

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Trigeminal pains are one of the most widespread world health problems [12]. They can be evoked following motivation of the trigeminal branches in the head, face and mouth structures [3]. In this context, pulpal pain occurs as the result of activation of nociceptive fibers in the dental pulp. Some of sensory nerves in dental pulp express the transient receptor potential V1 (TRPV1) channel. Activation of the TRPV1 via an inflammatory agent such as capsaicin, an active principle component of chili peppers, has been associated with enhanced calcium influx and induced the subsequent releases of vasoactive peptides such as calcitonin gene-related peptide (CGRP) and substance P, leading to neurogenic inflammation [456].
Pain as a multidimensional experience has an interruptive effect on other physiological processes [78]. Specially, orofacial pain has been shown to have a significant association with increases in mood distress and cognitive deficiency [9101112]. Raoof et al. [9] also found the disruptive effect of capsaicin-induced pulpal pain on the spatial learning and memory of rats. Moreover, it has been indicated that trigeminal neuralgia can impair spatial learning and memory in rats [13]. In a human study, a cognitive deficit in patients suffering from temporomandibular disorder has been reported [14].
Regular exercise is known to be a beneficial factor for neuronal survival and mental health [1516]. Specially, studies on healthy volunteers and patients suffering neurodegenerative disorders indicated that exercise enhances cognitive function, likely through an improvement of synaptic plasticity, neuronal excitability, and neurogenesis [171819]. In addition, exercise is well known as a non-pharmacological treatment for pain suppression. It has been indicated that exercise can reduce pain symptoms via activation of the opioid and cannabinoid endogenous analgesic systems [2021].
Moreover, regular exercise suppresses both inflammatory and neuropathic pain in rodents [2122]. Clinical studies have also indicated that exercise therapy may be helpful in temporomandibular disorder managements [2324]. However, the effectiveness of physical exercise on other types of trigeminal pain has been less studied. So, the aim of the present study was to investigate the effect of regular treadmill running exercise on capsaicin-induced pulpal pain and pain-induced deficits in passive avoidance learning and memory in rats.
Go to : 
MATERIALS AND METHODS
1. Animals
In this study, adult male Wistar rats (230–270 g, 2–3 months) were purchased from the animal house of Kerman University of Medical Sciences. Animals were housed in a temperature-controlled room (22.0 ± 1℃) under a 12:12 light-dark cycle with free access to food and water. The experiment protocols were authorized by the Institutional Animal Ethical Committee of Kerman University, Kerman, Iran (code: IR.KMU.REC.1395.608).
2. Experimental design
The animals were randomly divided into four experimental groups (n = 6) as follows:
The control group, which received no injection.
The capsaicin group, which received a cotton pellet, dampened with capsaicin solution (100 µg) into a prepared cavity on the left mandibular incisor.
The exercise group, the animals were given exercise on a treadmill for 4 weeks
The exercise + capsaicin group, which received capsaicin solution following 4 weeks exercise training.
The rats' age in all the groups was the same at the time of the induction of nociceptive responses and behavioral assessment. Following regular exercise training, capsaicin was used for dental pulp stimulation. Nociceptive response was recorded for 40 m. Ten minutes after evaluation of nociceptive behavior; passive avoidance learning was conducted in a shuttle box apparatus.
3. Exercise protocol
The rats in the exercise groups were forced to run on a motorized driven treadmill for 5 days per week. The exercise sessions were performed between 9 AM and 12 AM. Animals were habituated with the treadmill apparatus one day before exercise training. A mild regular exercise protocol (4 week) was used.
There was a gradual increase in running speed and duration [25]. In the first two weeks, the animals ran 2 sessions per day (15 m each) at a speed of 10 m/min. In the third and fourth weeks, they ran 3 and 4 sessions per day (15 m each), respectively, at a speed of 15 m/min. A five-minute break was considered between the sessions.
