

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
All movements we perform are controlled by the brain. The motor system controls complex neuromuscular networks. The motor system includes cortical and subcortical gray matter (GM); descending tracts including corticobulbar, corticospinal, rubrospinal, reticulospinal, vestibulospinal, and tectospinal tracts; the spinal cord GM; spinal nerves; and the cerebellum and basal ganglia. In addition, modulation based on sensory inputs is important for fine movement.1
The motor system allows people to perform various kinds of sports. The structural brain plasticity of elite athletes has been reported in several studies. Hänggi et al.2 reported structural brain changes in handball players and reviewed structural changes in ballet dancers, golf players, basketball players, speed skaters, high diving athletes, badminton players, mountaineers, Judo wrestlers, slackline dancers, gymnasts, climbers, martial artists, and endurance athletes. Structural changes were different according to specific sports, although sample size and analysis method also differed by sport.
Our laboratory has focused on basketball players since 2000. There was no overall volume difference in the cerebellum between basketball players and the control group.3 Nonetheless, a detailed analysis of the cerebellum in subdivisions4 showed a difference in vermian lobules VI–VII (declive, folium, and tuber) of elite basketball athletes, which resulted from an increase in white matter (WM) volume.5 In addition, the striatum volume was larger in the athletes group.6
The aforementioned studies from our laboratory showed structural changes in the cerebellum and striatum based on manual segmentation.7 Nevertheless, changes in the cerebral cortex of basketball players were not assessed. In this study, we analyzed structural changes in the cerebral cortex of basketball players using fully automated magnetic resonance imaging (MRI) brain analysis techniques.
Go to : 
METHODS
Subjects
The raw MRI data in this study were the same as those used in a study previously published.3 Briefly, MRI was performed on 19 male college basketball players (athlete group, AG) and 20 age-, sex- and height-matched controls who did not exercise regularly (control group, CG). All subjects had healthy general medical and neurological profiles. Individuals in the AG had played basketball for 8 years on average and had practiced for 5 hours daily on average.
MRI acquisition
MRI images were scanned on a 1.5-T Magnetom vision instrument (Siemens, Erlangen, Germany) using T1-weighted magnetization using a rapid acquisition gradient echo sequence (repetition time = 9.7 msec, echo time = 4 msec, flip angle = 12 degrees, slice thickness 2.0 mm without gap, field of view 250 mm, number of slices 90, and matrix 256 × 256).
CAT12 processing
The volumetric 3D T1 MRIs of all participants were processed using the Computational Anatomy Toolbox (CAT12, http://www.neuro.uni-jena.de/cat) in SPM12 (http://www.fil.ion.ucl.ac.uk/spm) for voxel-based morphometry (VBM) and surface-based morphometry (SBM, estimation of cortical thickness [CT], and fractal dimension [FD]).
For VBM, MRIs were bias-corrected and spatially normalized to the DARTEL template by DARTEL high-dimensional spatial normalization.8 The accuracy of spatial normalization was visually checked using the Check Registration tool with the Montreal Neurological Institute (MNI) template image. Normalized MRI images were segmented into GM, WM, and cerebrospinal fluid (CSF) using an adaptive maximum a posteriori technique. The voxel values of the GM partitions were multiplied by the nonlinear components, which were derived only from the normalization matrix to preserve the actual GM values locally (modulation process). Modulated GM was smoothed using an 8-mm full width at half maximum (FWHM) isotropic gaussian kernel (IGK). The values of individual intracranial cavity volume (ICV) were automatically calculated from the GM, WM, and CSF partitions.
Analysis of covariance (ANCOVA), controlled for age and ICV, was applied for analyses of regional volume changes of GM and WM.9 To exclude edge effects between the boundaries of different tissue types, an absolute threshold masking value of 0.15 was applied. The statistical threshold level was set to uncorrected P < 0.001, and the extent threshold was set to k
E > 100 voxels (338 mm3; 1 voxel = 3.375 mm3).
