

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Nefopam, a centrally acting non-opioid agent, has been widely used in European countries, and it could have useful applications in clinical practice due to its analgesic mechanism, which differs from that of other analgesic agents. It has been proposed in anecdotal literature to act through inhibition of monoamine reuptake, but the underlying mechanism of action remains inconclusive [1-5].
A recent study, which investigated the role of specific subtypes of the monoamine system in the antinociception of systemic nefopam for phase 1 of the formalin test, demonstrated a significant involvement of the dopamine D2 receptor, but not of alpha-1 or alpha-2 adrenergic receptors, in the formalin test [6]. However, there have been no such reports regarding phase 2 of the formalin test. Most of the previous studies did not look into nefopam's mechanism of action at the spinal level even though the spinal cord is also an important site of action of monoamines which mediates descending pain modulation [7,8]. Furthermore, previous reports demonstrated the analgesic effect of spinal nefopam [9-11], but the overall role of the spinal noradrenergic system in producing nefopam's analgesic effect remains unclear.
Several reports have demonstrated the benefits of using nefopam in clinical settings, which include synergistic analgesic interaction with opioids, a preventive effect on postoperative shivering, and a lack of analgesic antagonism by the antiemetic 5-HT3 receptor (5-hydroxytryptamine receptor subtype 3) antagonist, ondansetron [9,12-15]. Although alpha-adrenergic agents are being widely employed in the perioperative period, the role of noradrenergic modulation in the analgesic effect of nefopam has not been fully addressed. Furthermore, a recent study also demonstrated the important role of the alpha-2 receptor in the anti-shivering effect of nefopam, suggesting the possibility that the alpha-2 receptor is involved in nefopam's mechanism of analgesia [16].
This study was performed to evaluate the role of the spinal noradrenergic system and alpha-1 and alpha-2 receptors in the antinociceptive effect of nefopam at the spinal level.
Go to : 
MATERIALS AND METHODS
1. Animals and intrathecal catheter implantation
Male Sprague-Dawley rats weighing 225-250 g were housed in a room with a constant temperature of 22-23℃ and an alternating 12-hour light/dark cycle. Free access to water and food was allowed. All experiments were performed according to the International Association for the Study of Pain Guidelines for the Use of Animals in Research.
A polyethylene-5 (PE-5) catheter was implanted into the intrathecal (i.t.) space to allow administration of the experimental drugs as described in the previous study [9]. Under anesthesia with inhalation of sevoflurane, the dorsal part of the neck was dissected to reveal the atlantooccipital membrane and a PE-5 catheter was introduced through the membrane. The catheter was then advanced caudally 8.5 cm to the level of the lumbar enlargement. The other end of the catheter was exteriorized to the top of the head and plugged with wire for drug administration. Any rats showing neurological deficits after the surgery were sacrificed immediately with an anesthetic overdose. Each rat was given a subcutaneous injection of 5 ml saline before the end of the surgery, and the animals were housed in individual cages following the procedure.
2. Nociceptive test and behavioral study
The animals were restrained in a cylinder and were then injected subcutaneously with 50 µl of 5% formalin into the center of the hindpaw using a 30 gauge needle. Following the intraplantar injection of formalin, the animals displayed a typical flinching response, which represents initial acute nociception (phase 1, acute pain) by direct stimulation of the peripheral nociceptors, and a following facilitated state of spinal dorsal horn neurons as well as peripheral sensitization (phase 2, facilitated pain) [17]. After an animal was injected, an observer counted the number of flinching responses at 1 min and 5 min (phase 1, 0-9 min), and at 5-min intervals between 10-60 min (phase 2, 10-60 min). Each counting of flinching responses was of 1-min duration. The observer who carried out the behavioral testing was blinded to the agents used for i.t. treatment.
3. Drugs
This experiment used the following drugs: prazosin (alpha-1 receptor antagonist), yohimbine (alpha-2 receptor antagonist), and nefopam hydrochloride (Acupan®). The vehicle for the drugs was saline. All of the drugs were administered using a hand-driven gear-operated syringe pump through an i.t. catheter in a volume of 10 µl, followed by flushing the catheter with 10 µl of saline. In addition, 6-hydroxydopamine (6-OHDA) was used for selective denervation of spinal noradrenergic terminals and depletion of noradrenaline (NA) in the spinal cord [18].
