) sham group (no-treatment), 2) 6-OHDA-lesioned group, and 3) 6-OHDA-lesioned plus DBS group. To assess the efficacy of the 6-OHDA lesion, which induces Parkinsonian-like motor deficits, all rats were tested for apomorphine (Sigma-Aldrich Inc., St. Louis, MO, USA)-induced (0.05 mg/kg, s.c) rotational behavior at 21 days after the exposure to 6-OHDA. The results were expressed as ipsilateral turns/min. The rats exhibiting a vigorous rotational response (>100 total turns in a period of 45 minutes) to apomorphine (0.05 mg/kg, s.c.) were selected for further study (Fig. 1). It has previously been demonstrated that the rats meeting this criterion have greater than 95% depletion of striatal dopamine 19).
 | Fig. 1Apomorphine rotation test. This graph showed that the mean and 95% conÿdence interval in 6-OHDA group (134.3, 114.80 to 153.86) and 6-OHDA plus DBS group (127.66, 101.98 to 153.35). There was no statistical signiÿcance of radiation turns between two groups (p=0.6069). 6-OHDA: 6-hydroxydopamine, DBS: deep brain stimulation.
|
6-OHDA lesions
For medial forebrain bundle (MFB) lesioning, all of the rats were anesthetized with sodium pentobarbital (60 mg/kg, i.p.) and secured in Kopf stereotaxic apparatus (Phymep, Paris, France). We treated 12 rats with the injections of 3 ug of 6-hydroxydopamine (6-OHDA; Sigma-Aldrich, USA; 8 ug/2 uL in 0.02% ascorbic acid in normal saline, Sweden AB) into the right MFB, at the flow rate of 2 uL/min. The stereotaxic coordinates of the injection site at Bregma were: anteroposterior, −4.4 mm; lateral, −1.2 mm; and dorsoventral, −7.8 mm. All stereotaxic co-ordinates cited here are according to the stereotaxic atlas of Paxinos20). The rats were kept warm after the injections and were allowed to recover from anesthesia. They were returned to the animal facility for 3 weeks to allow the degeneration of dopaminergic neurons induced by the neurotoxin. After they stabilized, they were processed for microdialysis experiments.
Surgery and microdialysis procedures
The rats were anesthetized with sodium pentobarbital (50 mg/kg, i.p.). Using aseptic techniques, a guide cannula (CMA/12, CMA Microdialysis, Solna, Sweden) aimed to terminate in the dorsal striatum (anteroposterior, 1.0 mm; lateral, −3.2 mm and dorsoventral, −6.0 mm to Bregma) was stereotaxically implanted and attached to the skull using skull screws and dental cement. The cannula was then closed with a tight-fitting stainless-steel obturator. The microdialysis was performed with a 4 mm microdialysis probe (CMA/12, CMA Microdialysis), connected to a syringe pump via dual liquid swivel, that was inserted into the guide cannula which is then perfused with an artificial cerebrospinal fluid (145 mM NaCl, 2.7 mM KCl, 1.2 mM CaCl2, 1.0 mM MgCl2, 2.0 mM Na2HPO4; pH 7.4) at a constant rate of 1.0 mL/min. Dialysate samples were collected during 20 minutes of sampling intervals via outlet tubing connected to a microfraction collector (CMA Microdialysis). The position of the probes was verified by the histological examination at the end of the experiments. The rats suffering from bleeding was excluded from the analysis.
STN stimulation
Concentric bipolar platinum/iridium stimulating electrodes (style CB-BFE75, FHC, Inc., Bowdoin, ME, USA) were used. The stereotaxic co-ordinates of the stimulation site (STN) at Bregma were: anteroposterior, −3.6 mm; lateral, −2.5 mm; and dorsoventral, −8.0 mm. Stimuli were delivered for 20 minutes, with a programmable pulse generator (Iso-Flex/Master-8; AMPI, Israel) and a stimulus isolation unit. Stimulation parameters corresponded to those used in the clinical practice (frequency 130 Hz, 60 us rectangular pulse width and 200 uA intensity). At the end of each experiment, a mechanical lesion created by stimulating electrode in the STN, so that the position of the electrode could be checked at the end of experiments.