Coordinates were defined by the MNI coordinate system. The cluster regions were localized using information about their local maxima, based on the standardized template and defined by visual comparison with the atlas of Henri M. Duvernoy.10
For SBM, the CAT12 toolbox included the calculation of CT and the central surface of the bilateral hemisphere based on projection-based thickness.11 The pipeline of surface analysis included topological correction, spherical mapping, spherical registration, and FD.12 After GM and WM segmentations, the WM distance was calculated, and the local maxima were projected to other GM voxels calculated by the WM distance. The estimated GM and WM boundaries were constructed by classifying all WM voxels in the MRI. After computing the curvature of the surface, the surface representation was reconstructed to obtain the finest scale of the local surface curvature. The CTs of individuals in the AG and CG were calculated using the closest distance between the vertices of the two reconstructed surfaces. Additional surface parameters, such as the local gyrification index and FD, were extracted using the surface tool of CAT12.
The group differences of CT and FD were tested separately using ANCOVA, using the confounder of age with 5,000 permutations. The significance level was set to an uncorrected P < 0.001.
Go to :
RESULTS
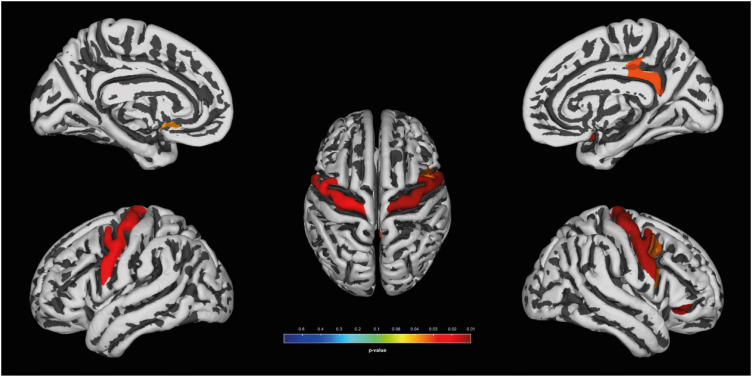
In the VBM of GM, increased GMV (Fig. 1) were found in the both precentral gyri and paracentral lobules (left, x, y, z= −2, −12, 72; kE = 344, Punc = 0.001; right, x, y, z = 10, −26,78; kE = 165, Punc = 0.001) and right antero-superior temporal gyrus (x, y, z = 51, 16, −16; kE = 1,404, Punc = 0.0012) of basketball players. In the VBM of WM (Fig. 2), the both precentral gyri (left, x, y, z = −14, −28, 62; kE = 406, Punc = 0.0008; right, x, y, z = 33, −30, 57; kE = 241, Punc = 0.001) of basketball players were shown the increased WM volumes.
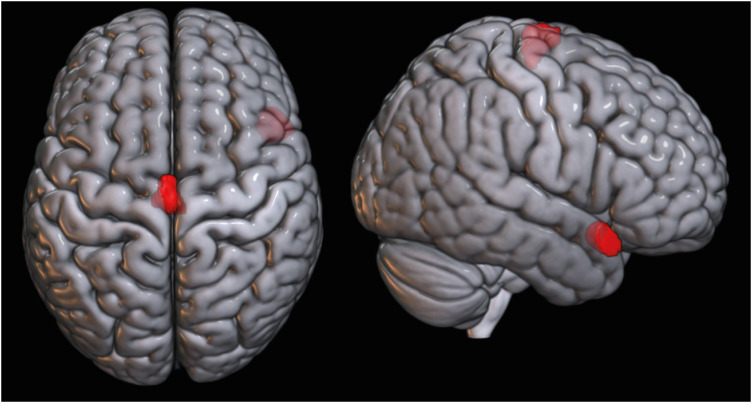 | Fig. 1Increased GM volumes in basketball players. Increased GM volumes are observed (uncorrected P < 0.001) in both precentral gyri, paracentral lobules and right antero-superior temporal gyrus of basketball players.GM = gray matter.
