

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Multiple cranial burr hole (MCBH) surgery is one of the indirect bypass surgeries used in moyamoya disease (MMD), which is performed only or additionally to other direct or indirect surgeries. Several authors reported the effectiveness of this method, but the treatment mechanism is still uncertain929315056). Usually it seems to be thought that the mechanism is to make simple pathways for extracranial vessels to grow into the ischemic brain.
Recently there were some reports that provided us a clue for thinking a different mechanism. Kim et al.34) reported that angiogenesis of the lower extremities could be improved by multiple burr holes on tibia via autologous transplantation of bone marrow stem cells in Buerger's disease. Furlanetti et al.14) reported a successful treatment on a child with total scalp avulsion by multiple burr hole trephination and skin grafting. They showed abundant granulation tissues growing out from the multiple cranial burr holes in his report. Crain et al.7) reported that transplanted bone marrow generates new neurons in the human brain. Iwashita et al.21) reported that the cranial bone flap contains enough bone marrow cells for autologous blood stem cell rescue.
Based on these reports, we hypothesized that calvarial bone marrow stem cells migrate into the ischemic brain via multiple burr holes and affect angiogenesis and neurogenesis in MMD (Fig. 1). For the verification of this hypothesis, demonstrations of following four postulates were needed : 1) existence of migrant and proliferating cells in the ischemic cortex around the burr hole, 2) bone marrow origin of the proliferating cells, 3) increased angiogenesis, and 4) neurogenesis around the burr hole.
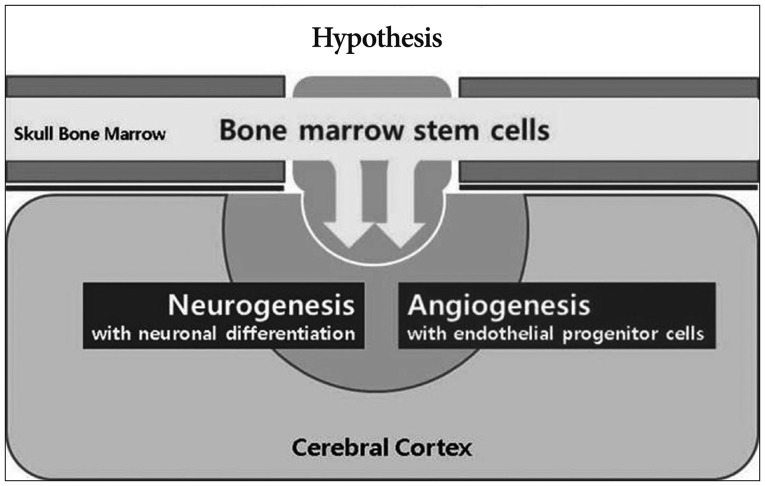
The purpose of this study is investigating the role of a burr hole and calvarial bone marrow-derived stem cells (BMSCs) in ischemic rat brain model to ascertain our hypothesis.
Go to : 
MATERIALS AND METHODS
This study used adult male Sprague-Dawley (SD) rats (Central Lab. Animal Inc., Seoul, Korea) weighing 200-250 g. All procedures were performed in accordance with the guidelines for care and use of laboratory animals approved by Chung-Ang University's Institutional Animal Care and Use Committee. Twenty SD rats were evenly divided into four groups : normal control group (n=5), burr hole group (n=5), ischemia group (n=5), and ischemia+ burr hole group (n=5). In the burr hole group, the rats underwent burr hole trephination and received a small cortical incision at the left parietal area without cerebral infarction. In the ischemia group, focal cerebral infarction was induced in the left frontoparietal area without a cranial burr hole or cortical incision. The ischemia+burr hole group underwent focal cerebral infarction, burr hole trephination, and a small cortical incision in the same area.
Focal cerebral infarction
Focal cerebral infarction was induced at the frontoparietal area by transient middle cerebral artery occlusion (MCAO) as described previously40). Briefly, under isoflurane anesthesia (Choongwae, Seoul, Korea), midline incision on anterior neck and exposure of common carotid artery (CCA), internal carotid artery (ICA) and external carotid artery (ECA) were performed. A 3-0 monofilament nylon suture with a rounded tip was introduced through the left ECA and was advanced to the ICA until slight resistance was felt. After 1 hour of occlusion, the suture was removed to restore blood flow, and the wound was sutured. The rat was allowed to recover from the anesthesia and was returned to its cage with free access to food and water.