Analytical procedures
The dialysate (injection volume 30 μL) was assayed for GABA and glutamate using high performance liquid chromatography (HPLC) coupled to a fluorescence detector. The detector was equipped with a high performance analytical cell (ESA model 5014), which is tailored for use in microdialysis applications. The mobile phase was composed of 75 mM monobasic sodium phosphate, 0.1 mM ethylenediaminetetraacetic acid, 1.4 mM octanesulfonic acid, and 10% acetonitrile, adjusted to pH 3.2 with HPLC grade phosphoric acid. The separation of monoamine metabolites was performed on Waters Nova-Pak C-18 column (4 mm, 150×3.9 mm). The flow rate of the system was 0.8 mL/min. GABA and glutamate in dialysates were expressed as percentages of three baseline samples collected immediately before DBS.
Tyrosine hydroxylase (TH) immunohistochemistry
After the microdialysis processes, all of the rats were deeply anesthetized with sodium pentobarbital (80 mg/kg, i.p.). The rats were intracardially perfused with 100 mL of saline followed by 500 mL of 4% paraformaldehyde in 0.1 M phosphate buffered saline (PBS) (pH 7.0). The brains were removed, postfixed overnight, and cryoprotected in 20% sucrose in 0.1 M PBS at 4°C. The brains were cut via cryostat sectioning into 30 um coronal sections at the level of the striatum and substantia nigra (Leica, Germany). The tissue was stored at −20°C for histochemistry and these slices were histochemically processed as free-floating sections.
The brain sections were washed in PBS containing 0.3% Triton X-100. The primary mouse monoclonal antibodies against tyrosine hydroxylase were used (TH, concentration 1: 2000; Zymed Laboratories INC., San Francisco, CA, USA). The primary antibodies were diluted with blocking solution (10% bovine serum in phosphate buffered saline, pH 7.4) and the tissues were incubated for overnight at 4°C with constant agitation. After rinsing in PBS, the sections were incubated for 2 hours at room temperature in biotinylated anti-mouse (Santa Cruz Biotechnology Inc., San Francisco, CA, USA) diluted 200X in phosphate buffered saline tween and 2% normal goat blocking serum. The sections were placed in the mouse ABC staining system (Santa Cruz Biotechnology Inc.) for 2 hours at room temperature. Following further rinsing with PBS, the tissue was developed using diaminobenzadine (Sigma-Aldrich Inc.) as the chromogen with nickel intensification. The sections were mounted on slides, air-dried, and cover slipped for microscopic observation.
Statistical analysis
The results were analyzed using repeated measures analysis of variance (ANOVA) for multiple comparisons where necessary as means±SE (standard error). Repeated measures ANOVA was used to determine the significance of any changes in the HPLC analysis of GABA and glutamate results and in histochemical data, among the different groups; this was followed by the Tukey technique. The criterion for statistical significance was considered as p values <0.05.
Go to :
RESULTS
Effect of STN-HFS on extracellular glutamate levels in the striatum of sham, 6-OHDA group, and 6-OHDA plus DBS group
As illustrated in Fig. 2, the average and SE of baseline glutamate level in the striatum corresponding to 100% was 543±72 nM (sham group), 491±87 nM (6-OHDA group) and 632±83 nM (6-OHDA plus group), respectively. There is a statistical significance in three groups (F2, 15=7.641, p<0.01). Post-hoc comparisons indicated that the striatal glutamate contents in the 6-OHDA plus DBS group were significantly high compared to the sham (p<0.01) and 6-OHDA group (p<0.01). Post-hoc comparisons also indicated that the striatal glutamate contents in the sham group were significantly high compared to the 6-OHDA group (p<0.05). HPLC analysis demonstrated a tendency of an increase in extracellular striatal glutamate during DBS and a decrease after DBS. The maximal peak of extracellular glutamate (110±8%) is shown at 20 minutes after DBS start time in 6-OHDA plus DBS group. The concentration of extracellular glutamate progressively declined to basal value in 20 minutes after DBS. However, it showed a tendency of maintaining the extracellular glutamate to high concentration state even after DBS, in the 6-OHDA plus DBS group compared to the 6-OHDA group.
