

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Due to different types of treatments and resources, trigeminal pain is one of the most distressing health concerns around the world. It's evoked by a sensory stimulus of the trigeminal branches in the head, face, and mouth structures through the initial processing in the trigeminal nucleus caudalis (Vc) [12].
Noxious stimulation and peripheral inflammation lead to a subsequent release of inflammatory mediators from the presynaptic trigeminal neuronal terminals. Induced cyclooxygenase-2 (COX-2) protein expression in the trigeminal tissue has also been reported. COX-2 is an inducible enzyme and commonly considered as a source of prostaglandin, induced by inflammatory stimuli. It has been demonstrated that COX-2 overexpression can induce hypersensitivity and excitability of nociceptor sensory terminals [34].
Brain-derived neurotrophic factor (BDNF), a key neurotrophin within the central nervous system, plays critical roles in neural survival and plasticity [5]. Various distributional studies of BDNF and BDNF receptors also indicated that they are strongly expressed in nociceptive fibers in the spinal cord and dorsal root ganglion, as well as the trigeminal ganglion and nuclei [67]. Interestingly, it has been shown that trigeminal pains can alter the synaptic releases of BDNF [8]. However, the modulating effects of BDNF on nociceptive pathways, and BDNF expression in inflamed tissues, are controversial.
Anti-nociceptive and pro-nociceptive, as well as different levels of BDNF expressions, in an inflammatory condition, influence trkB and trkC neurotrophin receptor agonists on thermal nociception in behavioral and electrophysiological studies [7910].
Orexin peptides (orexin-A and orexin-B), originally found in the lateral hypothalamus, stimulate target cells through two G-protein-coupled receptors, the orexin 1 and orexin 2 receptors, OX1R and OX2R, respectively. Orexin-A binds equally to both receptors, while orexin-B has a better affinity for OX2R [11]. Because of their broad central distribution, orexin peptides are involved in the regulation of various physiological functions, including pain and cognitive processes [1213]. Specifically, orexin A has a modulating role on trigeminal pain transmission. It has been shown that the posterior hypothalamic and intravenous administration of orexin-A can suppress trigeminal nerve firing in rats [1214]. Intra-Vc administration of orexin-A can also attenuate capsaicin-induced orofacial pain [15].
However, the underlying mechanisms of orexin-A inhibitory effect on trigeminal pain have not yet been fully investigated. In the present study, the effect of intra-Vc administration of OX1R agonist and antagonist on orofacial pain-induced changes in the COX 2 and BDNF expression was investigated in rats.
MATERIALS AND METHODS
1. Animals
Twenty-four adult male Wistar rats (230–270 g) were used. The rats were housed in a temperature-controlled room at 23 ± 1℃, with a standard 12 h light/dark cycle, and received food and water ad libitum. The experimental protocol was approved by the ethics committee of Kerman University of Medical Sciences, Kerman, Iran. All efforts were made to minimize distress.
2. Surgery
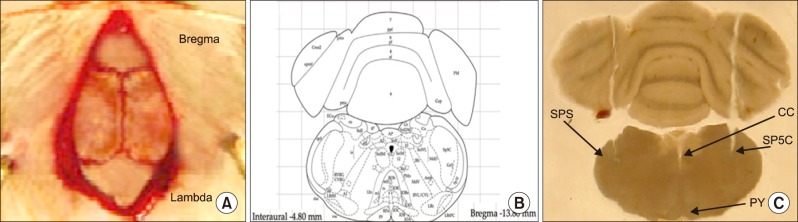
The rats were deeply anesthetized by an intraperitoneal (i.p) injection of a ketamine (100 mg/kg) and xylazine (10 mg/kg) mixture. The heads of the animals were fixed in a stereotaxic apparatus and a stainless steel guide cannula was implanted bilaterally in the skull (13.8 mm posterior to the bregma, 2.2 mm lateral from the midline and 7.5 mm deep to the cortical surface above the Vc) according to the Paxinos and Watson atlas of the rat brain [16]. The cannulas were attached to them using dental acrylic cement. The cannulas were then tightly closed by two stainless-steel screws.
After surgery, all the animals had at least 1 week of recovery before the microinjections. The location of the cannula in relation to skull coordinates, and a coronal section through the Vc, adapted from the atlas of Paxinos and Watson, are presented in Fig. 1. At the end of the behavioral experiment, methylene blue was injected via the guide cannula to verify the precise location of the cannula. If the cannula was misplaced, the rat's data was omitted from the analysis.