Burr hole trephination with minimal cortical incision
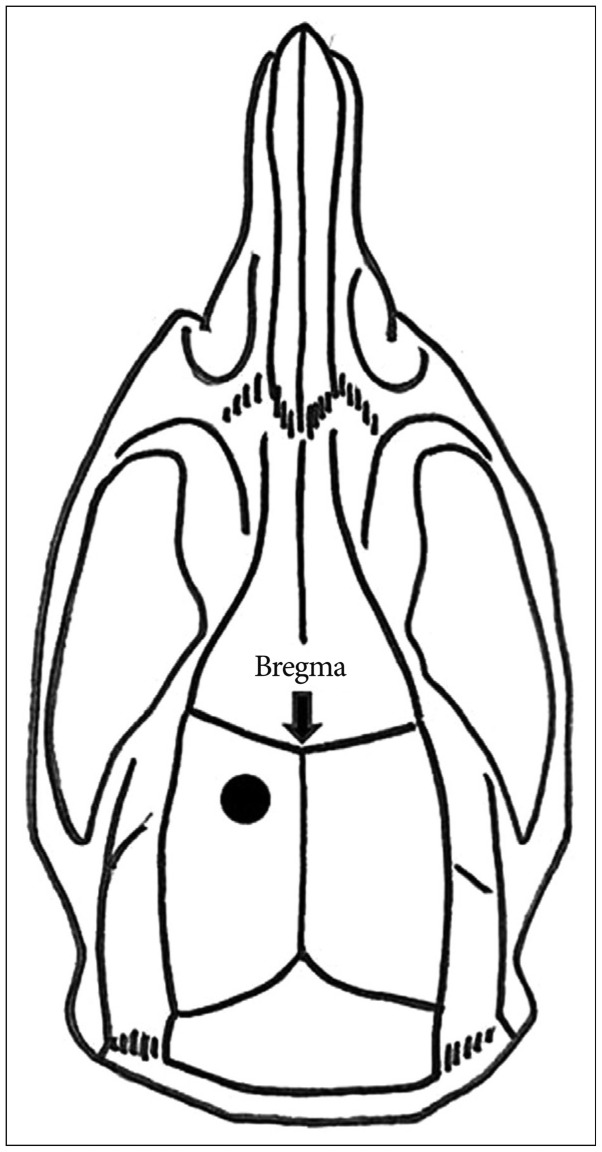
A burr hole was made seven days after the ischemic injury. Each rat in the burr hole group and the ischemia+burr hole group was placed in an induction cage with 4% isoflurane and 3 L/min of O2 for several minutes before being transferred to the operation table, where its head was fixed in a stereotaxic device (David Kopf Instruments, Tujunga, CA, USA). Inhalation anesthesia was maintained with 2-3% isoflurane, 1.5 L/min of N2O, and 1.5 L/min of O2. After a 1.5-cm midline incision was made on the scalp, burr hole trephination (diameter : 2 mm) was performed with an electric drill (Ideal Micro-Drill™, Cellpoint Scientific Inc., Gaithersburg, USA) in the left parietal area, 3 mm lateral and 3 mm posterior to the bregma (Fig. 2). Great care was taken to avoid damaging the underlying dura mater during drilling and removal of the cranial bone. Finally, a small incision was made at the dura and superficial cerebral cortex with a 26-gauge needle, similar to the usual techniques for MCBH surgery for MMD.