4. Depletion of spinal noradrenaline
Spinal catecholamine depletion was achieved using 6-OHDA (Sigma-Aldrich, St. Louis, MO, USA), which has been known to be taken up into adrenergic nerve fibers in the spinal cord and to destroy the fibers [18]. Intrathecal administration of 6-OHDA (20 µg/20 µl) or vehicle (control) was performed 3 days after surgery for catheter implantation for the behavioral study. Depletion of spinal NA was confirmed using liquid chromatography-tandem mass spectrometry (LC-MS/MS) for the dorsal half of the lumbar enlargement [19]. Using a stock solution of norepinephrine (Sigma-Aldrich, St. Louis, MO, USA), a calibration curve was obtained. Separation and detection were performed on a high-performance liquid chromatography (HPLC) system (Shimadzu HPLC system, Shimadzu Corp., Kyoto, Japan) coupled to a mass spectrometer (API 4000 Q TRAP, Applied Biosystems, Foster City, CA, USA). We identified the full scan of mass spectra and the product ion mass spectra of NA. The mass transition of m/z (mass-to-charge ratio) 234.233 to m/z 216.100 was used for NA. AB Sciex Analyst software (version 1.5.2) was used for data integration.
Three days after 6-OHDA injection, the animals' motor function was assessed by the righting and placing-stepping reflexes. The pinna reflex and corneal reflex were also examined to test for sensory deficits.
5. Experimental protocol
The dose of i.t. nefopam, 30 µg, was chosen according to our laboratory's previous work demonstrating its antinociceptive effect on flinching behavior in the formalin test [9]. The influence of spinal cord NA depletion on the antinociceptive effect of nefopam against formalin-induced pain was evaluated. Three days after i.t. catheter implantation, 6-OHDA or saline was administered through the catheter to deplete spinal NA. The change in antinociceptive activity of i.t. nefopam was examined 3 days thereafter.
Antagonism to the effect of nefopam by prazosin and yohimbine was also examined to further elucidate the mechanism of action of nefopam. Animals were intrathecally administered prazosin (3 µg) or yohimbine (10 µg), doses which have no effect on nociception as determined by a pilot experiment of our laboratory, or vehicle 10 minutes before i.t. nefopam followed by formalin injection [20]. Each group for behavioral and pharmacological study consisted of 8 animals, and each group for LC-MS/MS for NA measurement used 6 animals.
6. Statistical analysis
Data for flinching responses of each group are expressed as mean ± SEM, and differences among groups or treatments were analyzed with one-way ANOVA followed by Tukey's comparison. Data for NA concentrations are expressed as mean ± SD and analyzed by Student's t-test. A value of P < 0.05 was considered to indicate statistical significance.
Go to :
RESULTS
1. Effects of intrathecal nefopam on formalin-induced pain
Animals treated with i.t. vehicle showed typical biphasic flinching responses after formalin injection into the hindpaw. Administration of nefopam (30 µg) 10 minutes prior to the formalin injection significantly reduced the flinching responses in both phases of the formalin test compared to the control group (Fig. 1).
