

 PDF
PDF Citation
Citation Print
Print



서론
PI3K-AKT 신호전달은 세포 외 신호에 반응하여 단백질 합성, 세포증식 및 생존을 통해 세포성장을 조절하는 신호전달이다.1 성장인자, Chemokine, 그 외 다양한 신호전달은 세포 표면 수용체를 활성화시키고 PI3K의 인산화를 유발한다. 활성화된 PI3K는 원형질막의 지질을 인산화 하고 phosphatidylinositol (3,4,5)-trisphosphate (PIP3)를 생성한다. Serine/threonine-protein kinase (AKT)는 이러한 phosphoinositide docking site와 반응하여 막 주변으로 모여 활성화된다.2 활성화된 AKT는 다양한 세포 내 단백질을 인산화하며 세포생존, 세포성장, 세포증식, 세포이주 및 혈관생성에 영향을 미친다.3 Serine/threonine-protein kinase (AKT)는 세포생존, 전이, 약물 내성, 대사, 방사선 저항성을 포함하는 주요 세포 전달 체계의 주단백질이며,4 AKT1, AKT2, AKT3 등 세 가지의 AKT 동형단백질로 존재한다. 세 가지의 단백질은 높은 상동 관계를 가지지만 세 가지의 다른 유전자로부터 조직의존적으로 발현되므로 다른 기능을 가질 것이라고 추측되고 있다. 유전자제거 생쥐 연구에 따르면 AKT1은 세포생존에 필수적이고, AKT2는 포도당 항상성에 주된 역할을 한다고 보고되었으며, AKT3는 두뇌 발달에 영향을 미친다고 추측되고 있다.5 AKT가 억제되면 세포생존을 위한 중요한 신호전달의 억제를 초래할 수 있고, receptor tyrosine kinase (e.g. HER3, IGF-1R, insulin receptor)와 같은 다른 후속 단백질이 자극되어 이에 따라 암세포가 생존할 수 있게 된다.6
Phosphoinositide 3-kinase (PI3K)는 다양한 신호전달 에 의해 활성화되는 여러 가지 하위유형으로 나타난다. PI3K는 receptor tyrosine kinases에 의한 인산화,7 src의 non-receptor protein tyrosine kinases(PTK),8 focal adhesion kinase9 등에 의하여 활성화되거나 βγ subunit of activated heterotrimeric G proteins에 의해 활성화된다. PI3K의 길항제는 인산효소 Phosphatase and tensin homolog (PTEN)10,11와 Src homology 2 domain containing inositol polyphosphate 5-phosphatase (SHIP)11,12 가 있으며 Phosphatidylinositol (3,4,5)-trisphosphate (PIP3)에서 인산을 제거한다.
선행연구들에서, trichloroacetic acid (TCA)의 영향이 세포수준에서 평가되었고,13 세포 동물 모델에서 hydrophobically modified glycol chitosan (HGC)기반 나노방출제어시스템을 사용하여 TCA와 Epidermal growth factor (EGF)의 영향도 평가되었다.14 TCA가 인간치은섬유아세포에서 다양한 유전자 발현의 변화를 유도한다는 사실이 확인되었고, TCA와 같은 강산을 구강환경에 적용 시 발생될 수 있는 위험을 해결하는 방법으로 HGC 기반 나노방출제어시스템이 제안되었다. HGC기반 나노방출제어시스템의 안정성이 확인되었으며 TCA를 빠르게 방출하는 동시에 EGF를 천천히 방출하는 방법이 소개되었다. 인간치은섬유아세포와 성견의 견치구개점막의 조직세포의 유전자 발현의 변화를 또한 확인되었다.13,14 이전연구의 DNA microarray 분석과 successive Kyoto Encyclopedia of Genes and Genomes (KEGG) 신호전달 분석결과로부터 TCA와 EGF를 순차적으로 방출하고 적용할 때 유전자 발현이 변할 수 있는 29개의 세포생존에 관한 유전자가 선택되었다.
본 연구의 목적은 구강연조직재생에 있어 인간치은섬유아세포에 HGC기반 TCA-와 EGF- 담지형 나노방출제어시스템을 적용 시 TCA와 EGF의 순차적 방출이 세포생존 유전자들의 발현에 미치는 영향을 규명하는 것이다. 본 연구에서는 선행연구에서 언급된 PI3K-AKT 신호전달과 연관된 29개의 유전자들에 대한 실시간 중합효소연쇄반응 분석을 실시하였고 AKT에 대한 다른 28개의 유전자들의 영향을 확인하였다.