3. Drugs
Capsaicin was dissolved in Tween 80 and ethanol solution (1 ethanol, 1 Tween 80, 8 distilled water). Orexin-A and SB-334867-A (Tocris Bioscience, Bristol, UK) were dissolved in distilled water and dimethyl sulfoxide, respectively.
4. Microinjection
The rats were kindly handled and the microinjections were performed without any anesthesia. Orexin-A and SB-334867-A were administered into the Vc via the guide cannula (22-gauge) using an injection needle (27-gauge) connected by a polyethylene tube to a 1 µl Hamilton microsyringe. The injection needle was inserted 1 mm beyond the tip of the guide cannula. Infusions were delivered in a total injection volume of 2 µl (1 µl per side) over a period of 1 m. The injector was allowed to remain in the guide cannula for 30 s to avoid blocking.
5. Experimental groups
Four groups of rats (n = 6) were used: a normal control group (intact), a capsaicin-treated group which received intra-lip capsaicin injection with no cannulation; and orexin-A and SB-334867-A pretreated group, which received intra a Vc microinjection of orexin-A (100 pM) and SB-334867-A (80 nM) twenty minutes before capsaicin administration.
The dose of drugs and the number of animal per group were determined from previous studies [1517]. Besides, our previous study indicated that intra-Vc administration of orexin-A vehicle, distilled water, and DMSO as a vehicle of OX1Rs antagonist (SB-334867-A) had no effect on the strictness of capsaicin-induced orofacial pain. So in the current molecular trial we didn't evaluate vehicle groups [15].
The schematic drawing of the experimental procedures is presented in Fig. 2.
6. Pain induction
On the day of the test, rats were taken to the testing room and habituated for 30 m. Trigeminal pain was induced by a subcutaneous injection of capsaicin (100 µg/10 µl) with a 30-gauge hypodermic needle into the upper lip 20 m following the Vc administration of drugs.
Afterward, the animals were placed in a clear Plexiglas test box (30 × 30 × 30 cm), with a mirror placed at a 45° angle beneath the floor allowing for unbarred observation of the rats.
The nociceptive response was evaluated by measuring the number of seconds of a typical pattern of face rubbing behavior for 40 m, as previously described [15].
7. Immunofluorescence
Twenty-four hours after drug administration, the animals were anesthetized by i.p. injection of a ketamine/xylazine mixture (100 and 10 mg/kg, respectively) and perfused transcardially with 0.9% saline (100 ml) followed by 4% paraformaldehyde (500 ml).
The brains were post-fixed overnight in 4% paraformaldehyde. The sections were made from the rostral regions of the trigeminal nucleus caudalis (Vc), 200 µm in thickness, 2.2 mm lateral to the midline and 13.8 mm posterior to the bregma according to the Paxinos and Watson rat brain atlas. The samples were embedded in paraffin and then 2 µm sections were prepared from the Vc, containing paraffin blocks and were deparaffinized before immunostaining. The slides were hydrated through 100%, 90%, 70%, and 50% ethanol.
The sections then were treated for antigen retrieval using microwave treatment for 30 m in a citrate buffer (pH 6.0). They were washed in phosphate-buffer saline (PBS) for 3 m. The slides were dipped in hydrogen peroxide for 10 m and then exposed to anti-COX-2 or BDNF primary antibodies diluted at 1:500 (Santa Cruz Biotechnology, Inc, Dallas, TX) overnight in a humidity chamber.
The slides were washed in PBS and then incubated for 90 min with goat anti-mouse or goat anti-rabbit IgG-CFL 488 secondary antibodies at a dilution of 1:1000 (Santa Cruz Biotechnology, Inc, Dallas, TX) and washed with PBS. The sections were exposed to propidium iodide, at room temperature, and again washed with PBS.
Fluorescence microscopic images were captured with a digital camera, at a magnification of 20 times. All positive COX-2 or BDNF immuno-reactive cells in each Vc section were manually counted.
RESULTS
1. Pain assessment
The capsaicin injection into the upper lip evoked a significant pain response (P < 0.001). The cumulative time of pain response was 320.2 ± 22.63 s. Intra-Vc administration of orexin-A (100 pM), before the capsaicin injection could decrease the time of nociceptive behavior (214 ± 229) (P < 0. 05). However, in capsaicin plus SB-334867-A (80 nM)-treated rats, pain response (468.1 ± 35.64) was exaggerated as compared to the capsaicin-treated animals (P < 0.01) (Fig. 3).
