

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Glioblastoma (GBM) is one of the most common primary brain tumors in the central nervous system in adults, accounting for approximately 40% of all brain tumors5). Currently, temozolomide (TMZ)-based chemotherapy with concomitant radiotherapy is accepted as the first line therapy for malignant gliomas10). However, despite aggressive treatment, most patients with gliomas have a median survival time of about one year after diagnosis. The unsatisfactory response to TMZ treatment is related to the DNA repair protein, O6-methylguanine-DNA methyltransferase (MGMT). Accordingly, many studies have reported the use of TMZ in combination with other therapies to modify the expression of MGMT, including histone deacetylase inhibitors such as vorinostat and valproic acid9). Additionally, interferon-beta (IFN-β) has been reported in a phase I clinical trial in combination with TMZ11).
IFN-β is known to act as a drug sensitizer and is widely used in combination with other antitumor agents7). The combination of IFN-β and TMZ has shown significant synergistic antitumor effects in human glioma xenograft models. IFN-β is known to inactivate the expression of MGMT via p53 gene induction, thus it enhances the therapeutic efficacy of TMZ8). The results of the clinical trial using the combined treatment of IFN-β and TMZ showed that the median overall survival of the combination group was significantly greater (19.9 months) compared to the TMZ alone group (12.7 months) by about 7 months11).
Although the current study also utilizes the combined treatment of IFN-β and TMZ, our study is unique in that we focused on delivering the therapeutic agents to the tumor. Specifically, we used mesenchymal stem cells (MSCs) as a porter. MSCs have a pathotropism for human gliomas after intravascular or local delivery6). Therefore, considering that the poor survival of patients with human malignant gliomas is related, at least in part, to the inability to deliver therapeutic agents to the tumor, the tropism of human MSCs for malignant gliomas can be exploited for therapeutic purposes, i.e., MSCs can be used as delivery vehicles with tumor-targeting capabilities.
In the present study, we engineered human MSCs to secret mouse IFN-β (MSC-IFN-β) and investigated the combination effects of MSC-IFN-β and TMZ using in vitro and in vivo experiments.
MATERIALS AND METHODS
Cell culture and reagents
Human bone marrow-derived MSCs (Catholic MASTER Cells) were obtained from the Catholic Institute of Cell Therapy (Seoul, Korea). Cells were thawed, and the culture process was performed according to the manufacturer's instructions. Human bone marrow aspirates were obtained from the iliac crest of healthy donors aged 20 to 55 years after approval by the Institutional Review Board of Seoul St. Mary's Hospital (approval numbers KIRB-00344-009 and KIRB-00362-006). Bone marrow aspirates from each consented donor were sent to the GMP-compliant facility of the Catholic Institute of Cell Therapy (Seoul, Korea, http://www.cic.re.kr) for the isolation, expansion, and quality control of human bone marrow-derived MSCs. Cells were plated in a culture dish and cultured with human bone marrow-derived MSC basal medium supplemented with MSC growth supplement at 37℃ with 5% CO2. The MSC growth medium was used for all cell expansion procedures, unless otherwise mentioned. During cell expansion, cells were tested for bacterial sterility, mycoplasmal sterility, and the endotoxin level (<3 EU/mL).