Go to : 
연구 재료 및 방법
TCA-와 EGF- 담지형 나노방출제어시스템을 제작하기 위해 글리콜 키토산(1.0 g, 0.4 × 10-5 mol), 5β-콜란산(123.0 mg, 0.34 × 10-3 mol), N-hydroxy succinimide (NHS) (58.6 mg, 0.51 × 10-3 mol), N-(3-dimethylaminopropyl)-N’-ethylcarbodiimide hydrocholoride (EDC•HCl) (97.6 mg, 0.51 × 10-3 mol), TCA (Sigma-Aldrich Co., St. Louis, USA), 메탄올(Sigma-Aldrich Co., St. Louis, USA), EGF (Peprotech, London, UK)를 구매하였고 시약으로 사용하였다.14
5β-콜란산을 활용한 글리콜 키토산 나노입자들은 글리콜키토산(1.0 g, 0.4 × 10-5 mol)과 5β-콜란산 (123.0 mg, 0.34 × 10-3 mol)을 10 ml의 증류수/메탄올 용액(1;1, v/v)에 용해하여 제작하고 뒤따라 N-(3-dimethylaminopropyl)-N’-ethylcarbodiimide hydro- choloride (EDC•HCl) (97.6 mg, 0.51 × 10-3 mol)과 N-hydroxysuccinimide (58.6 mg, 0.51 × 10-3 mol)을 24시간동안 교차반응시키며 첨가하였다. 투석막(molecular weight cut-off: 3,500 g/mol)을 이용하여 24시간동안 투석진행 후 정제하였다. 증류수/메탄올 용액(1:3; 6 h, 1:1; 6 h, 3:1 6 h, v/v)을 투석 용매로 사용하였다. 정제과정에서 나온 산물은 감압하 동결건조 되었고 최종 HGC 나노입자(78.4%의 입자수득율)를 얻었다.14
TCA-담지형 나노입자들은 10 ml의 증류수에 HGC를 분산시켜 제조하였고, TCA (1.0 mg, 0.6 × 10-5 mol)를 0.025 ml의 증류수에 용해하여 용액을 제작하였다. 그 후, TCA 용액을 HGC 용액에 첨가하였고, 600 rpm으로 교차반응시켰다. 그 후에, TCA 입자를 정제하기 위해 24시간 투석이 투석막(MWCO: 3,500 g/mol)을 통해 진행되었고 감압하동결처리하여 최종 TCA-담지형 나노입자들을 얻었다. 첨가된 TCA의 양은 이온 크로마토그래피를 이용한 Cl 정량분석을 통해 측정하였다. 0.5%의 TCA 담지형 나노입자 용액을 제조하기 위해 4 ml의 증류수를 TCA (0.00714 mg TCA/1 mg TCA 첨가 나노입자)가 삽입된 2.8 mg의 나노입자에 첨가하였다.14
EGF-담지형 나노입자를 제조하기 위하여, 10 mg의 HGC를 10 ml의 증류수에 분산시킨 후 EGF (0.1 mg, 0.15 × 10-7mol)를 0.1 ml의 증류수에 용해시켜 용액을 제작하였다. EGF용액을 HGC용액에 첨가 후 300 rpm으로 교차반응시켰다. 그 후, 반응하지 않은 EGF를 입자들로부터 제거하기 위해 투석막(MWCO; 10,000 g/mol)을 통해 24시간 투석하였고 감압하 동결건조하여 최종 EGF-담지형 나노입자들을 얻었다. 담지형-EGF의 양은 ELISA (Human EGF Standard ABTS ELISA Development Kit, Peprotech, London, UK)를 통해 측정하였다. TCA-담지형 나노입자(2.8 mg)에 증류수 2 ml 추가한 용액과 EGF-담지형 나노입자(8.98 μg EGF/ 1 mg EGF 담지형 나노입자)에 배지 2 ml를 추가한 용액을 혼 합하여 0.5% TCA 담지형 나노입자와 80 μg의 EGF 담 지형 나노입자의 복합 용액을 제조하였다.14
세포 배양을 위해 ATCC(ATCC® PCS-201-018™, Manassas, USA)에서 인간치은섬유아세포를 구입하였다. 10% 우태아혈청 (FBS, Sigma-Aldrich Co., St. Louis, USA)과 항생제를 포함하는 dulbecco’s modified eagle’s medium (DMEM, Invitrogen Corporation, Carlsbad, USA)에 세포를 부유시켰다. 부유된 인간치은섬유아세포들은 37°C, 95% 공기, 5% 이산화탄소 하에 humidified incubator에서 배양되었다. 2 - 3주기 동안 배양된 인간치은섬유아세포를 본 연구에서 사용하였다.13
유전자 발현 분석을 위해 세포생존과 연관된 PI3K-AKT signaling 신호전달에서 총 29개의 유전자를 선택하였다(Table 1).13,14 이 유전자들은 relative mRNA expression 분석을 위해 실시간 중합효소연쇄반응으로 분석하였고 다음과 같이 분류하였다: 대조군(CON), TCA-담지형 나노방출제어시스템 투여군(EXP1), TCA-와 EGF-담지형 나노방출제어시스템 투여군(EXP2). 인간치은섬유아세포는 24-well plate에 각 CON, EXP1, EXP2 그룹으로 나누어 37°C, 5% 이산화탄소 하에서 배양하였고 세포 농도는 1 × 104/well로 유지하였다. 배양 48시간 후, TRIZOL lysis solution (TRIZOL® Reagent, GIBCO BRL, Carlsbad, USA)을 통해 총 RNA를 추출하여, NanoDrop 1000 (NanoDrop Technologies, Wilmington, USA)를 사용하여 RNA의 concentration 을 진행하였다. 총 RNA로부터 1 μg 추출하여 iScript cDNA Synthesis Kit (Bio-Rad Laboratories, Berkeley, USA)를 이용하여 cDNA로 역전사시켰다. 유전자 발현 은 TaqMan® Gene Expression Assay Kit (Applied Biosystems, Waltham, USA)을 사용하여 확인하였다. 본 연구의 실시간 중합효소연쇄반응으로 사용된 유전자들은 Table 1과 같이 정리하였다. Chromo4 reverse transcription-polymerase chain reaction (Bio-Rad Laboratories, Hemel Hepstead, UK)은 iQ Supermix (Bio-Rad)를 사용하여 진행하였다. 유전자 발현은 MJ OPticon Monitor Analysis Software (Bio-Rad)를 이용하여 정량화하였다.13 측정된 값은 GAPDH의 발현 레벨에 의해 수정되었고 EXP1과 EXP2의 발현 레벨은 다수의 CON에서 표현되었다.
Table 1
Gene information used in real-time PCR (Taqman® gene expression assay)
![]()
유전자들의 발현량에 대한 실시간 중합효소연쇄반응을 각 유전자마다 독립적으로 5회 반복 진행하였고, 평균값과 표준편차를 계산하였다. 평균값을 비교하기 위하여 일요인분산분석을 시행하였고, post-hoc 비교를 위해 Tukey’s multiple comparison test B를 진행하였다. AKT 발현에 통계학적으로 유의미한 영향을 미친 유전자를 결정하기 위해 단계적 다중회귀분석을 시행하였다. 모든 통계 분석을 위해 SPSS 18.0 software (SPSS 18.0, SPSS Inc., Chicago, USA)를 사용하였다.
