

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
A neural progenitor cell (NPC) has the capability to divide, migrate and differentiate into neurons, and NPC proliferation is an important pathway in functional recovery after spinal cord injury (SCI)1,2). Cellular and molecular mechanisms have been studied to understand NPC proliferation31). Recently, mitogen-activated protein (MAP) kinase signaling pathways are among the most important mechanisms of NPC proliferation, and extracellular signal-regulated kinases (ERKs), p38 kinases and c-Jun NH2-terminal kinases (JNKs) are well known MAP kinases5,8,15,29).
Curcumin (diferuloylmethane) is found in the rhizomes of the plant Curcuma longa Linn (tumeric). Traditionally, tumeric has been used as a dietary spice and as a treatment of diseases associated with injury and inflammation9,10). Curcumin has a diverse set of biologic effects, including anti-inflammatory and antioxidanteffects3,11,12,22). Recently, some studies suggest the possibility that curcumin could increase the proliferation of subventricular zone (SVZ) originated NPCs via a MAP kinase signaling pathway1,23,32).
However, SVZ-NPCs and spinal cord NPCs (SC-NPCs) have some differences involving gene expression and proliferation characteristics. Liu et al.19) reported different electrophysiological properties of mitogen-expansion between adult rat SC-NPCs and SVZ-NPCs. Also, Pfenninger et al.27) reported different gene expressions and proliferations between adult spinal cord ependymal cells and SVZ originated neural precursor cells.
Therefore, the objectives of this study are to evaluate the effect of curcumin on SC-NPC proliferation and to clarify the mechanisms of the MAP kinase signaling pathways in SC-NPCs.
MATERIALS AND METHODS
NPC cultures
To establish cultures of SC-NPCs, Sprague-Dawley rats weighting 250 g to 350 g were sacrificed and their spinal cords were extracted. The spinal cords were chopped using microscissors and incubated in a cocktail containing papain (0.1%, Worthington Biochemical Corp., Lakewood Township, NJ, USA), dispase (0.1%), DNase (0.01%), and MgSO4 (12.4%) in a Hanks balanced salt solution with glucose (0.45%) for 30 minutes at 37℃. The dissociated cells were cultured to form neurospheres in a neurobasal medium containing B-27, glutamin (2 mmol/L), penicillin/streptomycin (0.1 g/mL), fibroblast growth factor-2 (20 µg/mL), and heparin (2 µg/mL). Every 5 days, the neurospheres were dissociated with accutase and NPCs were plated into 96-well plates or culture dishes containing culture medium. The NPCs between 5 and 25 passages were used for the experiments.
NPC proliferation assay
The NPCs were suspended in 100 uL of a neurobasal medium containing curcumin and cultured in a 96-well plate. In different experiments, various concentrations of curcumin (0.1-50 µmol/L) were used. A 20-uL cell proliferation assay solution containing the tetrazolium compound MTS (3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfo-phenyl)-2H-tetrazolium), (CellTiter 96 Aqueous One Solution, Promega®, Fitchburg, WI, USA) was added to a well containing cultured cells and incubated for 2.5 hours (hrs) at 37℃. The optical density was measured at 490 nm with a microplate spectrophotometer. The same volume of culture medium with the assay solution was used as a blank. The optical density at 490 nm after reaction with MTS showed a proportional correlation to the number of cells. To study the effect of the duration of curcumin, cells were seeded at a density of 1×104 cells in 96-well plates. After 48 hrs, the cells were treated with different concentrations of curcumin (0.1, 0.5, 1, 10, 20, and 50 µM).
MAP kinase signaling pathway analysis
We used an immunoblotting method for evaluating the MAP kinase signaling protein. The protein concentration was mixed with a RIPA buffer (25 mM Tris-Cl pH 7.6, 150 mM NaCl, 1% NP-40, 1% sodium deoxycholate, 0.1% SDS) containing a protease inhibitor cocktail (Roche®, Mannheim, Germany) and immediately homogenized. The homogenate was centrifuged at 13000 rpm for 30 min at 4℃ and the supernatants were determined using the BCA protein assay (Sigma®, St. Louis, MO, USA). Proteins were separated by a 10% SDS-PAGE gel and transferred to a nitrocellulose membrane. Incubation was in a blocking solution of 5% non-fat dry milk in Tris-buffered saline containing 0.1% Tween 20 for 1 hr at room temperature. The membrane was incubated with a 1 : 1000 dilution of mouse monoclonal phospho-ERK, phospho-p38, phospho-JNK (Santa Cruz®, CA, USA), β-actin (Sigma®, St. Louis, MO, USA) and mouse monoclonal anti-β-actin (Sigma Aldrich®, Saint Louis, MO, USA) overnight at 4℃, and then with a horseradish peroxidase-conjugated secondary antibody for 1 hr at room temperature. The proteins were detected with chemiluminescence reagents. Immune-positive bands used an image J, version 1.46 r, computer-assisted image analyzer (National Institutes of Health, Bethesda, MD, USA).