|
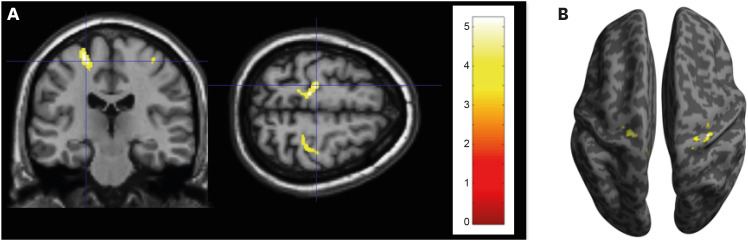 | Fig. 2Increased WM volumes in basketball players. Increased WM volumes are observed (uncorrected P < 0.001) in both precentral gyri of basketball players. (A) The left dominantly has increased WM volumes (radiological view). (B) The increased WM volumes are overlaid on the inflated brain surface. The color bar represents t values.WM = white matter.
|
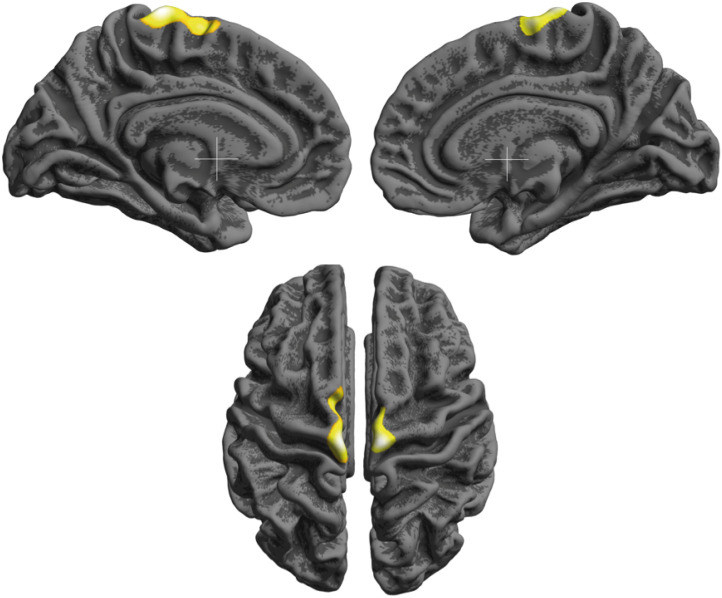
In the SBM, the CT of basketball players was left dominantly increased in the both pericentral area including paracentral lobules (Fig. 3, left, x, y, z = −6, −26, 74; kE = 991, Punc = 0.0007; right, x, y, z = 8, −24, 75; kE = 755, Punc = 0.0009). Basketball players had increased FD in both precentral gyri, the left subcallosal gyrus, and the right posterior cingulate gyrus (Fig. 4, FDR < 0.05), according to the region of interest analysis, defined by Destrieux atlas.13
Go to :
DISCUSSION
This study showed an increase of gray and white mater volumes in both primary motor area including superior part of precentral gyri and anterior paracentral lobules of the basketball players. In addition, we observed increased fractal complexity in the bilateral precentral gyri and, and increased CT of both pericentral areas and paracentral lobules. These findings imply that primary motor cortices are well developed in the basketball players. The motor homunculus map shows that anterior paracentral lobule controls lower extremity (thigh, leg, and foot), and small superior portion of precentral gyrus controls hip.14 Therefore, Figs. 1 and 3 suggest that the important role of the lower extremity motor control in elite basketball players. Optimal eye-hand coordination requires rapid processing of sensory information coming from external stimuli, and control of continuous movement in the arms, legs, and torso.4 These skills can be improved with practice, subsequently activating the neural circuit and related brain regions, including the motor cortices, the primary motor, premotor, and supplementary motor cortex, thereby enhancing the plasticity of a basketball player’s precentral gyrus. The pericentral area includes a portion of the precentral gyrus and of the postcentral gyrus. Somatosensory input is essential for precise motor performance based on learning and memory.15 In this context, increased CT of the primary sensory area is expected. Changes of the precentral gyrus in response to sporting activities were also reported in the brains of Judo wrestlers, gymnasts, and handball players.21617
The increased gray mater volume of the right antero-superior temporal gyrus is consistent with the report by Karnath et al.18 In their report, new anatomical functional brain localization based on MRI analysis was observed on patients with lesions. Karnath et al.18 reported that the right superior temporal cortex is closely related to spatial awareness based on patients with lesions leading to special neglect. These patients set up new functional localization of spatial awareness, which was previously considered a function of the parietal lobe.18 The specific increase in the right superior temporal lobe of basketball players could be understood by simulating a basketball game. Recognition of spatial information is key during a real basketball game.