BrdU injection and sacrifice
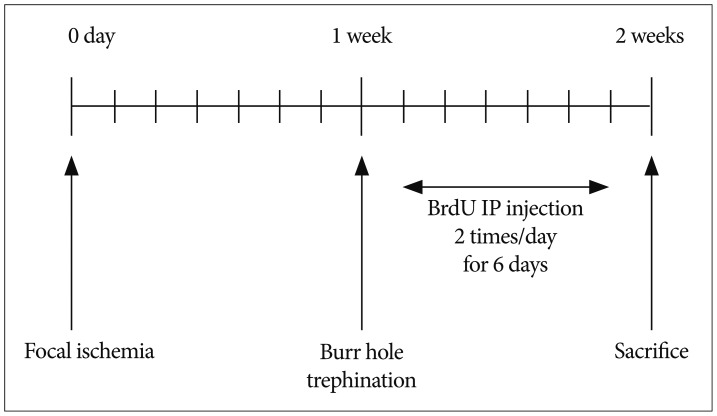
To detect proliferating cells, cell proliferation-specific marker 5-bromodeoxyuridine (BrdU; Sigma, 50 mg/kg dissolved in saline for each injection) was injected intraperitoneally, 2 times a day for 6 days beginning on the day after burr hole trephination. All rats in each group were injected with BrdU intraperitoneally according to the same schedule (Fig. 3). Seven days after burrhole trephination, the rats were sacrificed, and their brains were perfused with approximately 150 mL of 0.1-M phosphate-buffered saline (PBS) at a rate of 52 mL/min to clear the blood, followed by 450 mL of 4% paraformaldehyde.
Immunohistochemical analysis
The paraffin-embedded brain was cut into 5-mm-thick coronal sections with brain matrix (Ted Pella Inc., Redding, CA, USA). A series of adjacent 5-µm-thick sections were cut from each block in the coronal plane. Brain sections were deparaffinized in xylene and dehydrated through a graded ethanol series. Endogenous peroxidase activity was quenched by 30 minutes incubation in a freshly prepared 3% H2O2-methanol solution. Antigen retrieval was performed by boiling the brain sections in 0.1 mol/L citrate buffer (pH 6.0) in a microwave oven (650 to 720 W) for 8 minutes three times with 5 minutes cooling intervals at room temperature. After washing with 0.01 mol/L PBS buffer (pH 7.4), the sections were incubated with 20% normal horse serum (Vector Laboratories, Burlingame, CA, USA)-PBS for 30 minutes. After excessive normal serum was blotted from the sections, mouse anti-BrdU antibody (Biocare Medical, CA, USA) diluted at 1 : 100 in PBS buffer was added, and the slides were incubated at 4℃ overnight. After washing with 0.01 mol/L PBS buffer, the sections were incubated with 1 : 200 biotinylated horse anti-mouse Immunoglobulin G (rat absorbed, Vector Laboratories, Burlingame, CA, USA)-PBS containing 1.5% normal horse serum at room temperature for 30 minutes. After washing with 0.01 mol/L PBS buffer, the sections were incubated with an Elite Vectastain avidin-biotin complex (ABC)-peroxidase kit (Vector) and stained with diaminobenzidine (DAB; Sigma-Aldrich Co., St. Louis, USA).
The same staining procedures were then repeated with CD34 (1 : 100 rat monoclonal antibody, GeneTex, USA), vascular endothelial growth factor (VEGF) (1 : 200 rabbit polyclonal antibody, Abcam, Cambridge, UK), Nestin (1 : 100 rat monoclonal antibody, Santa Cruz, CA, USA), and DCX (doublecortin) (1 : 100 rabbit polyclonal antibody, Abcam, Cambridge, UK).
Go to :
RESULTS
Existence of proliferating cells
BrdU is a synthetic nucleoside that is an analogue of thymidine, a specific marker for cells that have proliferated after BrdU administration52). In the focal ischemia+burr hole group, numerous BrdU (+) cells were found in the cerebral cortex under the burr hole (Fig. 4); these cells were located at the surface of the cortical incision site and not in the white matter. In the burr hole group, a much smaller number of BrdU (+) cells were observed (Fig. 4). In the normal control and focal ischemia groups, no BrdU (+) cells were detected in the corresponding area (Fig. 4). Therefore, the proliferating cells appeared to come through the burr hole and were induced by cerebral infarction.
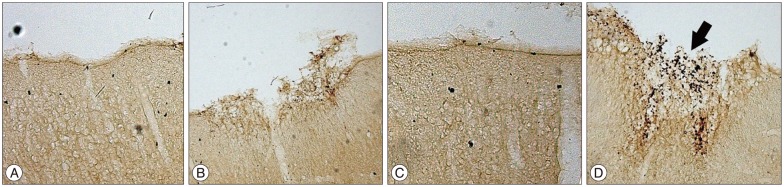 | Fig. 4Immunohistochemical stain with BrdU. A : Normal control group. B : Burr hole group. C : Ischemia group. D : Ischemia+burr hole group. In the ischemia+burr hole group, BrdU (+) cells (arrow) were found at the cortical injury site beneath the burr hole in the coronal section. In the burr hole group, a few BrdU (+) cells were detected. Original magnification ×200. BrdU : 5-bromodeoxyuridine.