4. Passive avoidance learning test
The apparatus that was used consists of two separate light and dark chambers with a grid floor. A poly methyl methacrylate (Plexiglass®) gate connected the chambers to each other. In the habituation trial, 30 m before the acquisition trial, the rats were located in the light chamber and permitted to enter the dark camber freely. During the acquisition phase, the rats were individually placed in the light chamber and after 5 s the guillotine gate was opened. Upon the entrance into the dark chamber, the guillotine door was lowered and a constant electric shock (0.5 mA, 50 Hz, 4 s once) was delivered through the grid floor.
After 30 s, the rats were returned to their home cage and 5 min later, the same course was repeated. If the animals did not enter into the dark room during 300 s, the acquisition of passive avoidance learning was considered successful. Otherwise, the rats received the shock a gain. The number of acquisition sessions was recorded.
Twenty-four hours later, in the retention test, the rats were placed in the light chamber and allowed to cross into the dark chamber. Step-through latency (STL) and time spent in the dark chamber (TDC) were recorded up to a maximum of 300 s.
5. Pain induction and management
Animals were familiarized with the experimental condition by placing them individually into the test box 30 min prior to experiments. Carbon dioxide (CO2) at a low concentration was used as an anesthesia because of its short onset and fast recovery times (total time of anesthesia was about 3 min) as well as its relative safety [2627].
After short-term anesthesia, a cavity (2 mm × 2 mm × 2 mm) was prepared in the gingival one-third of distal aspect of the left mandibular incisors using a small fissure bur in a high-speed handpiece with water coolant. With the help of magnification (2.5 ×), pulp exposure was prevented.
A small cotton pellet dampened with capsaicin solution (100 µg/10 µl) was left in the cavity under a light-cured glass-ionomer restoration. A capsaicin-evoked trigeminal nerve is a suitable laboratory method to investigate the mechanisms underlying orofacial pain. Capsaicin induces distinct and significant nociceptive behavior following intravibrissal pad administration in rats [28]. Additionally, it has also been reported that intradental applications of capsaicin is associated with considerable inflammatory pulpal pain [2930].
Upon pain induction, each rat was placed in the transparent box (30 × 30 × 30 cm) with a mirror situated at a 45° angle below the floor to detect unbarred observation of the animals. The rats' behavior was continually observed for 40 min (8 blocks of 5 min). In this period, the cumulative time of continuous shaking of the lower jaw and excessive grooming and rubbing of the mouth near to the procedure site was recorded as nociceptive behavior [928].
6. Statistical analysis
Statistical analyses were performed using SPSS, version 18.0 for Windows. Data are presented as mean ± standard error of mean.
One-way analysis of variance (ANOVA) followed up with Tukey's post hoc test was used to determine whether there are any statistically significant differences between the groups. Statistical significance was considered as P < 0.05.
Go to :
RESULTS
1. The effects of regular exercise training on capsaicin induced orofacial pain
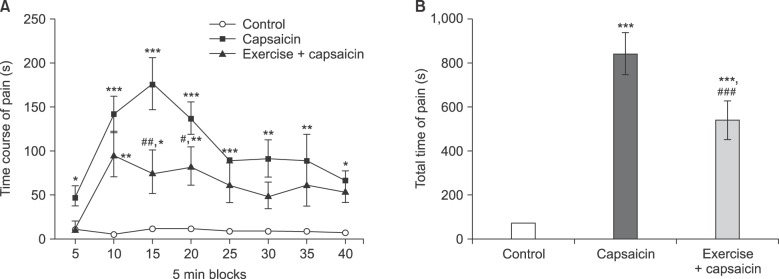
As shown in Fig. 1A, intradental application of capsaicin produced nociceptive behavior in rats. Regular exercise training could significantly attenuate capsaicin-induced pain at 15 (P < 0.01) and 20 min (P < 0.05) intervals after pain induction. Moreover, the total time of pain response was significantly decreased in the regular exercise group as compared to the capsaicin and intact groups (P < 0.001) (Fig. 1B).