The increased FD of both precentral gyri in basketball players is expected considering these areas are primary motor cortices and the plasticity observed in this study. Changes in the left subcallosal gyrus and right posterior cingulate gyrus were observed with low significance levels in this study. The posterior cingulate gyrus is activated during complex motor skill learning,19 and relate to voluntary and involuntary memory recall.20 The posterior cingulate cortex governs spatial attention-shifting tasks, with oculomotor approach behaviors based on internal expectancies. The neural network underlying this phenomenon is supposedly the retrosplenial-cingulate-thalamic-dorsolateral frontal cortical connectivity. Actually, patients with retrosplenial injury or an anterior thalamic stroke had driving disabilities due to impaired spatial aiming in far space.21 Auger and Magurie22 reported that good navigators decoded the number of permanent items in view significantly better than poor navigators, based on patterns of activity in the retrosplenial cortex. These data suggest a link between brain plasticity in basketball players and increased fractal complexity in the posterior cingulate gyrus. The functions of the subcallosal gyrus, a portion of the limbic system behind the cingulate cortex are reciprocal to those of the cingulate cortex. The subcallosal gyrus inhibits motor neuron activity, whereas the cingulate cortex enhances motor neuron functions (APA Dictionary of Psychology). This structure likely regulates movement, but more research is needed to accurately link the association with basketball play. FD was originally a mathematical index for characterizing fractal patterns of geometric objects. Currently, it is used to estimate cortical complexities in many neuropsychiatric and neurological disorders.23 Cortical complexity calculated by FD showed a significant change with age in normal children.24 FD is another way to measure the complexity of cortical folding and complements the gyrification index. FD renders the brain as a fractal structure and mathematically quantifies the degree of complexity.12 An increased FD value in the cortical area could suggest a more convoluted cortex structure.
The brain regulates every activity we perform, and it shows some plastic change from the synaptic to the behavioral level when learning is induced by continuous repetitive activity. Black et al.,25 showed increased synapse numbers per Purkinje cell as a result of acrobat motor skill learning, whereas simple exercise such as treadmill training induced increased vascular density.24 A few studies revealed specific strengthening of synaptic networks.262728 As previously mentioned, studies that analyzed the brains of elite athletes using MRI showed changes in tract, volume, or thickness of the brain region related to specific sports.2 These kind of brain plasticity is observed in the patient recovering stroke with constraint-induced movement therapy.29 The underlying cellular mechanisms are likely synaptogenesis, increased neuronal or glial cell volume, and axonal remodeling.730
Some limitations of this study are number of subjects and cross sectional study, the reason why we cannot describe cause and result of basketball training. We checked the training career as an athlete, but the objective data presenting motor skill related to basketball (parameters) were not checked, which should be considered in next study design like this researches.
As we hypothesized, the strong cerebral plasticity of both precentral gyri were shown by white and gray mater volume, as well as fractal complexity. The increase of CT in pericentral area and paracentral lobule was also observed bilaterally. These results suggest that motor system circuits favorably respond in basketball players. The moderate increase in fractal complexity of the right posterior cingulate gyrus implies that spatial aiming and navigation are important abilities for basketball players. Our data support the recently observed spatial awareness function of the superior temporal gyrus, which would be important in basketball payers. The plastic regions shown in this study, linked to functional neuroanatomy, were related to the competence required to play basketball. In addition, novel brain functions related to sports competency could be explored in the future.
Go to :
 XML Download
XML Download