Go to :
결과
나노방출제어시스템에서 HGC 나노입자들은 캡슐화되어 있으며 화학물질을 방출하고 중성 pH용액에서 잘 용해된다.14 Chitosan의 hydrophilic cationic amine group (-NH2)는 hydrophobic cholanic acid와 결합한다. Hydrophobic cholanic acid는 물에서 핵을 형성하는 반면 hydrophilic cationic chitosan은 겉껍질을 형성하였다. 음이온의 TCA와 EGF는 이온결합을 통해 chitosan 껍질에 담지되었으며 in vitro와 in vivo 상황에서 순차적으로 방출될 것으로 생각되었다(Fig. 1).14
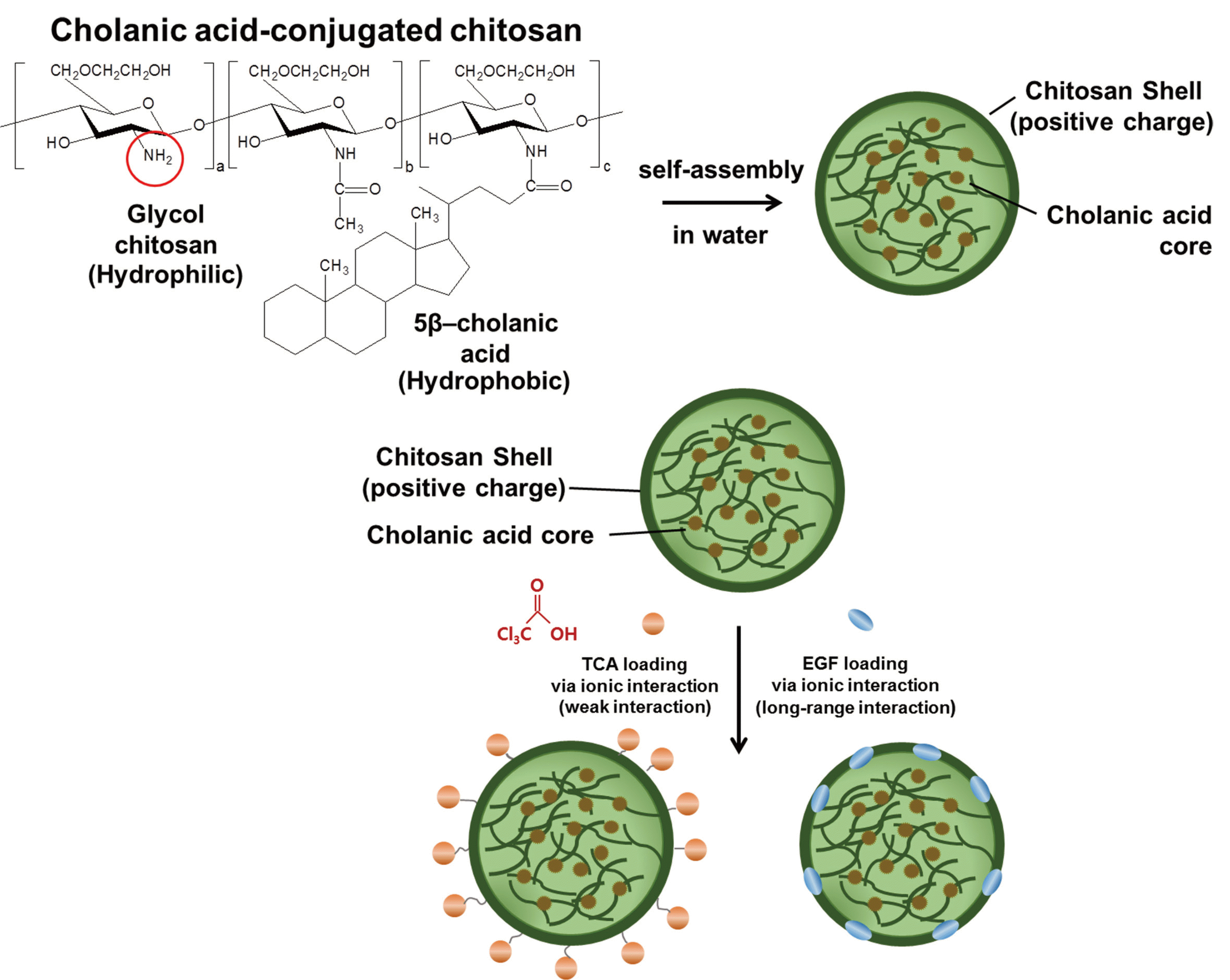 | Fig. 1Principle of HGC-based TCA- and EGF- loaded nano-controlled release system. The hydrophilic cationic amine group (-NH2) of chitosan was combined with hydrophobic cholanic acid. Hydrophobic cholanic acid formed nuclei in water, while hydrophilic cationic chitosan formed the outer shell. The anionic TCA and EGF were loaded on the chitosan shell through ion bonding.
|
실험 48시간 이후 실시간 중합효소연쇄반응 분석을 통하여 상향 조절된 유전자를 확인하였다. 성장인자와 연관된 신호전달의 유전자 실시간 중합효소연쇄반응 분석결과는 다음과 같다. Insulin like growth factor 1 receptor (IGF1R), epidermal growth factor receptor (EFGR), insulin receptor substrate 1 (IRS1)은 대조군에 비하여 EXP1과 EXP2에서 상향조절되었으며(P < 0.001), EXP2, EXP1, CON 순으로 유의미한 상향조절이 관찰되었다(P < 0.001, Fig. 2).