Adenovirus infection to drive IFN-β expression
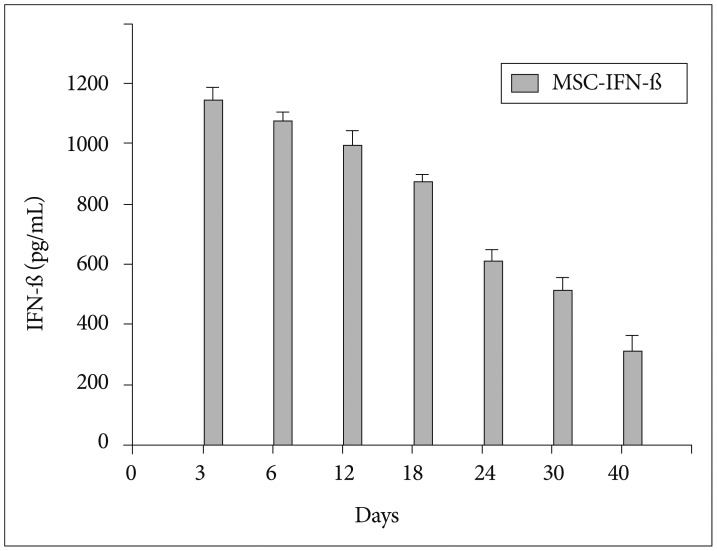
The recombinant adenoviral vector encoding the gene for enhanced green fluorescent protein (Ad-GFP) and mouse IFN-β (Ad-IFN-β) was constructed and produced using the AdEasy vector system, following the manufacturer's instructions (Quantum Biotechnologies, Carlsbad, CA, USA)3). MSCs were infected with a multiplicity of infection of 50 for Ad-GFP or Ad-IFN-β. Enzyme-linked immunosorbent assays (ELISA) was done to confirm the transgene expression of MSC-GFP and MSC-IFN-β. The IFN-β proteins were checked in both supernatants (Fig. 1) and the supernatants at day 3 after infection were acquired for in vitro experiments.
Cell line
The GBM cell line GL26 was obtained from American Type Culture Collection and cultured in Dulbecco's modified Eagle's medium (Invitrogen, Grand Island, NY, USA). All media was supplemented with 2 mmol/L L-glutamate, 100 U/mL penicillin, 100 µg/mL streptomycin, and 10% fetal bovine serum purchased from Invitrogen. Cells were incubated at 37℃ in a humidified atmosphere containing 5% CO2/95% air.
Antitumor effect of TMZ and IFN-β in glioma cells
To analyze the antitumor effects of TMZ and IFN-β in the GL26 glioma tumor cell line, cells were washed twice with the medium and incubated with medium containing MSC-GFP (control), medium containing MSC-IFN-β, or medium containing MSC-IFN-β with 20 µM TMZ. After exposure to the various conditions, cells were detached by trypsinization and the viable cell population was determined using 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)1).
Animal experiments (tumor size and survival evaluation)
The established human glioma tumor cell line, GL26, was used in this experiment. C57BL/6 mice (6-8 weeks old; Harlan-Sprague-Dawley) were used in accordance with institutional guidelines under the approved protocols. For the intracranial implantation of syngeneic GL26 tumor cells in the brains of C57BL/6 mice, animals were stereotactically inoculated with 1×105 GL26 cells [in 3 µL of phosphate-buffered saline (PBS)]into the right frontal lobe (2 mm lateral and 1 mm anterior to bregma, at a depth of 2.5 mm from the skull base) using a guide-screw system (Hamilton syringe, Hamilton Company, Reno, NV, USA). To increase the uniformity of the xenograft take and growth, cells were injected simultaneously using a multiport Microinfusion Syringe Pump (Harvard Apparatus, Holliston, MA, USA). Animals were anesthetized with xylazine/ketamine during the procedure.
For the survival experiment, intracranial glioma-bearing mice were randomly divided into four groups (n=5 in each groups) after tumor implantation and treated with intratumoral injections of saline (PBS), TMZ alone, MSCs infected with Ad-IFN-β (MSC-IFN-β), or a combination of TMZ with MSC-IFN-β. TMZ was injected intraperitoneally and MSC-IFN-β was injected intracranially, directly to the tumor. The therapeutic treatments were administered on the 14th day after tumor inoculation. The brains from the treated mice (n=5/groups) were serially sectioned (20 µm-thich sections, obtained every 200 µm into the tumor) and then stained with hematoxylin and eosin. Tumor size was determined as described previously4). The section with the maximum tumor area was calculated via a computer using MetaMorph software (Molecular Devices, Sunnyvale, CA, USA).