2. Acquisition test
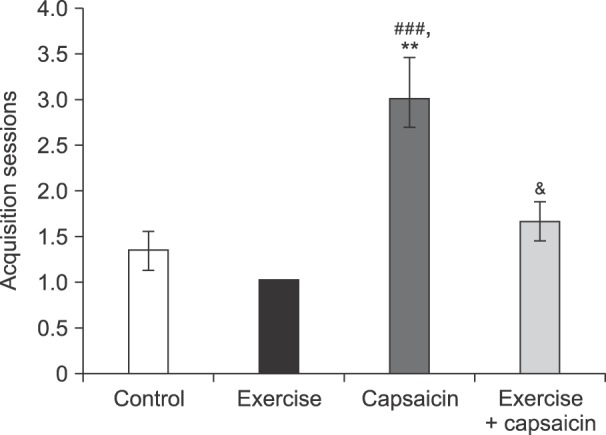
In capsaicin-treated rats, the number of trials required to achieve acquisition was significantly increased as compared to the control (P < 0.01) and regular exercise (P < 0.001) groups. Moreover, the increased number of trials induced by capsaicin application was reversed by regular exercise (P < 0.05) (Fig. 2).
3. Retention test
The results of the retention test indicate significant differences in the STL [F (3, 23) = 12.21, P = 0.001] and TDC [F (3, 23) = 7.77, P = 0.001] among the experimental groups (Fig. 3A and B).
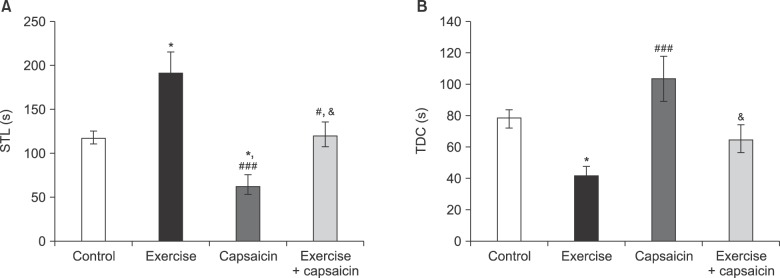 | Fig. 3Effects of capsaicin and regular exercise on (A) step-through latency (STL), and (B) the time spent in the dark chamber (TDC) in the retention test. Data are presented as mean ± SEM. *P < 0.05 versus control group. ###P < 0.001, #P < 0.05 versus exercise group. &P < 0.05 versus capsaicin group.
|
In capsaicin-treated rats, significant decreases in STL were observed as compared to the control (P < 0.05) and regular exercise groups (P < 0.001). Furthermore, regular exercise could suppress capsaicin-induced decreased STL (P < 0.05) (Fig. 3A).
In the capsaicin group, TDC was significantly increased as compared to exercise group (P < 0.05). Additionally, pretreatment with regular exercise caused a decreased TDC in capsaicin treated rats (P < 0.05) (Fig. 3B).
Go to :
DISCUSSION
The present study reveals that capsaicin-induced dental pulp pain impaired passive avoidance learning in rats, while regular running exercise attenuated the pain as well as memory deficiency induced by capsaicin. The positive effects of physical exercise on decreasing the symptoms of various types of pain have been reported using clinical and preclinical studies. Hung et al. [31] reported that treadmill exercise can improve spinal cord nerve injury in rats. They suggested that this beneficial effect may be related to down regulation of pro-inflammatory interleukin-6 (IL-6) and IL-a1, and up-regulating of anti-inflammatory cytokines such as IL-10. Moreover, regular aerobic exercise has been shown to reverse sensory hypersensitivity in a rat neuropathic pain model [21].
Additionally, while there is no study on the effects of exercise on pain arising from the tooth pulp, two recent studies revealed that exercise helps to mitigate temporomandibular pain [2324]. Almost all previous studies have indicated that exercise therapy can decrease pain symptoms in subjects suffering from pain. However, the present study was conducted to investigate the preventive effects of exercise on nociceptive behavior. We could find no experimental evidence for comparison with our results.