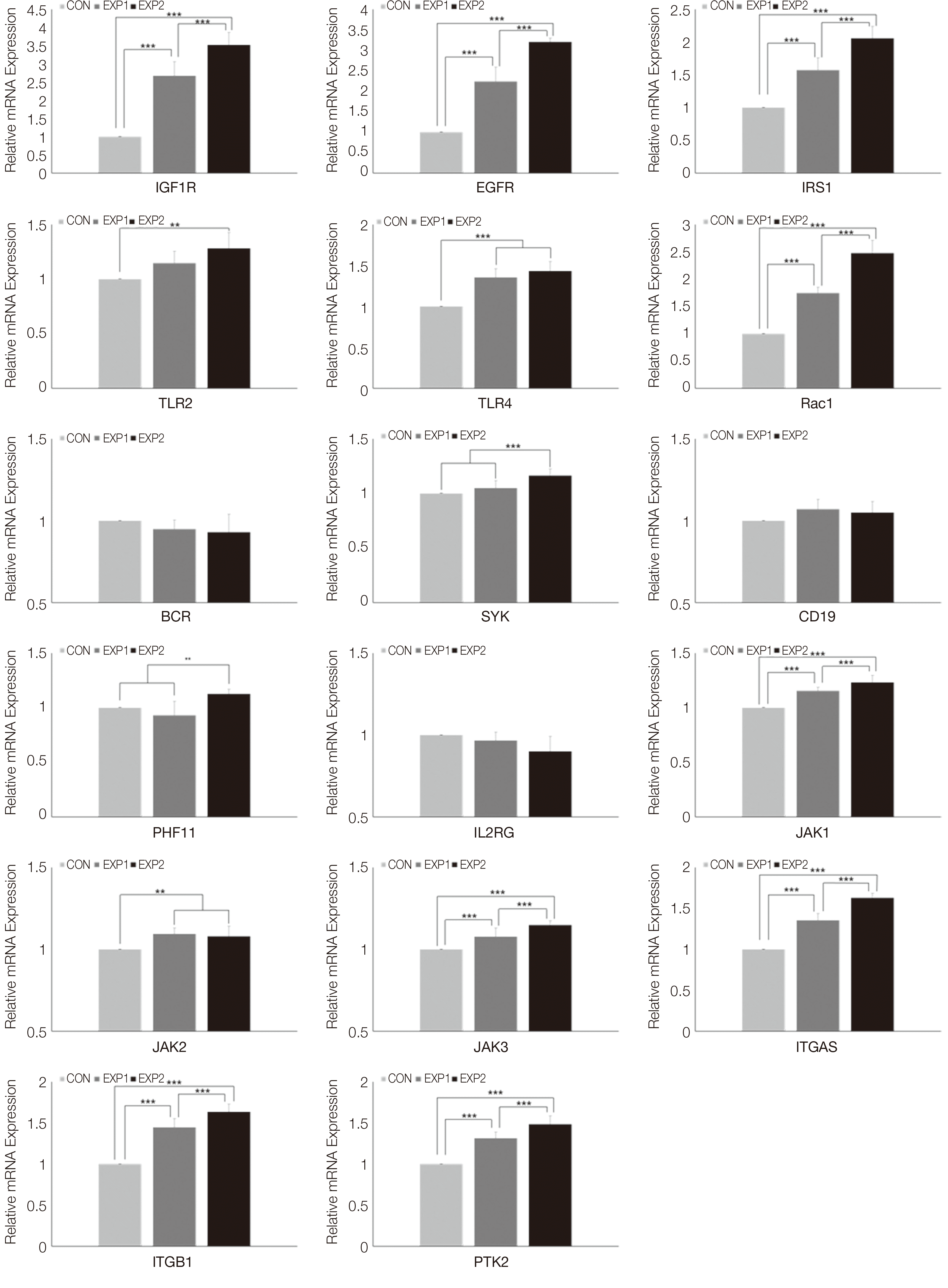 | Fig. 2Real-time PCR analysis results of Growth factor (IGF1R, EFGR, IRS1), Toll-like receptor signaling pathway (TLR2, TLR4, Rac1), B cell receptor (BCR) signaling pathway (BCR, SYK, CD19, PHF11), JAK/STAT signaling pathway (IL2RG, JAK1, JAK2, JAK3), Focal adhesion associated genes (ITGA5, ITGB1, PTK2). One-way ANOVA (***: P < 0.001, **: P < 0.01, *: P < 0.05).
|
Toll-like receptor signaling 신호전달의 유전자 실시간 중합효소연쇄반응 분석 결과는 다음과 같다. Toll like receptor 2 (TLR2)에서는 대조군에 비하여 EXP2에서 유의미한 상향조절이 관찰되었다(P < 0.01). Toll-like receptor 4 (TLR4)는 대조군에 비하여 EXP2와 EXP1에서 상향조절되었으며(P < 0.001), EXP2와 EXP1사이에 는 유의미한 차이가 없었다. Rac family small GTPase 1 (Rac1)은 대조군에 비하여 EXP1과 EXP2에서 상향조절 되었으며(P < 0.001), EXP2, EXP1, CON 순으로 유의미한 상향조절이 관찰되었다(P < 0.001, Fig. 2).
B cell receptor signaling 신호전달의 유전자 실시간 중합효소연쇄반응 분석 결과는 다음과 같다. B cell receptor activator for RhoGEF and GTPase (BCR)과 CD19 molecule (CD19)는 CON, EXP1, EXP2 사이에 어떠한 상관관계도 관찰되지 않았다. Spleen associated tyrosine kinase (SYK)와 PHD finger protein 11 (PHF11)은 대조군에 비하여 EXP2에서 유의미하게 상향조절되었다(각각 P < 0.001, P < 0.01)(Fig. 2).
JAK/STAT signaling 신호전달의 유전자 실시간 중합효소연쇄반응 분석 결과는 다음과 같다. Interleukin 2 receptor subunit gamma (IL2RG)는 CON, EXP, EXP2 사이에 어떠한 상관관계도 관찰되지 않았다. Janus kinase 1 (JAK1)과 Janus kinase 3 (JAK3)는 대조군에 비하여 EXP1과 EXP2에서 상향조절되었으며(P < 0.001), EXP2, EXP1, CON 순으로 유의미한 상향조절이 관찰되었다(P < 0.001). Janus kinase 2 (JAK2)는 대조군에 비하여 EXP2과 EXP1에서 상향조절되었고 EXP2와 EXP1 사이에 유의한 차이는 없었다(P < 0.01, Fig. 2). 국소 접착과 연관된 신호전달의 유전자 실시간 중합효소연쇄반응 분석 결과는 다음과 같다. Integrin alpha 5 (ITGA5), integrin beta 1 (ITGB1)과, protein tyrosine kinase 2 (PTK2)는 대조군에 비하여 EXP1과 EXP2에서 상향조절되었으며(P < 0.001), EXP2, EXP1, CON 순으로 유의미한 상향조절이 관찰되었다(P < 0.001, Fig. 2).
Chemokine signaling 신호전달의 유전자 실시간 중합 효소연쇄반응 분석 결과는 다음과 같다. G protein subunit beta 1 (GNB1)은 대조군에 비하여 EXP1과 EXP2 에서 상향조절되었으며(P < 0.001), EXP2, EXP1, CON 순으로 유의미한 상향조절이 관찰되었다(P < 0.001). G protein subunit gamma 2 (GNG2)은 대조군에 비하여 EXP2와 EXP1에서 상향조절되었고 EXP2와 EXP1사이에 유의한 차이는 없었다(P < 0.001, Fig. 3).