Evaluation of apoptosis by immunohistochemistry
Apoptotic cells were visualized using a terminal deoxynucleotidyltransferased UTP nick end labeling (TUNEL) assay kit (Roche, Basel, Switzerland) developed using Cy3-conjugated streptavidin (Jackson ImmunoResearch Laboratories, West Grove, PA, USA). Endogenous peroxidase activity was blocked by 3% H2O2 for 10 min at room temperature. After washing with PBS, 50 µL of TUNEL reaction mixture was pipetted onto the sections, which were then incubated in a humidified chamber at 37℃ for 1 h. The reaction was stopped by adding wash buffer. In all sections, nuclei were stained with 4,6-diamidino-2-phenylindole (DAPI) (Sigma-Aldrich, St. Louis, MO, USA) for counterstaining. Apoptotic cells were also measured using a computer with the MetaMorphsoftware (Molecular Devices, Silicon Valley, CA, USA).
Statistical analysis
All data are expressed as the mean±the standard error of the mean. All statistical comparisons between the groups were determined using one-way analyses of variance and Student's t-tests. Survival analyses were conducted by a log-rank test based on the Kaplan-Meier method. Probability values of <0.05 were considered statistically significant.
RESULTS
Combined antitumor effect of MSC-IFN-β and TMZ in human glioma cells
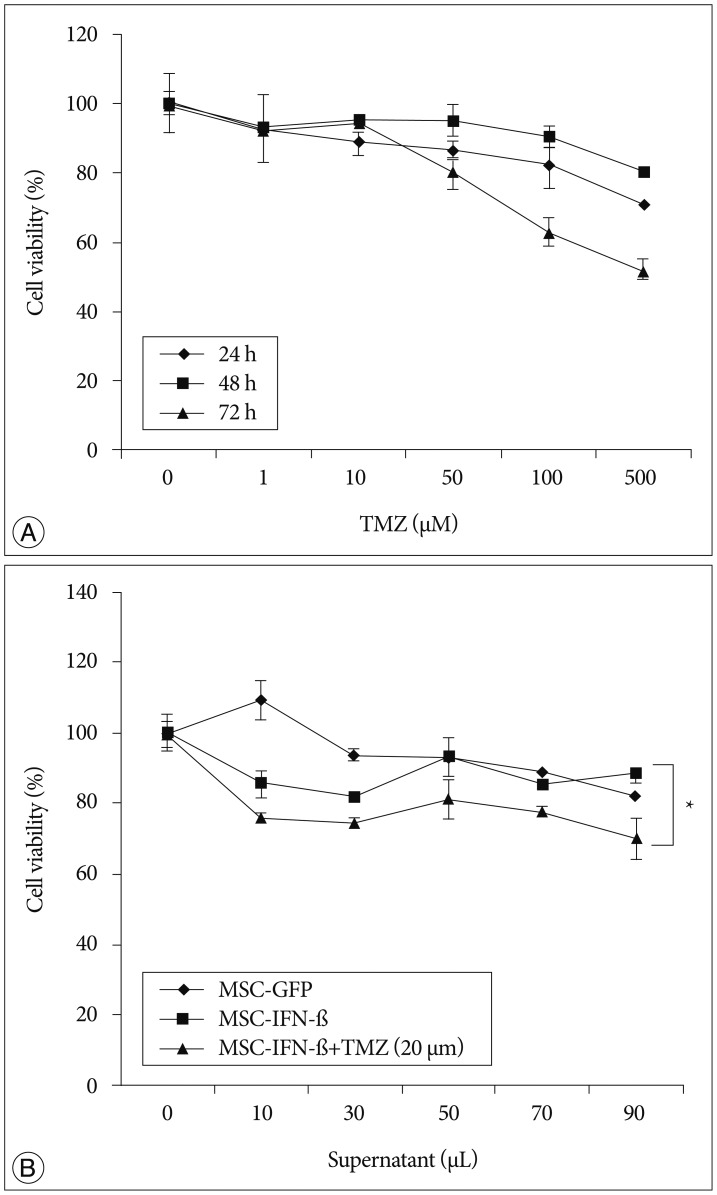
To confirm the transgene expression of MSC-IFN-β, the IFN-β protein was checked in supernatants from MSC-IFN-β by enzyme-linked immunosorbent assays (Fig. 1). IFN-β protein production was well detected in the supernatants from IFN-β-transduced MSCs. To examine the involvement of MGMT in the GL26 cell line, the expression of the MGMT was analyzed by western blot and reverse transcription (RT)-polymerase chain reaction (PCR). As a result, MGMT was not detected in both examinations (data not shown). As GL26 cells were sensitive to temozolomide, to determine the combination effect with MSC-IFN-β clearly, a temozolomide concentration of 20 µM was chosen as a subtoxic dose for the following in vitro combination experiments (Fig. 2A).