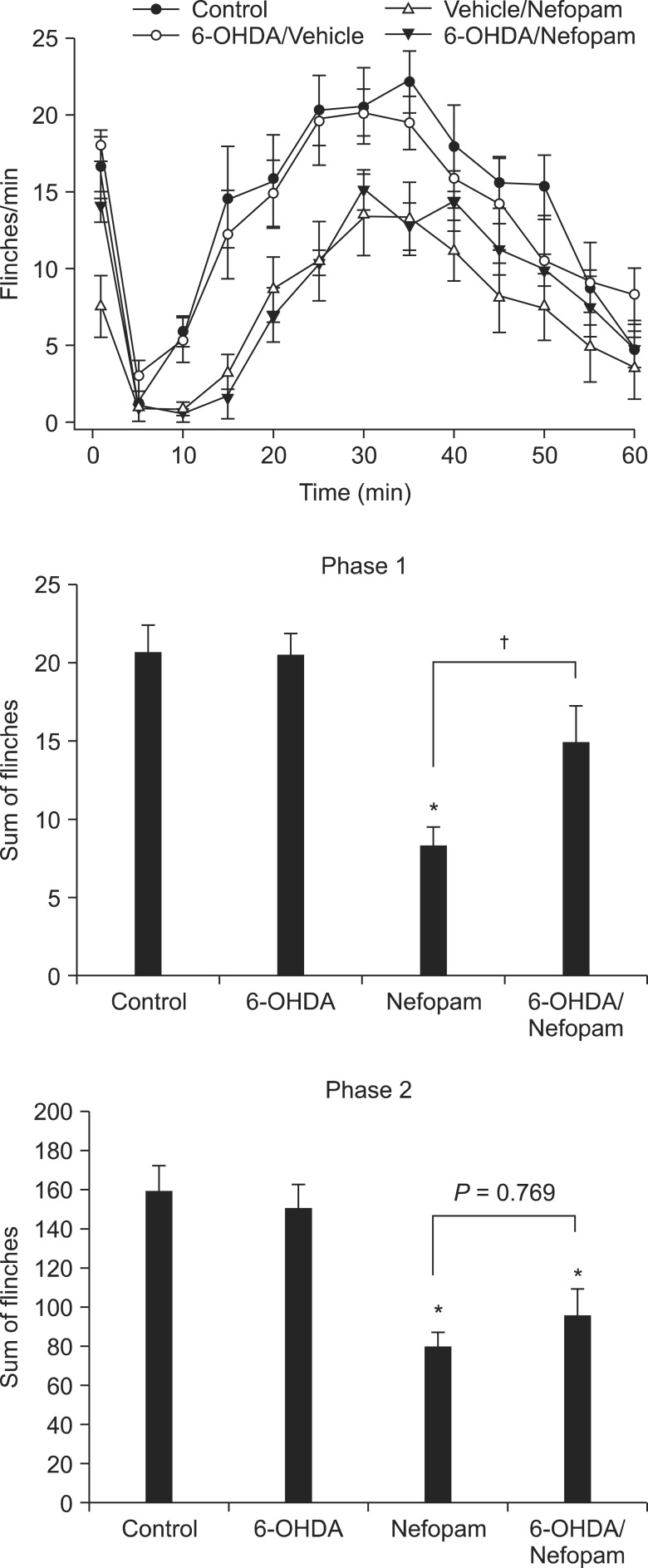 | Fig. 1Effects of intrathecal 6-OHDA treatment on flinching behavior and antinociception produced by intrathecal nefopam in the formalin test. Treatment with intrathecal 6-OHDA, 3 days prior to the formalin test, does not alter the flinching responses in either phase. However, 6-OHDA treatment attenuates the antinociceptive effect of i.t. nefopam significantly during phase 1, but not phase 2. *P < 0.01 when compared to control, †Indicates P < 0.01.
|
2. Influence of NA depletion on the antinociceptive activity of intrathecal nefopam
Treatment with 6-OHDA significantly reduced the content of NA in the spinal cord compared to controls (P < 0.01, naïve; 82.01 ± 14.48 ng/ml vs. 6-OHDA-treated; 15.01 ± 4.96 ng/ml). Depletion of spinal NA did not impair the motor or sensory function of the animals. The flinching responses of the 6-OHDA treatment group were not different from the controls in either phase, as previously shown in a study using a tonic nociceptive modality similar to phase 2 of the formalin test [21]. The antinociceptive effect of i.t. nefopam shown in vehicle-pretreated rats was significantly reduced in 6-OHDA-pretreated rats during phase 1, but not during phase 2 (Fig. 1).
3. Antagonism of antinociceptive activity of intrathecal nefopam by alpha-adrenoreceptor antagonists
During phase 1 of the formalin test, the sum of flinches in animals treated with i.t. nefopam following prazosin was not statistically different from that of animals treated with i.t. nefopam alone, although the difference was very close to the level of statistical significance (P < 0.06, Fig. 2). However, treatment with yohimbine prior to i.t. nefopam partly, but significantly, reduced the antinociceptive effect of nefopam in phase 1. The sum of flinches for animals that received both yohimbine and nefopam was significantly different from that of the control group.