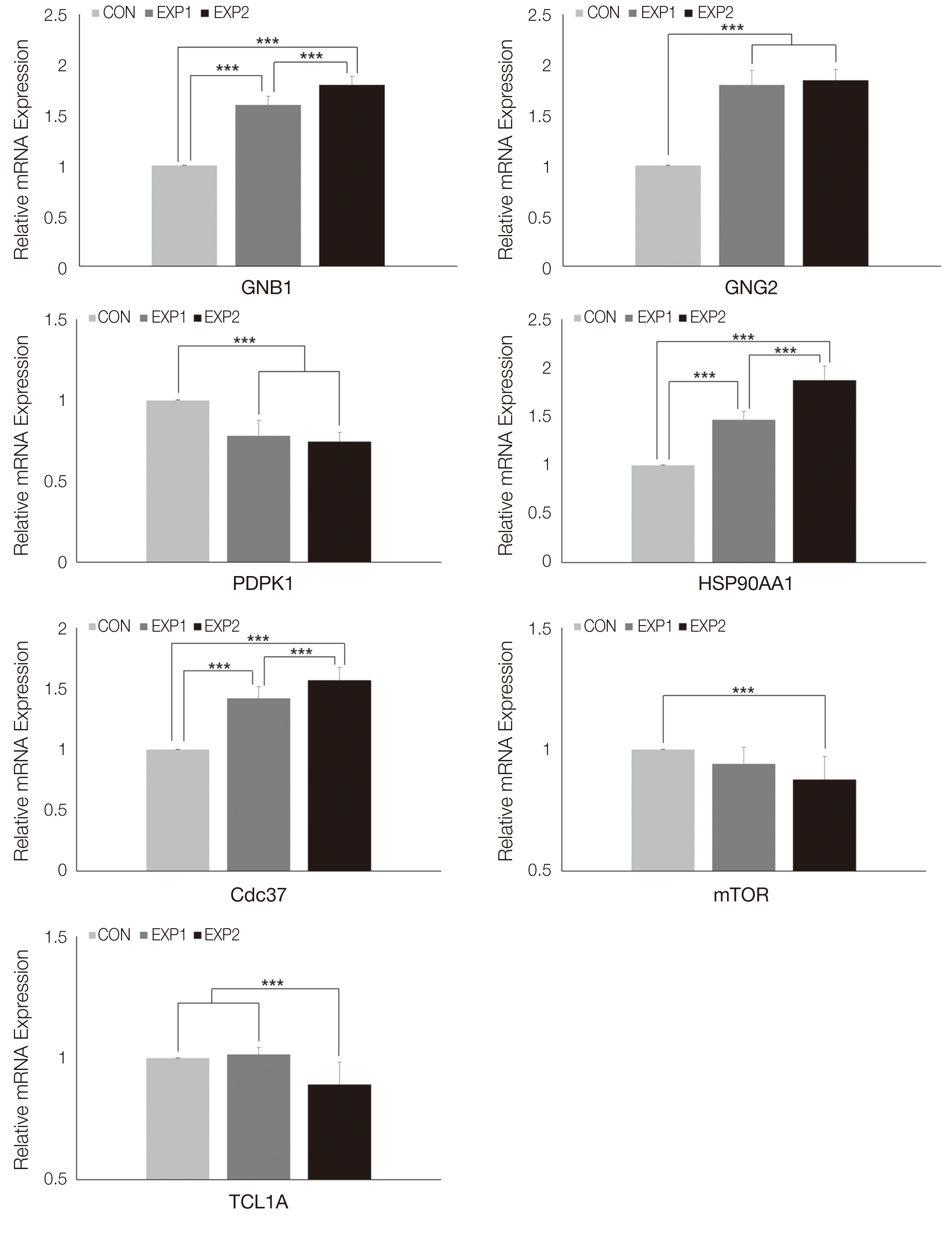
PI3K-AKT 신호전달에서 AKT 생성에 직접적으로 관여하는 유전자 실시간 중합효소연쇄반응 분석 결과는 다음과 같다. 3-phosphoinositol dependent protein kinase 1 (PDPK1)은 대조군에 비하여 EXP2와 EXP1에서 하향조절되었다(P < 0.001). Heat shock protein 90 alpha family class a member 1 (HP90AA1)과 cell division cycle control protein (CDC37)은 대조군에 비하여 EXP1과 EXP2에서 상향조절되었으며(P < 0.001), EXP2, EXP1, CON 순으로 유의미한 상향조절이 관찰되었다(P < 0.001). Mechanistic target of rapamycin kinase (mTOR)은 대조군에 비하여 EXP2에서 하향조절되었다(P< 0.001). T cell leukemia/lymphoma 1 a (TCL1A)는 대조군에 비하여 EXP2에서 하향조절되었다(P < 0.001, Fig. 3).