To assess the combined antitumor effects of MSC-IFN-β and TMZ in the GL26 cell line, cells were incubated in culture medium containing 0-90 µL MSC-IFN-β supernatant and/or 20 µM TMZ for 72 h. Subsequently, the number of viable cells was analyzed via MTT assay. As shown in Fig. 2B, a significantly enhanced antitumor effect was observed for the combined treatment of MSC-IFN-β and TMZ (p<0.005, combined treatment vs. each single treatment). This result suggests that the combination of MSC-IFN-β and TMZ has combined antitumor effects in the glioma cell line.
Effects of the MSC-IFN-β and TMZ combination on tumor growth and the survival of glioma-bearing mice
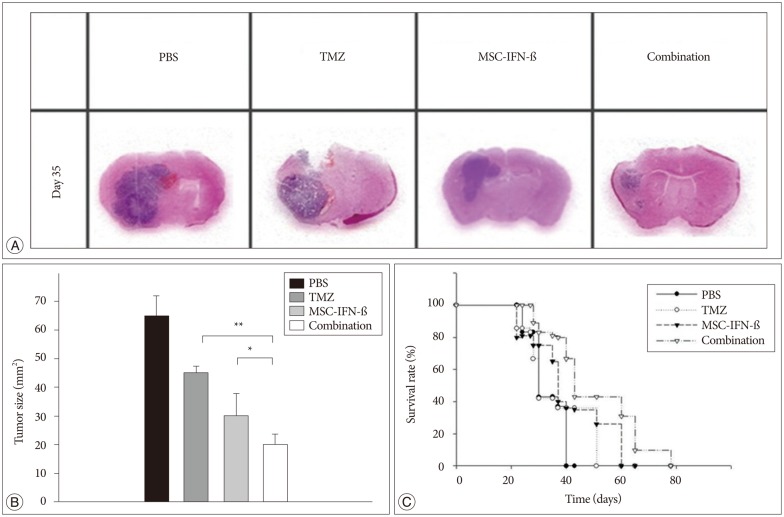
To determine whether the MSC-IFN-β treatment alone and combined with TMZ showed antitumor effects on gliomas in vivo, glioma-bearing mice were treated with MSC-IFN-β, TMZ, or both. Tumor sizes were verified by histological staining (Fig. 3A). The average tumor area in each treatment group was decreased compared with PBS-treated mice. Additionally, as shown in Fig. 3A, the tumor sizes were mostly decreased in the combined treatment group. Histological analyses were performed for each treatment group, and the combined treatment was significantly better than MSC-IFN-β or TMZ alone (p<0.05, MSC-IFN-β vs. combination; p<0.01, TMZ vs. combination) (Fig. 3B). These results show that MSC-IFN-β and TMZ alone reduced the rate of tumor growth in glioma-bearing mice, while the combination of MSC-IFN-β and TMZ, had significantly better antitumor effects. Next, survival analyses were conducted, as shown in the survival curve of glioma-bearing mice (Fig. 3C). The survival of mice in the combined treatment group was significantly prolonged compared with the survival in each of the monotherapy groups (p<0.05).