Statistical analysis
All statistical comparisons were computed using SPSS 17.0 (SPSS Inc., Chicago, IL, USA). Data are expressed as mean±standard error of the means. Repeated measure analysis of variance was used to compare groups. Null hypotheses of no difference were rejected if p-values were less than 0.05.
RESULTS
Curcumin increases the proliferation of SC-NPCs
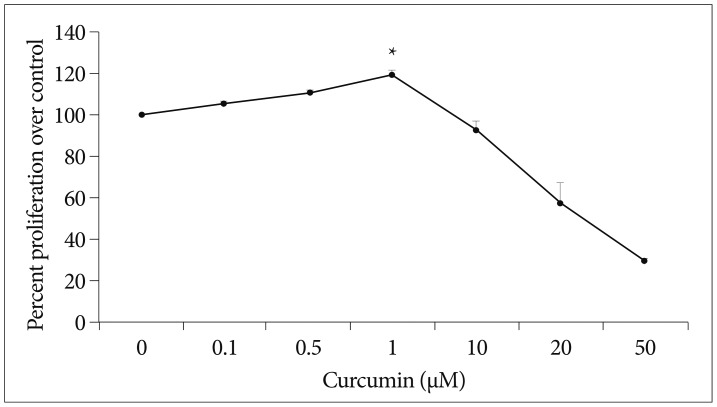
To study that the effect of curcumin on the proliferation of SC-NPCs, SC-NPCs growing in 96-well plates were maintained in medium lacking or containing increased concentrations of curcumin, and the cell proliferation rate was quantified at different time points. After 48 hours of curcumin treatment, lower dosage (0.1, 0.5, 1 µM) of curcumin increased NPC proliferation, whereas high dosage (≥10 µM) caused a decrease in NPC proliferation (Fig. 1). The result shows a few percent improvement compared to each control group.
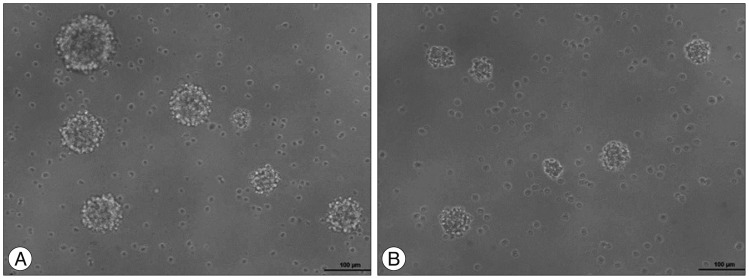
To evaluate the neurosphere formation, dissociated SC-NPCs were diluted in a culture medium and plated into 96-well plate dishes and cultured for 48 hrs. The cells were treated with 1 µM curcumin for 24 hrs. NPC proliferation and neurosphere formation of the curcumin treated group (Fig. 2A) were increased compared to the control group (Fig. 2B).
The proliferation of NPC via the MAP kinase pathway
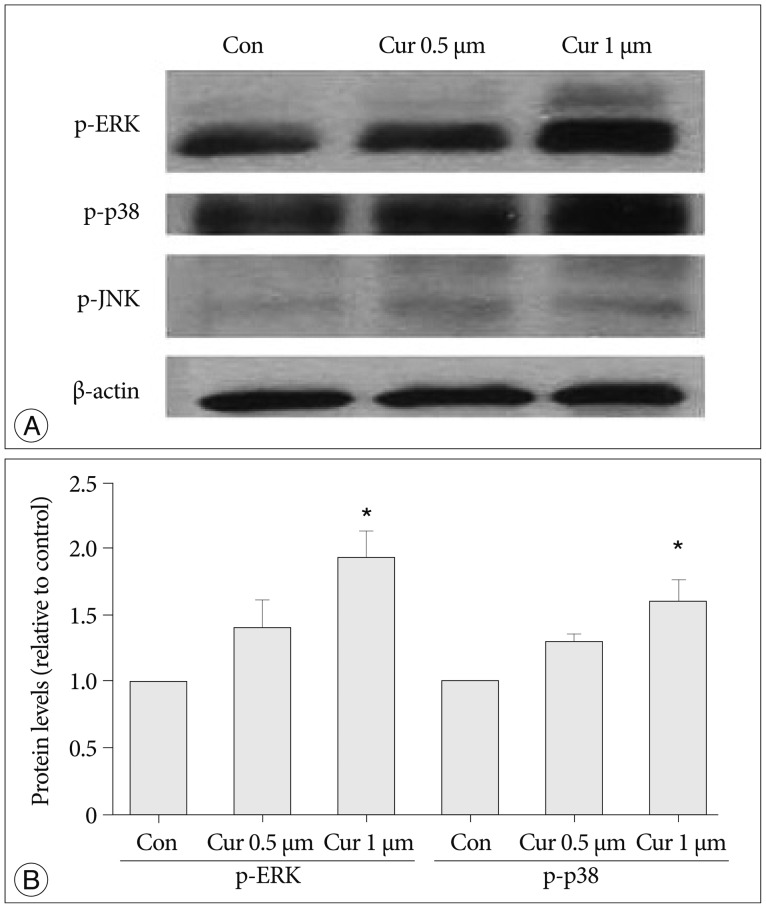
We investigated curcumin treatment associated with MAP kinase signaling pathways, especially involving ERK, p38, and JNK proteins related to NPC proliferation19,29). Immunoblot analysis was used on antibodies against phospho-ERK (p-ERK), phospo-P38 (p-p38), and phosph-o-JNK (p-JNK), and levels of β-actin were determined as a control against possible protein loading variability. The p-ERK protein and p38 protein levels show increases depending on curcumin dosage (0.5, 1 µM, p<0.05), whereas p-JNK and β-actin protein levels show marginal differences without depending on curcumin dosage (Fig. 3).