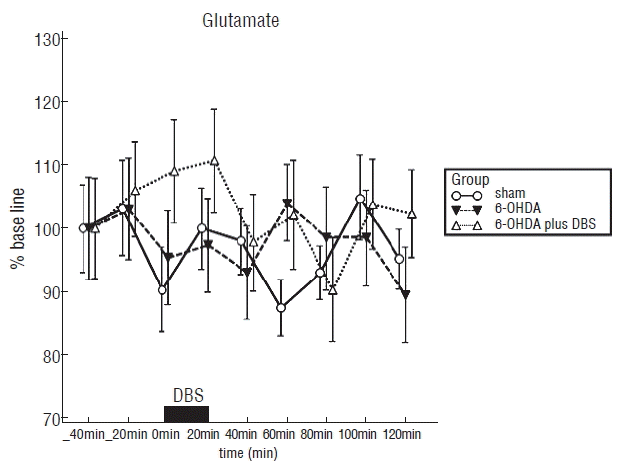 | Fig. 2E°ect of high frequency stimulation of subthalamic nucleus on extracellular glutamate in the striatum measured by microdialysis in rats. Each point represents the mean value±standard error from six rats per group: normal rats without deep brain stimulation (sham group), 6-OHDA-lesioned rats without deep brain stimulation group (6-OHDA group), 6-OHDA-lesioned rats with deep brain stimulation (6-OHDA plus DBS group). 6-OHDA: 6-hydroxydopamine, DBS: deep brain stimulation.
|
The effect of STN-HFS on extracellular GABA levels in the striatum of sham, 6-OHDA group, and 6-OHDA plus DBS group
As illustrated in Fig. 3, the average and SE of baseline GABA level in the striatum corresponding to 100% was 67±15 nM (sham group), 48±19 nM (6-OHDA group) and 75±17 nM (6-OHDA plus group), respectively. There is a statistical significance in the three groups (F2, 15=5.826, p<0.05). Post-hoc comparisons indicated that the striatal GABA contents in the 6-OHDA plus DBS group were significantly high compared to the 6-OHDA group (p<0.01). However, there was no statistical significance of GABA contents between the sham and 6-OHDA plus DBS group (p>0.05). Post-hoc comparisons also indicated that the striatal GABA contents in the sham group were significantly high compared to the 6-OHDA group (p<0.01). HPLC analysis demonstrated a tendency of an increase in extracellular striatal GABA during DBS and a decrease after DBS. The maximal peak of extracellular GABA (114±6%) is shown at 40 minutes after DBS start time, in 6-OHDA plus DBS group. The concentration of extracellular GABA gradually declined to basal value in 40 minutes after DBS. However, it showed the tendency of maintaining the extracellular GABA to high concentration state even after DBS, in the 6-OHDA plus DBS group compared to the 6-OHDA group.
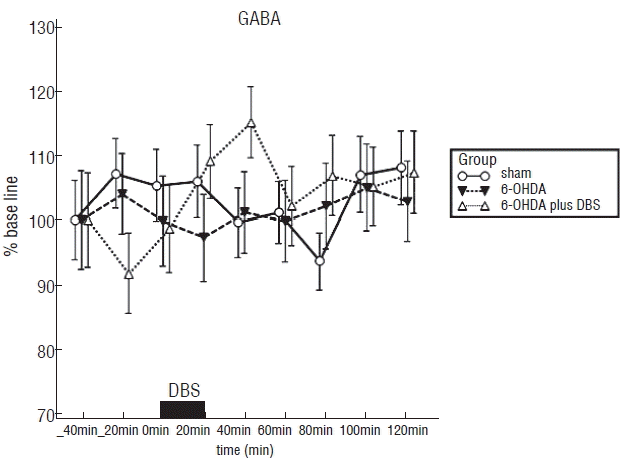 | Fig. 3Effect of high frequency stimulation of subthalamic nucleus on extracellular GABA in the striatum measured by microdialysis in rats. Each point represents the mean value±standard error from six rats per group: normal rats without deep brain stimulation (sham group), 6-OHDA-lesioned rats without deep brain stimulation group (6-OHDA group), 6-OHDA-lesioned rats with deep brain stimulation (6-OHDA plus DBS group). GABA: gamma-aminobutyric acid, 6-OHDA: 6-hydroxydopamine, DBS: deep brain stimulation.