Effects of the combined MSC-IFN-β and TMZ treatment on apoptotic cell death in intracranial glioma
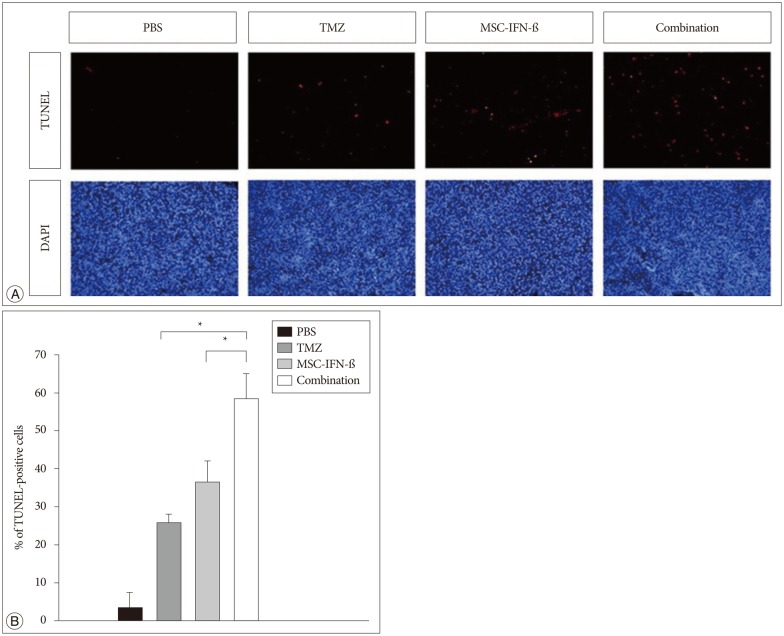
To figure out the mechanism of the combined treatment, we focused on apoptosis. To determine whether apoptotic cell death was involved in the antitumor activity of transplanted MSC-IFN-β and TMZ, TUNEL staining and counterstaining with DAPI were conducted. TUNEL staining demonstrated a significant increase in the number of apoptotic cells in the combined treatment group compared to the number in each of the monotherapy groups (p<0.05, TMZ vs. combination and MSC-IFN-β vs. combination) (Fig. 4). These results demonstrate that the antitumor activity of transplanted MSC-IFN-β and TMZ is mediated by apoptosis.
DISCUSSION
By engineering MSCs to secret IFN-β and using them in combination with TMZ, the current study demonstrates that when applied in combination, MSC-IFN-β and TMZ have significant antitumor effects in vitro, and are able to significantly reduce the tumor size and enhance the survival rates in GL26 mice. We demonstrated that the combination of MSC-IFN-β and TMZ showed significant antitumor effects in vitro, and significantly reduced the tumor size and enhanced the survival rates in vivo when compared with each of the monotherapy groups (MSC-IFN-β or TMZ alone). In addition, though it was not significant, the tumor size of glioma-bearing mice treated with MSC-IFN-β alone was reduced more than the tumor size in the TMZ monotherapy group in the in vivo experiment. This result shows the therapeutic advantage of pathotropism, which was acquired by using the MSCs. Through the intratumoral accumulation of IFN-β and the pathotropism of MSCs, combination therapy can deliver a higher concentration to the tumor and exert synergic effects.
Moreover, in the previous study that combined IFN-β and TMZ, continuous intravascular infusion of the drug was needed11). However, in this study, by intratumorally injecting MSC-IFN-β, continuous intravascular infusion was not needed to maintain the therapeutic concentration. Although our study showed advanced antitumor effects using the combination of MSC-IFN-β and TMZ for glioma treatment, we should compare these results with the outcomes of combining TMZ and IFN-β without MSCs in the future.
To evaluate the mechanism of the combination effect, we conducted TUNEL staining to demonstrate apoptotic cell death. The results showed a significant increase in the number of apoptotic cells in the group treated with the combination therapy. Further studies are needed to determine the exact mechanisms of apoptosis in this combination therapy. Other studies have suggested additional potential mechanisms, with one being the inhibition of angiogenesis2). More specifically, MSCs have been shown to suppress human glioma growth through the inhibition of angiogenesis2). Additional studies (such as signal network research) are needed to establish the mechanisms of the antitumor effect as apoptosis, as was observed in this study.
CONCLUSION
In summary, we focused on the pathotropism of MSCs and attempted use these cells as a delivery vehicle. In addition, we used IFN-β to down-regulate the expression of MGMT to enhance the antitumor effects of TMZ. Using both MSC-IFN-β and TMZ in combination, we found significantly enhanced antitumor effects in vitro and in vivo. These results suggest that the combination of MSC-IFN-β and TMZ could be considered as a new option for the treatment of malignant gliomas.
 XML Download
XML Download