|
Origin of the proliferating cells
CD34 molecule is a cell surface glycoprotein that functions as a cell-cell adhesion factor. It is also known as a marker protein for the mesenchymal stem cells of bone marrow17). In the focal ischemia+burr hole group, CD34 (+) cells were found at the cortical incision site under the burr hole, in the same area as the BrdU (+) cells (Fig. 5). In this group, BrdU (+) or CD34 (+) cells were found mainly around the cortical injury area. This finding suggests that these cells originated from the bone marrow, BMSCs, rather than endogenous migration from the subventricular zone (SVZ).
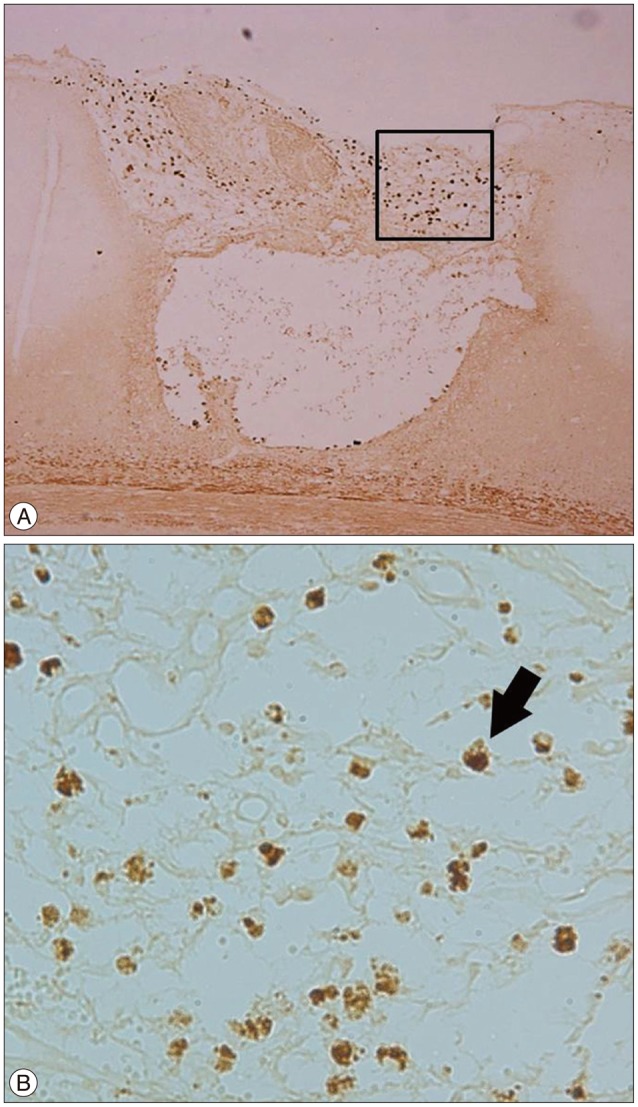
Angiogenesis
VEGF is a subfamily of growth factors-specifically the platelet-derived growth factor family of cystine-knot growth factors. The normal function of VEGF is to create new blood vessels during embryonic development, new blood vessels after injury, muscle following exercise, and new vessels (collateral circulation) to bypass blocked vessels12). In the focal ischemia+burr hole group, VEGF (+) cells increased around the cortical incision site, the same area where the increased number of BrdU (+) or CD34 (+) cells were detected (Fig. 6). The increase in VEGF expression and CD34 (+) cells in the same area is suggestive of increased angiogenesis at the ischemic cerebral parenchyma around the burr hole by BMSCs.