|
Histological controls
At the end of the experiment, histologic evaluation of the location of stimulating electrode tip in STN was examined by cresyl violet stained section (Fig. 4). As illustrated in Fig. 5, dopamine depletion was severe in the substantia nigra and striatum of 6-OHDA group. Optical density measurement of TH immunoreactivity in the substantia nigra and striatum in 6-OHDA group showed a mean depletion of 90% compared to the unlesioned side.
 | Fig. 4Cresyl violet stained photomicrographs showing the sites of deep brain stimulation in the subthalamic nucleus. Scale bar: 250 um. STN: subthalamic nucleus.
|
 | Fig. 5Photographs of coronal rat brain sections at nigral and striatal levels. Sham group: tyrosine hydroxylase (TH) immunostaining at nigral and striatal levels has no di°erence between ipsilateral and contralateral side. 6-OHDA group: TH immunostaining at nigra and striatum in ipsilateral side to 6-OHDA lesioned site shows remarkable decrease compared with contralateral side. 6-OHDA plus DBS group: TH immunostaining at nigra and striatum in ipsilateral side to 6-OHDA lesioned site shows mild increase compared with the ipsilateral side in 6-OHDA group. Scale bar=500 um. 6-OHDA: 6-hydroxydopamine, DBS: deep brain stimulation, ST: striatum, SN: substantia nigra, VT: ventral tegmental area.
|
Go to :
DISCUSSION
HFS of the STN is a potent tool for treatment of Parkinson disease (PD). It has been reported to result in clinical improvement in PD patients and animal models, through reducing levodopa requirements and levodopa-induced dyskinesia10,14,21). However, the mechanisms by which STN-HFS affects the basal ganglia output remain unclear.
The STN plays a key role in controlling the output nuclei of the basal ganglia, the substantia nigra pars reticulata (SNr), and the internal segment of the globus pallidus (GPi). It plays an important role in controlling the movement, so it is a key structure in the indirect pathway of basal ganglia circuit18). It receives its main inputs from the cortex, thalamus, and brainstem through its glutamatergic projections exciting the GABAergic neurons of the SNr and GPi, which consequently reinforce the inhibitory effects of the basal ganglia on the thalamic and brainstem premotor networks5).
In the basal ganglia model, PD bradykinesia results from disequilibrium between indirect and direct pathways and an overactivity of the STN and the GPi6).
In terms of principal neurotransmitters in basal ganglia circuit, glutamate is a major excitatory neurotransmitter, GABA is major inhibitory neurotransmitter, and dopamine has excitatory or inhibitory role that depends on the situation.
It was known that the basal ganglia PD model showed reduced excitatory input from the cortex to striatum, reduced inhibitory output (direct pathway) from striatum to GPi, and increased inhibitory output (indirect pathway) from striatum to external segment of globus pallidus (GPe) and STN6,26).
Our result demonstrates that there is statistical significance of extracellular glutamate and GABA contents among the sham, 6-OHDA, and 6-OHDA plus DBS group. HPLC analysis showed a tendency to decrease extracellular glutamate and GABA of the striatum in the 6-OHDA group compared to the sham group. These findings were consistent with those of the intact and PD basal ganglia model. In other words, the 6-OHDA group represented reduced inhibitory input (indirect pathway) and reduced excitatory input (direct pathway) from SNc to striatum.
There is one interesting result in HPLC result on 6-OHDA group and 6-OHDA plus DBS group. The initial extracellular GABA and glutamate contents before HFS-STN differed between the 6-OHDA group and 6-OHDA plus DBS group. It may be reasonable to assume that the cause of this difference is due to microlesioning effect of subthalamic nucleus, which is a clinical effect similar to lesion provoked by the insertion of the electrode itself without stimulation9). We speculate that microlesioning effect of subthalamic nucleus contributed to the increase of extracellular glutamate and GABA in the 6-OHDA plus DBS group before stimulation. It is important to rule out the microlesioning effect of subthalamic nucleus to examine accurately the effect of stimulation. However, the duration of microlesioning effect of subthalamic nucleus remains unknown. We inserted stimulation electrode and microdialysis cannula at the same time, because it was technically very difficult to insert them at different times. This is a limitation of our study and we could not rule out the microlesioning effect of subthalamic nucleus.