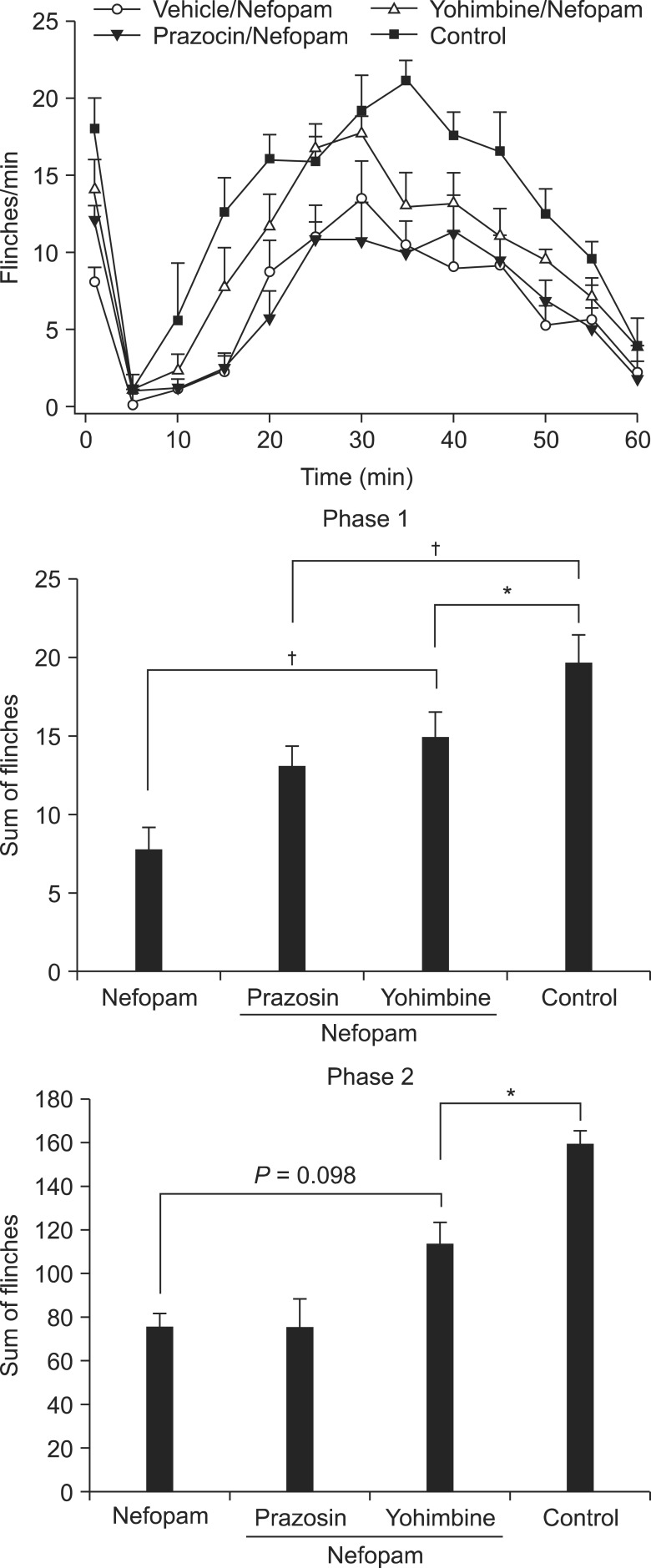 | Fig. 2Effects of antagonism of the alpha-2 adrenoreceptor on the antinociceptive effect of i.t. nefopam. Yohimbine, an antagonist of the alpha-2 receptor, but not the alpha-1 receptor, reduces partially, but significantly, the antinociceptive effect of i.t. nefopam during phase 1. During phase 2, the flinching responses of nefopam-treated animals are not different from those of the yohimbine plus nefopam-treated group. *Indicates P < 0.05, †P < 0.01.
|
In contrast, during phase 2, pretreatment with prazosin or yohimbine failed to reverse the antinociceptive effect of nefopam.