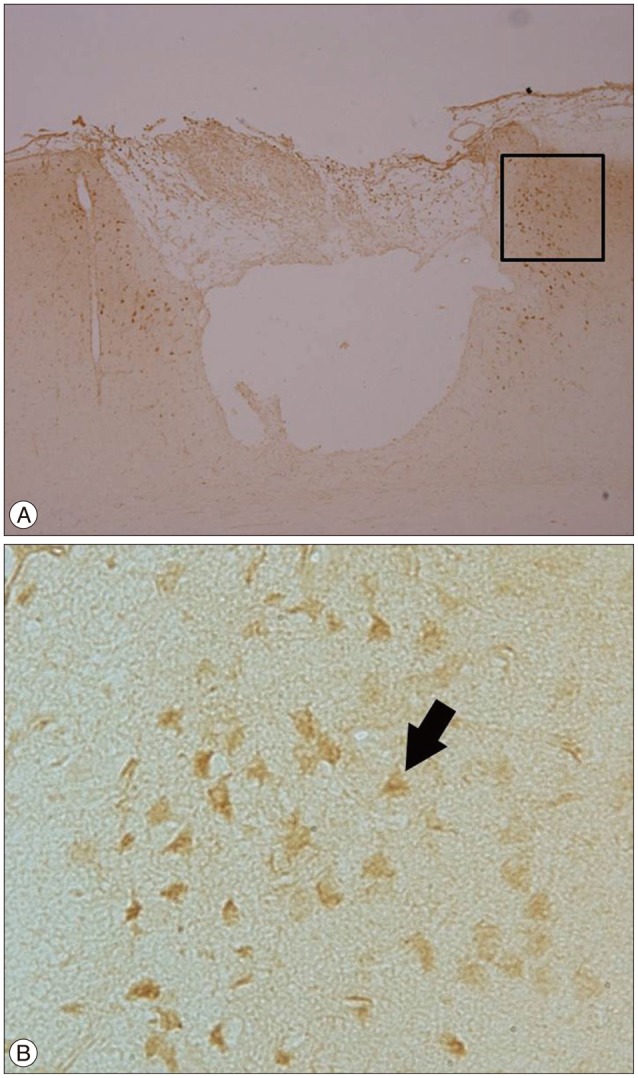 | Fig. 6Immunohistochemical stain with VEGF in the ischemia+burr hole group. Many VEGF (+) cells (arrow) were found under the burr hole. A : Original magnification ×100. VEGF (+) cells markedly increased at the cerebral parenchyma adjacent to the cortical incision site below the burr hole. B : Black rectangle in A, original magnification ×400. VEGF : vascular endothelial growth factor.
|
Neurogenesis
DCX is a microtubule-associated protein expressed by neuronal precursor cells and a marker for immature neurons in embryonic and adult brains. This protein is considered the "gold standard" for the measurement of neurogenesis due to the exclusive expression of DCX in developing neurons6). Nestin, a protein marker for neural stem cells and their immediate or differentiated progeny, is an intermediate filament protein expressed in dividing cells during the early stages of development in the central nervous system41). These two markers indicate neurogenesis. DCX (+) cells were found at the cortical incision site under the burr hole, where the BrdU (+) cells and CD34 (+) cells were also found (Fig. 7). In addition, the Nestin (+) cells were found in the same area as the BrdU (+) cells and CD34 (+) cells (Fig. 8). There was no DCX or Nestin (+) cells in the deep white matter around the burr hole and cortical incision site. The expression of these two markers in the same area as the CD34 (+) cells without any connection to the SVZ suggests that increased neurogenesis might be related to the BMSCs from the burr hole.