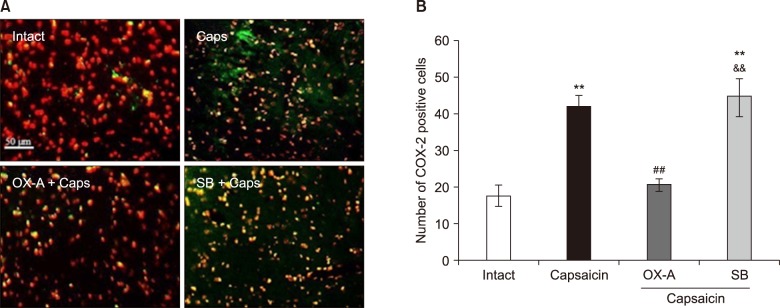
2. The effects of Vc administration of OX1Rs agonist and antagonist on COX-2 expression in the Vc of capsaicin-treated rats
As shown in Fig. 4, the number of COX-2 immuno-reactive cells was significantly increased in the capsaicin-treated group (42.01 ± 3.14) (P < 0.01). Pretreatment with orexin-A (100 pM) could diminish the capsaicin effects on COX-2 induction (P < 0.01). Furthermore, in SB-334867-A (80 nM) pretreated rats, the number of positive COX-2 cells (44.64 ± 5.39) was significantly higher than that in the control and orexin-A injected rats (P < 0.01) (Fig. 4).
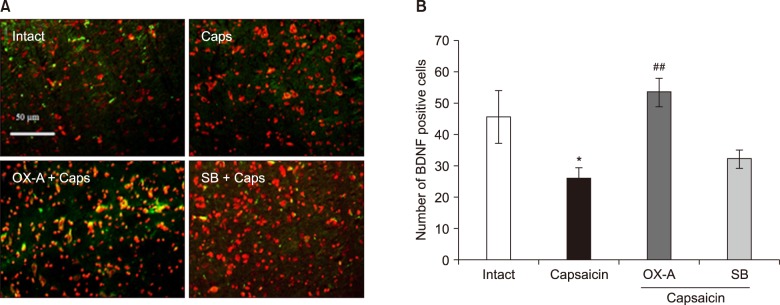
3. The effects of intra-Vc administration of OX1Rs agonist and antagonist on BDNF expression in the Vc of capsaicin-treated rats
In capsaicin-treated rats, the number of BDNF immuno-reactive cells (26.31 ± 3.33) was significantly decreased as compared with control rats (46.02 ± 8.39) (P < 0.05). However, pretreatment with orexin-A (100 pM), before the capsaicin injection, could prevent capsaicin-induced BDNF down-regulation (P < 0.001). Additionally, in SB-334867-A (80 nM) pretreated rats, before capsaicin injection, BDNF reactive cells (32.45 ± 2.87) were significantly decreased as compared to orexin-A injected rats (P < 0.01) (Fig. 5).
DISCUSSION
The present study showed that intra-lip capsaicin injection increased pain behaviors and COX-2 protein levels in the Vc. Interestingly, activation of OX1Rs in the Vc could reduce capsaicin-induced pain as well as COX-2 expression. There is growing evidence indicating the role of COX-2 in the modulation of trigeminal pain. Consistent with present results, Geo et al. showed that tooth movement can induce COX-2 expression in the Vc which correlates with an increase in nociceptive response [3]. Moreover, central inhibition of COX-2 has been shown to suppress formalin-induced temporomandibular joint inflammation in rats [18].
Capsaicin, as a potent activator of the TRPV1 channel, is known to promote inflammatory pain via the release of pro-inflammatory neuropeptides such as neurokinin A, substance P, and calcitonin gene related peptide (CGRP) in the trigeminal nerves [192021]. Interestingly, these neuropeptides have been shown to induce COX-2 expression in the trigeminal nucleus caudalis and trigeminal ganglion [2223].
In addition, capsaicin can produce hyperalgesia and pain hypersensitivity by alterations in the sensitivity of the peripheral terminals in response to normal and noxious stimulations [2425]. This situation is generally characterized by overexpression of COX-2 in the spinal and supra-spinal sites [426]. So, capsaicin-induced COX-2 upregulation may be somewhat correlated with the hyperalgesic and hypersensitivity effects of capsaicin. It may increase the sensitivity of Vc second order neurons which cause the enhancement and facilitation of synaptic transmission and inflammatory events [3].