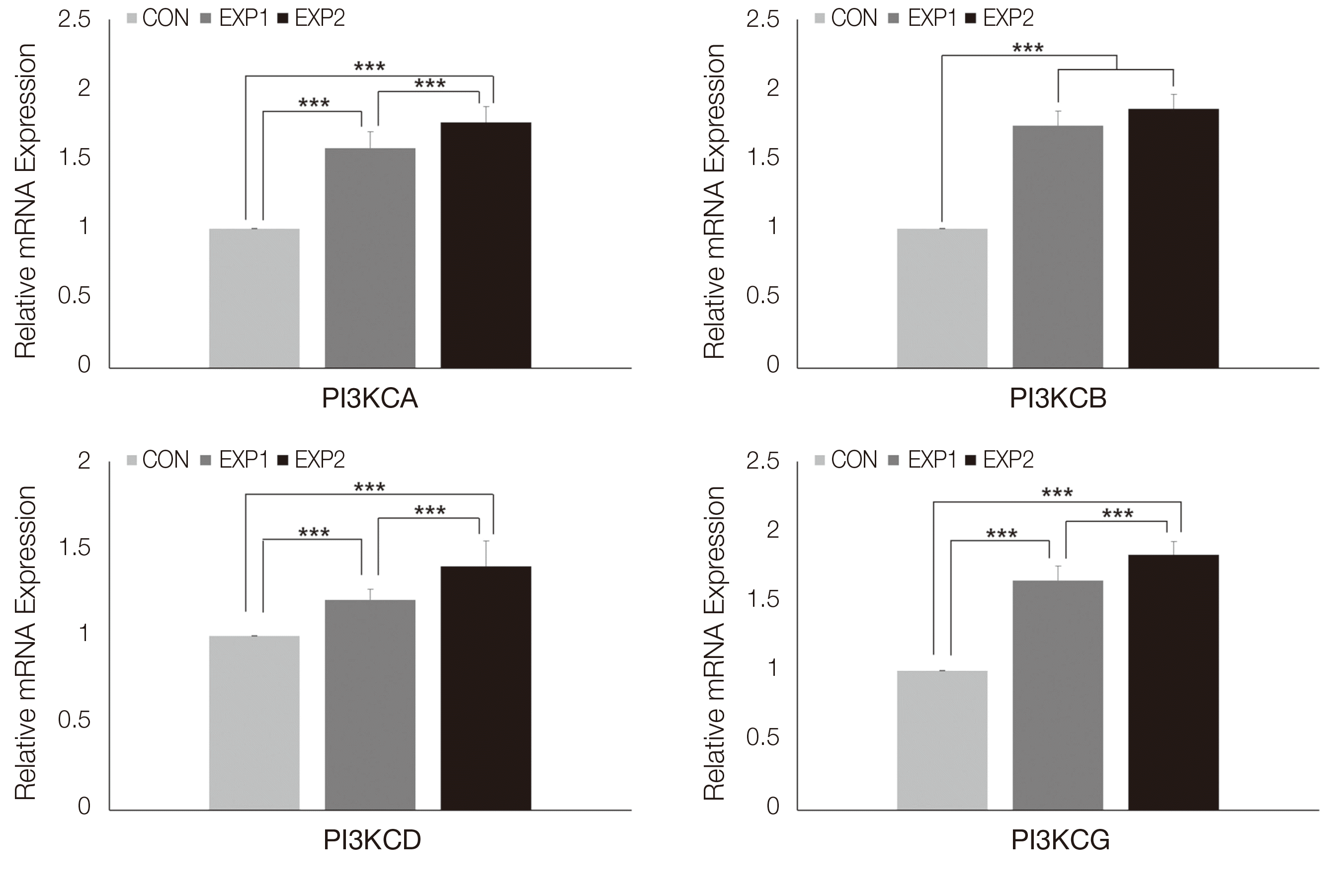
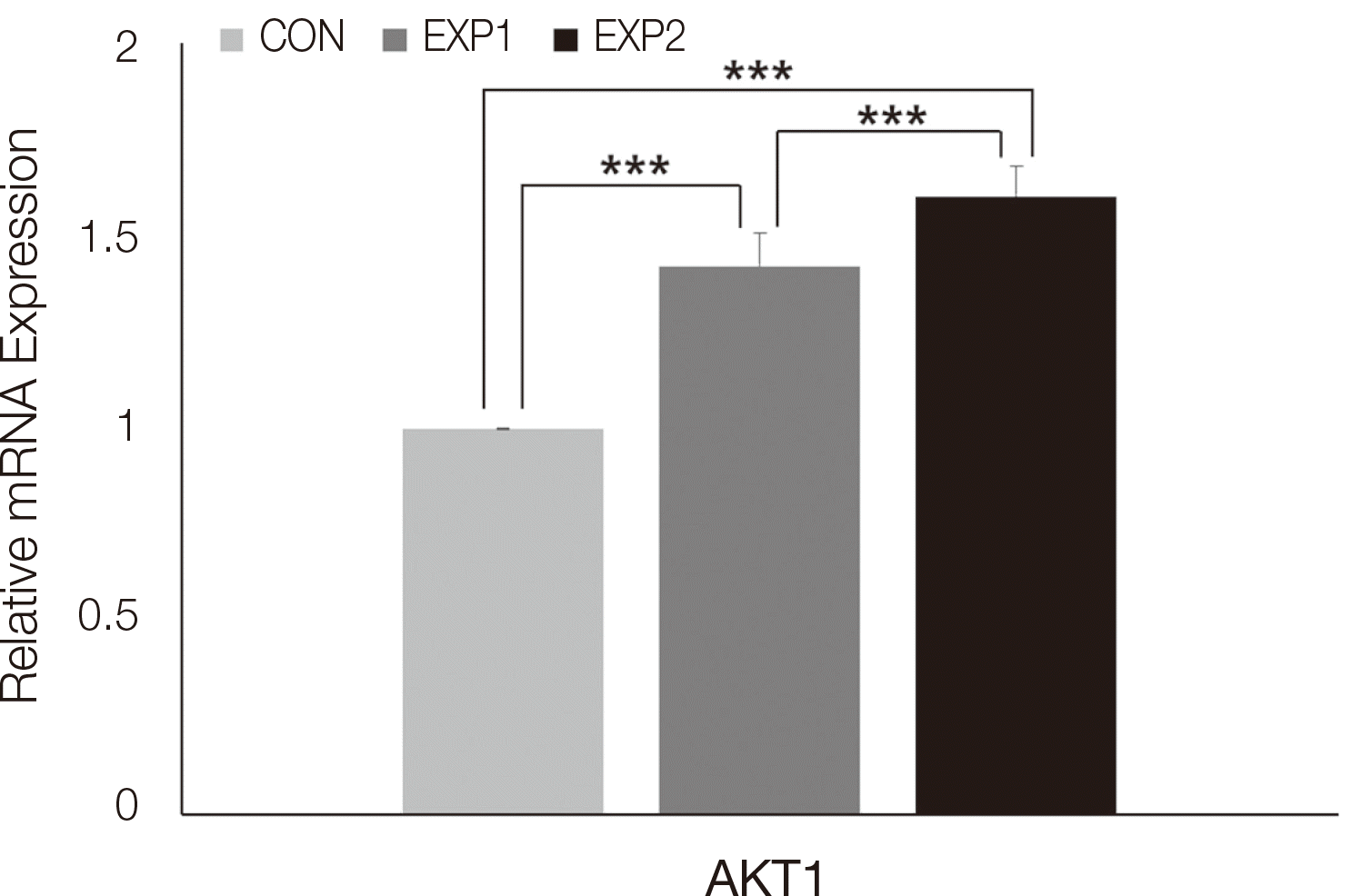
PI3K의 유전자 실시간 중합효소연쇄반응 분석 결과는 다음과 같다. Phosphatidylinositol-4,5-bisphosphonate 3-kinase catalytic subunit alpha (PI3KCA), Phosphatidylinositol-4,5-bisphosphonate 3-kinase catalytic subunit delta (PI3KCD)과, Phosphatidylinositol-4,5-bisphosphonate 3-kinase catalytic subunit gamma (PI3KCG)은 대조군에 비하여 EXP1과 EXP2에서 상향조절되었으며(P < 0.001), EXP2, EXP1, CON 순으로 유의미한 발현 증가가 관찰되었다(P < 0.001). Phosphatidylinositol-4,5-bisphosphonate 3-kinase catalytic subunit beta (PI3KCB)는 대조군에 비하여 EXP1과 EXP2에서 상향조절이 관찰되었고 EXP1과 EXP2 사이에 유의한 차이는 없었다(P < 0.001, Fig. 4). AKT1 유전자 실시간 중합효소연쇄반응 분석 결과, 대조군에 비하여 EXP1과 EXP2에서 상향조절되었으며(P < 0.001), EXP2, EXP1, CON 순으로 유의미한 상향조절이 관찰되었다(P < 0.001, Fig. 5).