Go to :
DISCUSSION
This study demonstrated that depletion of spinal NA attenuated the analgesic action of i.t. nefopam during phase 1, but not during phase 2, of the formalin test. Of spinal alpha-adrenoreceptors, the alpha-2 receptor rather than the alpha-1 receptor plays a major role in the antinociceptive effect of i.t. nefopam during phase 1, but other mechanisms seem to be contributing to the analgesia by nefopam. During phase 2, neither the alpha-1 nor the alpha-2 receptor is involved in the mechanism of nefopam's analgesia.
Pretreatment with 6-OHDA produced a significant reduction of nefopam's antinociceptive effect during phase 1. In line with this result, a significant reduction of the effect of nefopam due to pretreatment with 6-OHDA was also observed in a previous study using acute thermal pain in rats [4]. During phase 2, however, no difference in the antinociceptive effect of nefopam was produced by 6-OHDA pretreatment. This finding could be in support of the important involvement of monoamine systems other than the noradrenergic system, including spinal serotonergic systems, in the effect of nefopam [6,22,23].
Monoamines of the spinal cord including serotonin or noradrenaline have been known to mediate descending pain modulation [7,8]. Several studies have demonstrated the involvement of serotonergic or noradrenergic descending pathways in the analgesic action of nefopam [3-5,23]. However, discrepant results were presented by these studies, among which the animal species, pain modalities, agents used for monoamine depletion, and routes of administration differed. These inconsistent findings could be related to the notion that descending modulation mediated by serotonin or noradrenaline, the main target of nefopam, could be inhibitory or facilitatory depending on the pain stimuli [7,8]. In addition, actions other than inhibition of monoamine uptake have been found to be a part of the analgesic mechanism of nefopam in previous studies. These actions involve the transient receptor potential vanilloid subtype 1, voltage-sensitive calcium channels, and the glutamatergic system [24-26]. Such multiple mechanisms of nefopam might be a reason for the different results regarding the role of monoamines in the antinociception of nefopam.
In a previous study, blockade of neither the alpha-1 receptor nor the alpha-2 receptor through systemic administration of antagonists significantly altered the antinociceptive effect of nefopam during phase 1 of the formalin test [6]. Inconsistent with this previous study, in the current study, the alpha-2 receptor, but not the alpha-1 receptor, was shown to mediate the antinociceptive effect of i.t. nefopam during phase 1. To the best of my knowledge, this is the first study to examine the role of spinal alpha-1 and alpha-2 receptors in the antinociception of nefopam during phase 2 of the formalin test. Blockade of the alpha-2 receptor produced only a modest, but insignificant, increase of flinching responses in phase 2 compared to nefopam treatment in the current study, while the important role of the alpha-2 adrenoreceptor in analgesia has been demonstrated in various pain models at peripheral, spinal, or systemic levels [27-29]. In contrast with the alpha-2 receptor, spinal alpha-1 receptors have been known to play a minor role mainly by facilitating the excitability of alpha-motor neurons of the ventral horn, in which alpha-1 receptors are widely distributed [30-32]. The activity of the alpha-1 receptor was even reported to be related to the reduction of the analgesic activity produced by alpha-2 receptor agonists [33]. The results of the current study also indicate a less important role of spinal alpha-1 receptors compared to alpha-2 receptors. In addition, the observed lack of changes in flinching responses of spinal NA-depleted animals could be related to the opposite effects of alpha-1 and alpha-2 adrenoreceptors as well as major roles of other monoamine systems.
One of the main beneficial characteristics of nefopam observed in previous studies could be a synergistic or adincditive analgesic interaction with various other analgesic agents luding morphine, paracetamol, or ketoprofen [34-36]. Interestingly, the anti-shivering effect of nefopam was reported to be related to the alpha-2 receptor in a recent animal study [16]. Therefore, although nefopam's analgesia was not completely reversed by blockade of the alpha-2 receptor, the results of this study could provide the initiative for further investigation into the possible interaction of nefopam with alpha-2 agonists, as both are widely used in the care of patients during the perioperative period.
In conclusion, spinal noradrenergic modulation plays an important role in the antinociceptive effect of i.t. nefopam against formalin-elicited acute initial pain, but not facilitated pain, and this action involves the spinal alpha-2 rather than the alpha-1 receptor.
Go to :
 XML Download
XML Download