The data indicated that Vc microinjection of orexin-A can inhibit capsaicin-induced COX-2 up-regulation, supporting the potent anti-inflammatory properties of orexin-A. These results are consistent with some in-vitro and in-vivo studies which indicate the anti-inflammatory and neuro-protective effects of orexin-A [2728]. It has been shown that Vc administration of orexin-A can diminish capsaicin-induced orofacial inflammation in rats [15]. Moreover, the inhibitory effects of orexin-A on A- and C-fibers in Vc neurons have been reported [1214]. In contrast, there is just one report that shows that the blockage of orexin receptors can inhibit trigeminal neuron activation via inhibiting the expression of protein kinase A and interleukin 1 beta in the spinal trigeminal nucleus [29].
Activation of the trigeminal nerves and associated glial cells are accompanied with the release of glutamate and various pro-inflammatory peptides that initiate and promote an inflammatory response within the spinal trigeminal nucleus [3]. It has been reported that orexin-A can decrease the expression of tumor necrosis factor alpha in murine microglia cells [30]. Surprisingly, it has been demonstrated that tumor necrosis factor alpha induces orexinergic neural toxicity [31]. In addition, orexin-A can inhibit neurogenic dural vasodilation via the inhibition of CGRP releases from trigeminal neurons [14]. So, the inhibitory effects of orexin-A on COX-2 induction may be partly mediated by suppression of the release of pro-inflammatory peptides from second order neurons in the Vc. However, further studies are needed to elucidate this important issue.
Inflammatory challenges are commonly associated with an increase in pro-apoptotic and oxidant markers which raise the risk of neural loss and deficiency [32]. Specifically, capsaicin has been shown to provoke cell apoptosis [33]. On the other hand, the anti-apoptotic properties of orexin-A have been reported [34]. Butterick et al. demonstrated that orexin-A decreases lipid peroxidation and apoptosis in embryonic rat hypothalamic cells [27]. Moreover, the orexin system has a protective effect against focal ischemia by the modulation of inflammatory responses in mice [35]. In the present study, the decreased expression of capsaicin-evoked COX-2 may be somewhat mediated by the antioxidant and anti-apoptotic properties of orexin-A. However, further studies are needed to explain the detailed role(s) and exact mechanism(s) of orexin-A in this regard.
The data showed that capsaicin-induced trigeminal pain decreased Vc expression of BDNF that was blocked by intra-Vc administration of orexin-A. BDNF as a widespread neurotrophin factor is known to be implicated in the regulation of nociceptive neurotransmission and excitability [6]. BDNF, mRNA, and protein are well expressed in primary trigeminal neurons [36]. Moreover, BDNF localization in the second-order sensory neurons of the Vc has been reported [37]. Peripheral irritation of the trigeminal nerves can induce changes in the expression of BDNF in the trigeminal ganglion [38]. Tarsa et al. reported that a greater extent of BDNF expression in the trigeminal ganglion was observed in mice than in rats, following pulp inflammation [39]. However, in-vitro administration of BDNF had no significant effect on capsaicin-induced CGRP releases from cultural trigeminal ganglion neurons [8]. According to data, capsaicin induces loss and probably defects in the transmission of BDNF, might reveal neural damage and toxic effects of capsaicin, which may be reflected by the increase in the number of intracellular reactive oxygen species as well as apoptosis [4041]. These events have been shown to reduce BDNF expression again [42].
The present data also indicated that a Vc microinjection of orexin-A could enhance BDNF levels in the Vc of capsaicin-treated rats. Harada et al. reported that an intra-hypothalamic microinjection of orexin-A can suppress BDNF down-regulation, induced by cerebral ischemic stress and neuronal damage in rats [43]. Moreover, orexins can increase the mRNA expressions of BDNF and neurotrophin-3 in rat primary cortical neuron cultures [44]. The current study seems to establish the possibility that there is a functional interaction between orexin-A and BDNF in the modulation of trigeminal pain. It may reduce the sensitivity of the trigeminal nerves via the alteration of the synaptic releases of sensory neuropeptides that are colocalized with BDNF, containing nociceptive fibers. Interestingly, in-vitro administration of BDNF can reduce release of substance P from the dorsal horn sensory neurons [45].
Our previous study indicated that orexin-A administration in the Vc of rats can suppress capsaicin-induced orofacial pain in a dose-dependent manner [15]. So, in the present study the most effective antinociceptive doses of OX1R agonist and antagonist were used to evaluate orexin-A effects on the induction of COX-2 and BDNF in the Vc of rats.
In conclusion, these results suggest that the inhibitory effects of orexin-A on trigeminal pain at least in part may be related to preventing the change in inflammatory and trophic agents including COX-2 and BDNF in Vc neurons. However, further studies need to clarify the exact mechanism (s) underlying the effects of orexin-A on trigeminal pain modulation.


 XML Download
XML Download