AKT에 가장 유의미하게 영향을 미친 유전자를 확인하기 위해 다중회귀분석을 진행하였고 종속변수로 AKT1을 설정하였으며 본 연구에 사용된 유전자를 독립변수로 설정하였다. ITGB1, CD19, JAK3, PHF11, PDPK1, GNB1이 AKT1 표현에 영향을 주는 요소로 선택되었다(P < 0.001). 가장 영향력 있는 유전자는 ITGB1으로 확인되었다(P < 0.001, Table 2).
Table 2
The influential factors on the expression of AKT1 after administration of the e system to induce sequential release of TCA and EGF, as determined by multiple regression analysis
| Dependent variable | Model | Regression Results | R | R2 | Sig.1) |
|---|---|---|---|---|---|
| AKT1 | 1 | AKT1 = 0.061 + 0.941·[ITGB1] | 0.999 | 0.998 | < 0.001 |
| 2 | AKT1 = 0.264 + 0.969·[ITGB1] - 0.213·[CD19] | 1.000 | 0.999 | < 0.001 | |
| 3 | AKT1 = 0.401 + 1.019·[ITGB1] - 0.197·[CD19] - 0.223·[JAK3] | 1.000 | 1.000 | < 0.001 | |
| 4 | AKT1 = 0.666 + 1.108·[ITGB1] - 0.184·[CD19] - 0.699·[JAK3] + 0.109·[PHF11] | 1.000 | 1.000 | < 0.001 | |
| 5 | AKT1 = 1.261 + 1.157·[ITGB1] - 0.244·[CD19] - 1.240·[JAK3] + 0.215·[PHF11] - 0.150·[PDPK1] | 1.000 | 1.000 | < 0.001 | |
| 6 | AKT1 = 1.122 + 0998·[ITGB1] - 0.169·[CD19] - 1.216·[JAK3] + 0.251·[PHF11] - 0.110·[PDPK1] + 0.124·[GNB1] | 1.000 | 1.000 | < 0.001 |
![]()
Go to :
고찰
세포생존과 증식에 중요한 조절요소로 기능을 하는 것은 AKT이다.15 AKT는 보존된 domain 구조를 가지고 있고, 특정 pH domain, 중앙 kinase domain, signaling 분자들과 상호관계를 조절하는 carboxyl-terminal 조절 domain을 지니고 있다.15 AKT와 함께 PI3K도 성장인자와 다른 세포 외 자극들에 의해 반응하는 signaling 신호전달 안에서 중요한 역할을 담당하고, 영양대사, 세포성장, 세포사멸, 세포생존을 포함한 다양한 세포 기능을 조절하는데 영향을 미친다.15 선행예비연구들에서 HGC기반 TCA-와 EGF-담지형 나노방출제어시스템을 투여 시 단백질합성, 세포증식, 세포생존 등에 관련된 신호전달에서 상향조절되는 유전자들이 보고되었으며, 본 연구에서는 특별히, TCA와 EGF 투여 시 인간치은섬유아세포의 PI3K-AKT 신호전달에서 유전자 발현의 변화를 알아보았다. IGFR과 EGFR은 receptor tyrosine kinase (RTK)로 통칭될 수 있으며, 이러한 RTK들은 성장인자와 같은 polypeptide ligand와 결합하여 세포 반응을 조절하는 signaling cascade를 촉진한다.16 IRS는 인산화된 IGF1R에 의해 활성화되고 IRS는 PI3K의 활성에 도움을 주는 scaffold 역할을 한다.17 본 연구에서 IGF1R, EGFR, IRS1의 real time-PCR 분석 결과 EXP1과 EXP2에서 모두 유의하게 발현이 증가되었으며, EXP2에서 증가된 발현량이 EXP1보다 많았고, 이러한 유전자들의 발현량 증가는 PI3K와 AKT의 활성 증가를 이끌 수 있음을 의미한다. EXP1에서 증가된 발현량이 EXP2보다 적은 것은 EXP1에서 TCA의 독성이 영향을 미쳤을 것으로 생각된다. TLR은 선천 면역 센서 중 하나로 pathogen-associated molecular patterns (PAMPs)를 인식한다. TLR은 염증반응을 일으키며 PI3K를 활성화시킨다.18 TLR2는 EXP2에서만 유의미하게 발현량이 증가되었고 TLR4는 EXP2와 EXP1에서 유의미한 발현량 증가가 모두 관찰되었다. Rac1은 EXP1과 EXP2에서 모두 유의미하게 발현이 증가되었으며 EXP2에서 증가된 발현량이 EXP1보다 많았다. EXP1에서 TLR2는 유의미한 발현량 증가를 보이지 않았으나 TLR4에서는 유의미한 발현량 증가를 보였고 EXP2에서 TLR2와 TLR4에서 모두 유의미한 발현량 증가를 보였으므로 TLR2와 TLR4에 의해 활성화되는 Rac1은 EXP2에서 더 많은 발현량 증가를 했다고 유추할 수 있다. SYK는 BCR이 항원을 인지한 이후 B 림프구의 활성에 중요한 유전자이다. BCR에 의한 SYK의 군집은 이후 PI3K를 포함하는 signaling의 활성에 필수요소이다.19 CD19는 BCR과 떨어져 있지만 활성화된 B cell 표면과 연관성이 있어 유사한 측면을 보이고 SYK의 tyrosine 인산화와 연관이 있다.20 활성화된 SYK와 같은 protein tyrosine kinase (PTK)는 PHF11과 같은 세포질 수용체 단백질을 인산화21하고 PHF11은 BCR 자극 이후 인산화되어 PI3K 활성에 영향을 미친다.22 본 연구에서 EXP1과 EXP2에서 BCR과 CD19는 유의미한 변화를 보이지 않았고 SYK와 PHF11은 EXP2에서만 유의미한 발현량 증가를 보였다. 