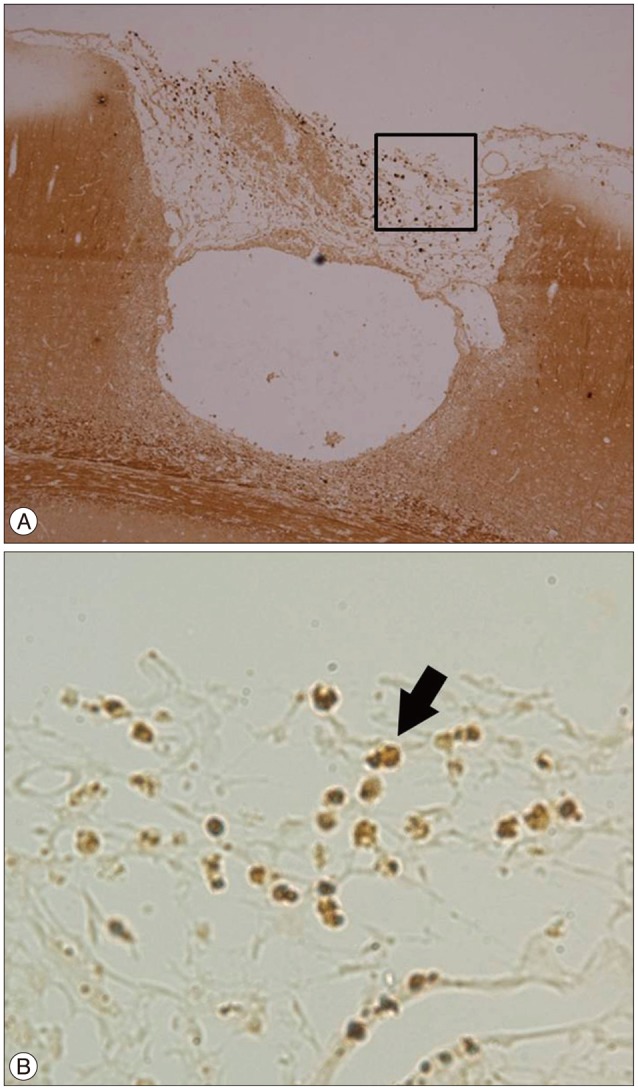
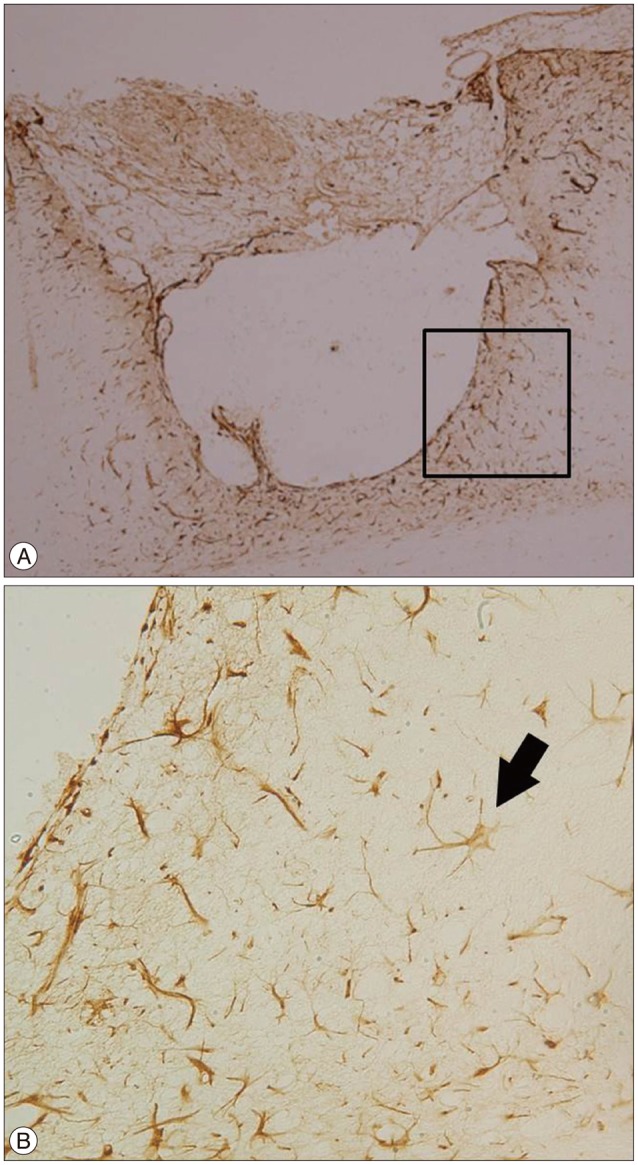
Go to :
DISCUSSION
MCBH surgery for MMD
Several surgical techniques have been developed to prevent recurrent cerebral ischemia in patients with MMD. These techniques fall into two basic categories : direct bypass surgery, such as anastomosis of the arteries between extracranial (superficial temporal artery, STA) and intracranial (middle cerebral artery, MCA) circulations203343); and indirect bypass surgery, in which the STA or vascularized tissues, such as the temporalis muscle, dura mater, and omentum, are placed directly on the cerebral cortex to enhance collateral formation. Indirect bypass surgeries are technically simple and have been widely used. These surgeries include encephaloduroarteriosynangiosis (EDAS)48), encephalomyoarteriosynangiosis (EMAS)51), and simple burr holes without vessel synangiosis (MCBH surgery)929).