DISCUSSION
SCI has been thought to be irreversible and the cause of critical sequela. Many studies have reported a desire to minimize secondary injury and increase functional recovery from neurological deficit16,17,21,30). Recently, NPC proliferation after SCI plays an important role in functional recovery from neurological dys-function18,26).
This study finds that curcumin stimulates NPCs proliferation through a MAP kinase signaling pathway, and especially involves the ERK protein and the p38 protein. Also, the effect is biphasic in that low concentrations stimulated cell proliferation, while high concentrations inhibited it15,24,28,29).
Previous studies have documented that curcumin could affect proliferation of stem cells through diverse pathways. Kim et al.13) found that a low concentration of curcumin stimulates proliferation and stemness acting signals of pre-adipocytes by the phosphoinositide 3-kinase pathway, whereas it inactivates p38 kinases and JNK protein. Mujoo et al.25) reported that curcumin induces differentiation of embryonic stem cells through nitric oxide-cyclic signaling. Kim et al.15) documented that curcumin stimulates proliferation of embryonic NPCs and neurogenesis in the adult hippocampus. The study suggested that a MAP kinase signaling pathway is connected to NPC proliferation through ERKs and the p38 protein.
In the present study, we investigated the effect of curcumin on SC-NPCs and analyzed MAP kinase signaling. According to the results, SC-NPC proliferation increased with lower dosage (0.1, 0.5, 1 µM) of curcumin. But, higher dosage (10, 20, 50 µM) of curcumin decreased the NPC proliferation rate. These results are comparable with previous studies showing that curcumin stimulates NPC proliferation relative to the dosage15,28,29). Consequently, for effective clinical applications, these results suggest that proper dosage is important. Therefore, in future studies, we expect that setting the appropriate curcumin-dosage with human cells will be very important.
We found that curcumin affects the proliferation of SC-NPCs, and likewise it affects SVZ-NPCs via the MAP kinase signaling pathway, as we analyzed MAP kinase proteins in this study. Especially, the MAP kinase signaling pathway was revealed as an important mechanism for most stem cell proliferation. There are many different types of MAP kinase and stem cells originated each organization was founded to have slightly different mechanism of MAP kinases5,8,15). Our findings are the first study to show that SC-NPCs are more relevant to ERK and p38 protein associated MAP kinases. The ERK protein has been reported to be concerned with cell proliferation, survival and apoptosis7), and the p38 protein played a vital role in cellular responses to external stress signals like anti-inflammatory reactions6). Anti-inflammatory effects might promote the proliferation of NPCs. Our finding suggests these effects are relevant to curcumin effect NPC proliferation. We consider that this result would be helpful to more selective agonists for NPC proliferation.
However, this study has some limitations. First, this study involves only in-vitro experiments. The spinal cord is a complicated environment, so we need further in-vivo study to evaluate the curcumin effect in live tissue. Second, we used rat SC-NPCs in this experiment. The rat SC-NPC is different from the human SC-NPC, so the proper dosage could be different in humans.
CONCLUSION
This study suggests that curcumin stimulates NPCs proliferation through a MAP kinase signaling pathway, and especially involves the ERK protein and the p38 protein. Also, the effect is biphasic in that low concentrations stimulated cell proliferation, while high concentrations inhibited it. In future studies, we should evaluate the effect of curcumin and MAP kinase signaling in a SCI model.
 XML Download
XML Download