세포표면에 존재하는 BCR은 항원에 반응하여 활성화되는 단백질이지만 본 연구에서는 항원에 관한 실험이 없기 때문에 유의미한 변화를 보이지 않은 것으로 추측되며, BCR의 활성에 따라 CD19 의 활성도 변화하기 때문에 CD19도 유의미한 변화를 보이지 않았다. 세포질의 SYK와 PHF11은 EXP2에서 발현량이 증가되었는데 EXP1에서는 TCA의 독성이 영향을 미친 것으로 보인다. IL2RG는 골수의 미성숙 혈액 생성 세포의 표면에 위치한 단백질로 사이토카인 수용체이다. IL2RG의 활성은 IL2 signaling에 필요하며 tyrosine phosphorylation을 야기하여 Janus family tyrosine kinases (e.g. JAK1, JAK3)를 활성화시킨다.23 JAK가 인산화되면 결과적으로 PI3K 신호전달을 활성화시킨다.24 본 연구에서 EXP1과 EXP2에서 IL2RG 유의미한 변화를 보이지 않았고 JAK1과 JAK3는 EXP1과 EXP2에서 모두 유의미하게 발현량이 증가되었으며 EXP2에서 증가된 발현량이 EXP1보다 많았다. Extracellular matrix (ECM)이 integrin receptor (e.g. ITGA5, ITGB1)에 의해 군집되면 Focal adhesion kinase (FAK)(e.g. PTK2)는 integrin에 반응하여 인산화되며 PI3K 신호전달의 활성에 영향을 준다.25 본 연구에서 ITGA5, ITGB1, PTK2의 real time-PCR 분석 결과 EXP1과 EXP2에서 모두 유의미하게 발현량이 증가되었으며 EXP2에서 증가된 발현량이 EXP1보다 많았다.
세포 외의 chemokine, hormone 등이 세포 표면의 G-protein-coupled receptor (GPCR)과 결합하면 세포내의 G protein (e.g. GNB1, GNG2)의 상호관계를 이끌고 신호를 전달하게 하여 PI3K와 같은 downstream signal transduction 신호전달을 유발한다.26 본 연구에서 G-protein 중 GNB1과 GNG2의 실시간 중합효소연쇄반응 분석을 통해 EXP1과 EXP2에서 발현량이 증가된 것을 관찰하였다. PI3K의 형성은 PDPK1과 AKT의 membrane-based colocalization를 촉진시킨다. PDPK1은 자신의 T-loop를 인산화시키는 능력으로 인해 세포 안에서 구성적으로 활성화된다. PDPK1가 membrane으로 이동하면서 mTOR2와 함께 AKT1의 활성화를 돕는다.27 TCL1A 또한 AKT의 Pleckstrin homology domain에 결합하여 AKT 활성에 영향을 준다.28 그러나 본 연구에서는 PDPK1, mTOR, TCL1A는 EXP1과 EXP2에서 발현량이 감소되었다. TCA와 EGF 투여 시 발현량이 감소하는 것에 대해서는 추가적인 연구 및 고찰이 필요하다. HSP90AA1과 CDC37은 복합체로서 세포 내 AKT와 관련이 있다. HSP90AA1은 복합체 내에서 AKT의 안정성을 위해 필요하다.29 HSP90AA1과 CDC37은 EXP1과 EXP2에서 모두 유의미하게 발현이 증가되었으며 EXP2에서 증가된 발현량이 EXP1보다 많았다. PI3K-AKT signaling 신호전달에서 여러 신호전달의 단백질로부터 활성화된 PI3K는 AKT를 활성화시키고 AKT는 세포생존, 성장, 증식, 세포이주 및 혈관생성에 기여한다.2,3 EXP1과 EXP2에서 PI3KCA, PI3KCB, PI3KCD, PI3KCG, AKT1 모두 발현이 증가되었다.
세포생존과 관련된 PI3K-AKT signaling 신호전달의 29개의 유전자에 대해 비교 분석한 결과 HGC기반 TCA-와 EGF-담지형 나노방출제어시스템이 PI3K-AKT 신호전달에서 여러 유전자의 발현량을 증가시키고 어떠한 유전자가 세포생존에 영향을 줄 수 있는지 확인할 수 있었다. 다만, 이러한 반응은 항상 일정하지 않으며 그 이유는 TCA의 세포독성인 것으로 추측된다. 상향 조절된 PI3KCA, PI3KCB, PI3KCD, PI3KCG, AKT1은 세포생존에 중요한 역할을 수행한다. AKT는 세포생존, 전이, 약물 내성, 대사, 방사선 저항성을 포함하는 주요 세포 전달 체계의 주단백질이며 활성화된 AKT는 다양한 세포 내 단백질을 인산화하며 세포생존, 성장, 증식, 세포이주 및 혈관생성에 영향을 미친다.3,4 이에 따라, AKT의 발현량 결과값을 종속변수로 설정하여 다중회귀분석을 실시하였다. 다중회귀분석 결과로써 가장 유의미한 영향을 주는 요소는 ITGB1이었다. ECM과 결합되어 PI3K 생성을 촉진하는 ITGB1이 세포생존 및 증식 등의 주요 세포 전달체계의 주단백질인 AKT 발현에 가장 영향력 있는 유전자라는 것이 본 연구에서 규명되었다.
Go to :
 XML Download
XML Download