MCBH surgery is a simple, safe, and effective procedure for revascularization of the ischemic territory914262829303145). Because MBCH surgery implies no injury to the branches of the ECA, it may be performed as an adjunct to other direct or indirect bypass surgeries. Neovascularization and the disappearance of moyamoya vessels occur after MCBH surgery, as in other indirect surgeries45). Endo et al.9), Kawaguchi et al.29), Kawamoto et al.31), and Suzuki et al.50) reported its effectiveness for pediatric cases of definite MMD, and Kawaguchi et al.27) and Yanakawa et al.56) used it for adult cases of definite MMD. Recently, arachnoid opening has been recommended during MCBH surgery21113).
However, the mechanism for the efficacy of MCBH surgery in MMD is still in question, and no previous studies attempted to identify it. Most authors have assumed that the burr holes function as simple pathways between the ischemic brain and the extracranial tissue richly supplied by extracranial circulations (galea, temporal muscle, and dura)914262829303145). Considering that increased angiogenesis has been shown to occur with multiple drilling at the tibia in Buerger's disease34), and that highly-vascularized tissues have been shown to grow out from multiple cranial burr holes in total scalp avulsion injury14), it seems probable that our hypothesis is one of the treatment mechanisms of MCBH surgery for MMD.
Origin of proliferating cells
The proliferating cells observed in this study could have originated from three areas : the SVZ (endogenous migration); the circulating blood, including endothelial progenitor cells (EPCs) (hematogenous immigration); or the bone marrow of the skull (direct infiltration).
Adult neurogenesis in the SVZ and subgranular zone (SGZ) of the hippocampus was established in the late 1990s11542). It appears to be limited to a few brain regions : the SVZ, SGZ, and posterior periventricular (PPv) area1). Recent studies have found that brain ischemia significantly upregulates adult neurogenesis in the SVZ and SGZ44). In addition, NPCs in the SVZ have been shown to migrate to ischemic regions, where they appear to form proper neuronal subtypes to replace damaged neurons in the striatum and cortex23). However, we could not find any progenitor cells in endogenous migration courses, such as the pathway along the cortico-callosal border corresponding to the lateral migratory stream and the white matter around the cortical incision and burr hole. For this reason, the possibility of endogenous migration seems very low.
Humans have two types of bone marrow : red marrow and yellow marrow. Red marrow consists of hematopoietic stem cells and mesenchymal stem cells, EPCs, and very small embryonic-like stem cells. Red marrow is located in flat bones, such as the pelvis, sternum, cranium, rib, vertebrae, scapulae, and epiphyseal ends of long bones. Yellow marrow consists of fat cells and is located in the medullary cavity of long bones. EPCs circulate in the blood and have the ability to differentiate into endothelial cells that make up the lining of blood vessels. Adult EPCs were first characterized in 1997 by purification of CD34 (+) hematopoietic progenitor cells in vitro3). These cells expressed various endothelial markers and incorporated into neovessels at the site of ischemia18). EPCs play a role in tumor growth, metastasis, and angiogenesis and are considered novel therapeutic targets for cancer therapy. In this study, CD34 (+) cells were found only at the site of the burr hole in the ischemia+ burr hole group. Although this finding does not exclude the possibility of hematogenous migration of cells, the existence of CD34 (+) cells (BMSCs) under the burr hole suggests the origin of the BMSCs is the cranial bone marrow.
It has been shown that highly-vascularized tissues can "grow out" from cranial burr holes during treatment for total scalp avulsion14). Thus, it is possible that these tissues can "grow-in" through burr holes after MCBH surgery for MMD. This led us to hypothesize that BrdU (+) or CD34 (+) cells migrate from the cranial bone marrow.
Angiogenesis
Embryologically, vasculogenesis is mediated by angioblasts during the fetal period, but it is mediated by bone-marrow-derived endothelial progenitor cells in adulthood3). Unlike vasculogenesis in the fetal period, adult angiogenesis involves external factors, such as hypoxia, neoplasm, and VEGF expression1849).
Ischemic heart diseases and ischemic limb circulatory disorders can be treated by increasing angiogenesis at the ischemic sites. Therefore, recent progress in stem cell research has led to laboratory and clinical applications involving stem cell transplantation to accelerate angiogenesis42536). These approaches are based on the ability of stem cells to increase the expression of VEGF in ischemic tissues. From a therapeutic point of view, MMD is similar to ischemic heart disease and ischemic limb circulatory disorders. MMD does not require neuronal regeneration, but instead, angiogenesis.