Capsaicin as a potent ligand of TRPV1 channels has been shown to induce neurogenic inflammation via the release and overexpression of sensory neuropeptides and pro-inflammatory cytokines [46]. These peptides might exaggerate pain responses and neuronal damage [3233]. There is growing consensus that physical exercise alters the levels of pro-inflammatory cytokines. Treadmill exercise can suppress the expression of tumor necrosis factor-α in the spinal cord of osteoarthritic rats [22]. Exercise may mitigate some of the spinal cord nerve injury related risk through acute disturbances of the inflammatory environment [34]. Surprisingly, a clinical study in patients with vestibular migraine by Lee et al. [35] revealed that exercise significantly suppresses the levels of pro-inflammatory cytokines through the inhibition of COX-2 activity. So, in the present study, it is likely that long lasting exercise exerts preventative effects on capsaicin-induced nociceptive responses via suppression of the induction of pro-inflammatory mediators as well as decreasing the sensitivity of trigeminal nociceptive nerves to capsaicin. Obviously more studies are needed to clarify this issue.
The effects of exercise therapy on endogenous pain-relieving peptides have been proposed [36]. It has been reported that exercise-induced analgesia is attenuated by naloxone, a u-opioid receptor antagonist [37]. Moreover, Stagg and colleagues reported that regular aerobic exercises suppressed pain hypersensitivity in rats through up-regulation of endogenous opioid peptides, beta-endorphin, and met-enkephalin, in the periaqueductal grey matter and rostral ventromedial medulla, the areas that are considered as the main brain centres involved in pain modulation [21]. Regular running exercise has also been shown to increase the serum and cerebrospinal fluid content of endogenous opioids [38]. Dietrich and McDaniel [39] stated that regular exercise decreases pain sensitivity via the endocannabinoid analgesic systems in rats. So, it is possible to assume that exercise can diminish capsaicin-induced pulpal pain through the activation of the endogenous analgesic system.
It is well established that physical exercise improves neuronal survival and synaptic plasticity, predominately by promotion of the expression of endogenous neurotropic factors and motivation of neurogenesis in memory-related areas [4041]. Here, as shown in Fig. 3, regular exercise could suppress capsaicin-induced passive avoidance learning and memory impairment in rats. It is well documented that cognitive dysfunction occurs in painful situations [42]. In particular, it has been reported that the spatial learning and memory of rats is impaired following acute intra-lip or chronic intra-dental capsaicin administrations [1029]. Capsaicin-evoked pulpal nociception has been shown to be associated with an increased Bax/Bcl2 ratio as well as raised caspase 3 activity in the hippocampus of rats [29]. Otherwise, it has been shown that regular exercise can prevent apoptosis and neuronal damage by decreasing the levels of reactive oxygen species and other measurable markers of oxidative stress in brain [4344]. In the present study, the possible modulatory effects of regular running exercise on the induction of apoptotic factors and strengthening of the antioxidant defence system, may be protective against capsaicin-induced cognitive deficiency. However, additional experimental studies are still required to elucidate this issue.
It has been shown that exposure to psychophysiological stressors can alter the severity of nociceptive responses. Exercise enhances the body's ability to respond to stress through changes in neurotransmitter secretions such as dopamine, serotonin, and some kind of hormones [4546]. However, it's not clear how much effect the physical stress due to exercise has on nociceptive responses. The issue would be helpful to understand the mechanism underlying the effects of exercise on pain behaviors.”
In the current study, the mechanism underlying the inhibitory effects of exercise on capsaicin-evoked pulpal nociception was not investigated. Pharmacological studies as well as evaluation of the impact of regular exercise on induction of pro-nociceptive mediators in the trigeminal sensory nuclear complex may help in clarifying the exact mechanisms of exercise-inhibited nociceptive reaction in response to pulp irritation.
According to the present data, regular exercise can decrease the severity of capsaicin-induced pulp nociception in rats; however, additional studies are required to confirm the clinical efficiency of regular exercise on modulation of pulpal nociception.
In conclusion, our findings indicate that regular exercise as a non-pharmacological treatment can decrease the severity of capsaicin-induced pulpal pain, and pain-related cognitive deficiency in rats. However, the exact mechanism by which this occurs remains to be clarified.
Go to :
 XML Download
XML Download