HPLC analysis showed a tendency of extracellular glutamate and GABA contents in striatum increasing during DBS and decreasing after DBS in 6-OHDA plus DBS group, compared to sham group and 6-OHDA group. These findings suggest that HFS-STN may influence extracellular glutamate and GABA contents in the striatum. There are questions regarding the way in which HFS-STN is able to increase extracellular glutamate and GABA contents in the striatum.
In terms of glutamate, it has a role of excitatory input to striatum from cortex and SNc D2 receptor, which is reduced in the PD basal ganglia model compared to the intact basal ganglia model19,20). We speculate that HFS-STN might contribute to the enhanced reduction of excitatory input and antidromic activation of hyper-direct pathway between STN and cortex25). Conclusively, the increase of extracellular glutamate contents in striatum during DBS showed the enhanced reduction of excitatory input to striatum.
In terms of GABA, it has a role of inhibitory input to striatum from SNc D1 receptor, which is reduced in the PD basal ganglia model compared to the intact basal ganglia model19,20). Striatal GABA increase could be related to the glutamate increase provoked by the STN-HFS. The interaction between GABA and glutamate is supported by the study of Segovia et al showing glutamate increases striatal GABA from stimulation of GABA interneurons22). Conclusively, the increase of extracellular GABA contents in striatum during DBS showed the enhanced reduction of inhibitory input to striatum.
Our results may be similar to results of Bruet et al3). However, there are some differences between our results and their results. First, target sites of 6-OHDA lesion are different in modelling Parkinson’s disease in rat. Our target site is MFB, while their target site is SNc. MFB lesions are almost complete and very few dopaminergic neurons in the SNc compared to SNc lesions4,7). We think that MFB lesions are adequate to the advanced Parkinson’s disease. Second, microlesion effect of STN is a unique feature of our results. Third, statistical methods of analyzing neurotransmitter change by HFS-STN are different. We used repeated measures ANOVA, while they used Mann-Whitney U test. We focused on the aspect of neurotransmitter change on consideration of time variable in both inter-group and within-group by HFS-STN.
Finally, we provided the evidence that HFS-STN may have an important role in enhancing the reduction of direct and indirect pathway inputs from SNc to striatum. We suggest that the reinforcement of the effects of inputs from direct and indirect pathways of SNc to striatum can be mediated by HFS-STN.
These data may elucidate functional aspects of the basal ganglia network involved in the reduction of motor impairments in PD patients undergoing this type of electrical stimulation. It is also expected that these data will be useful in the research of Parkinson’s disease dementia, because pyramidal neurons in the hippocampus contain glutamatergic and GABAergic synapses16).
It is not clear if the STN stimulation significantly influences the striatal dopamine system9–11). The changes of glutamate and GABA in striatum by HFS-STN may be inferred to be correlated to the striatal dopamine system. Our histology control data demonstrated the increased optical density of TH immunoactivity in the 6-OHDA plus DBS group compared to 6-OHDA group. This suggests that STN-HFS may promote TH recovery in both striatum and substantia nigra. According to other studies, the pathophysiology of this finding might be showing the increase in metabolic activity of dopamine neurons and the compensation for the loss of dopamine neuron8,27).
The resulting functional STN-HFS effect in the striatum is illustrated by an increase of striatal glutamate and GABA in both 6-OHDA plus DBS group, suggesting that the results of STN-HFS are not restricted to the direct STN targets but are more involved in the widespread adaptive changes within the BG.
The main limitations of our study include as follows: the stimulation of STN during anesthetic state, which was done on the rats not freely moving; short-term and only one time stimulation of STN, which is not a chronic stimulation; and the insufficiency of STN stimulation for sham group and in post-stimulation behavioral study.
Go to :
 XML Download
XML Download