Our data suggest that angiogenesis results from the migration of VEGF-expressing CD34 (+) EPCs into the ischemic cortex through a burr hole.
Neurogenesis
Under physiological conditions, neurogenesis occurs exclusively within the dentate gyrus of the hippocampus and olfactory bulb in normal adult mammalian brains, including humans810). Altered environments brought about by active exercise3253), learning16), seizure46), aging37), or global cerebral ischemia39)) can enhance neurogenesis.
Since the discovery that neurons are differentiated from adult BMSCs in humans and rats, there has been remarkable progress in the use of BMSCs for various neurological diseases55). Theoretically, stem cell transplantation is necessary to achieve therapeutic effects by way of neuroregeneration in various central nervous system diseases, including cerebral infarction. Little is known about the functional status of these newly generated neurons, although Bang et al.5) found that bone marrow stem cells can lead to limited clinical improvement after acute cerebral infarction.
In the ischemia+burr hole group, DCX (+) cells or Nestin (+) cells increased in the ischemic brain under the burr hole. In the other groups, these cells could not be detected. Although Jiang et al.22) reported that cortical neurogenesis occurs in the cerebral cortex of adult rats after temporal MCAO, the distribution pattern of BrdU (+) cells was random throughout cortical layers II through IV, with the highest density in the ischemic boundary zone, which differs from the results of our study. Therefore, it is possible that the neuroprogenitors observed in our study might have originated from the cranial bone marrow and migrated through the burr hole.
Clinical applications
Recently, several techniques have been developed to accelerate the expression of VEGF in ischemic tissues, including direct injection of VEGF protein19), gene therapy with a vehicle such as a plasmid and virus3854), cell therapy with EPCs or BMSCs45), or combined therapy with two or more of the above techniques. With direct protein injection, however, the protein has a short half-life, so this technique requires a relatively high protein concentration and repeated procedures, which increases the possibility of side effects47). In addition, the safety of gene therapy with a vehicle has not been fully demonstrated for direct application to patients35). Although transplantation of autologous BMSCs into the corresponding muscle during encephalomyosynangiosis operation has been reported35), direct transplantation of autologous BMSCs does not yet appear acceptable.
Nonetheless, the results of this study suggest that specific techniques can be used to improve angiogenesis and neurogenesis after MCBH surgery or other indirect revascularization surgeries for MMD. First, minimizing the use of bone wax at the burr-hole surface may improve migration of calvarial BMSCs into the ischemic brain. Second, additional decortication at the inner rim of the burr hole may enlarge the exposed bone marrow to the ischemic cortex, thus enhancing migration of BMSCs. Third, fenestration of the dura matter and arachnoid membrane may eliminate the barrier to migration of BMSCs into the brain. Fourth, administration of exogenous cytokine (granulocyte macrophage-colony stimulating factor) may enhance the proliferation and mobilization of calvarial BMSCs. Fifth, filling the burr hole with a material that prevents fibrosis or adhesion may increase the duration of BMSC migration.
Limitations
This study has some limitations. First, although some studies have attempted to establish an experimental animal model for MMD24), there is currently no established model for this condition. We used a focal ischemia model, MCAO, but its pathophysiology is different from that of moyamoya disease. Second, our findings may have been influenced by traumatic brain injury due to the cortical incision, and this could have affected the stem cells activities. Third, in the absence of confocal microscopic evidence with double and triple fluorescent stainings, the existence of BrdU (+), CD34 (+), DCX (+), or Nestin (+) cells in the same area does not guarantee that all of the cells are the same BMSCs that originated from the burr hole. Fourth, hematogenous immigration could also have occurred and requires further study.
Go to :
CONCLUSION
In conclusion, BMSCs exist at the cortical incision site below a burr hole in ischemic rat brain, and angiogenesis and neurogenesis increased at the same area. These stem cells seem to come from the cranial bone marrow through a burr hole. Based on these findings, we would like to suggest that increased angiogenesis and neurogenesis by calvarial BMSCs through burr holes may play some important role in the MCBH surgery for MMD.
Go to :
 XML Download
XML Download