

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Paclitaxel (Taxol) is a widely used chemotherapeutic agent for the treatment of solid cancers. Its anti-neoplastic activity is known to increase microtubule stabilization [1]. However, it has several side effects, including peripheral neuropathy and allodynia, which lead to poor compliance and discontinuation of the therapy [2,3]. The induction of peripheral neuropathy is the main dose-limiting side effect of paclitaxel treatment [4]. Although the exact mechanism of paclitaxel-induced neuropathic pain is not fully understood, several reports have shown that the derangement of nerve fiber endings may play a role in the development of this pain [5,6]. Sensory neurons and satellite cells located at the dorsal root ganglion (DRG) are altered by paclitaxel treatment [7-9]. In addition, it is well known that apoptosis is evident in DRG neurons following nerve injury, which may induce allodynia [10-12]. Neurotoxic effects in DRG cells after paclitaxel treatment have also been reported [7,8].
Ethyl pyruvate (EP) is a stable, lipophilic derivative of pyruvic acid with known anti-inflammatory and antioxidant properties. It has been reported that administration of EP may reduce brain and spinal cord injuries in cerebral and spinal cord ischemic/reperfusion-induced injury models and spinal cord compression injury models [13-15]. EP is also known to inhibit apoptosis in various models of organ injury [15-17] and have analgesic effects on spinal nerve ligation- or inflammation-induced painful behaviors [18-20]. For example, we recently reported that EP has a strong anti-allodynic effect that acts through the inhibition of tumor necrosis factor (TNF)-α expression and apoptosis in the DRG after spinal nerve ligation injury [18]. However, the analgesic effect of EP has not been evaluated carefully in chemoneuropathic pain, such as paclitaxel-induced allodynia.
Therefore, the specific aim of the present study was to evaluate the analgesic effect of EP on mechanical allodynia and apoptosis in DRG cells after paclitaxel administration.
Go to : 
MATERIALS AND METHODS
This study was reviewed and approved by the Institutional Animal Care and Use Committee (IACUC) of the Asan Institute for Life Sciences, Asan Medical Center. The committee follows the Institute of Laboratory Animal Resources (ILAR) guide.
1. Experimental animals
Male Sprague-Dawley rats (160-180 g) were housed 3 per cage in a room maintained at 21 ± 1.0℃ with an alternating 12 h light-dark cycle. Rats were provided with water and food ad libitum and permitted to acclimate for at least 3 days prior to their use in experiments. The animals were allowed to adapt to the laboratory for at least 2 h before testing, and they were used only once. To reduce variation, all experiments were performed during the light phase of the cycle (10:00-17:00).
2. Experimental protocol and drug administration
The rats were randomly divided into 3 groups (21 rats per group): 1) the control group, which received only vehicle; 2) the paclitaxel group, which received paclitaxel; and 3) the EP group, which received EP after paclitaxel administration. Paclitaxel (Taxol®; Bristol-Myers-Squibb, Montreal, Canada; 6 mg/ml) was diluted with saline to a concentration of 2 mg/ml and injected intraperitoneally at a dose of 2 mg/kg on 4 alternate days (days 0, 2, 4, and 6), as previously described [5,6,21,22]. EP (98% pure; Sigma-Aldrich Co., St. Louis, MO, USA) was dissolved in normal saline to a concentration of 10 mg/ml and injected intraperitoneally at a dose of 50 mg/kg on 6 consecutive days after final paclitaxel administration. The dose of EP was chosen based on preliminary studies and previous publications [18]. In addition, previous studies have evaluated the therapeutic effect of EP administered repeatedly in several animal disease models [23,24]. Thus, following previous reports, we repeatedly administered EP on 6 consecutive days but with a modification. During EP administration, the control and paclitaxel groups were treated with the vehicle for EP (normal saline) on the same 6 consecutive days.
Behavioral tests were performed before and at 7 and 14 days after final administration of paclitaxel in order to measure behavioral responses to mechanical stimuli. DRG apoptosis was evaluated by immunohistochemical detection of activated caspase-3 14 days after final paclitaxel administration. All testing was performed by 1 examiner who was blind to the study groups.
3. Behavioral tests
Behavioral tests were performed during the same time period on each testing day (10:00 AM to 4:00 PM). Rats were positioned in individual plastic cages with wire-mesh bottoms and permitted to acclimate for 20 minutes. Baseline testing was performed before drug administration to verify that the animal responses were normal. To test for mechanical allodynia, a von Frey filament (Stoelting, Wood Dale, IL, USA) was used to stimulate the plantar surface of the left hind foot. Eight calibrated von Frey filaments (0.41, 0.70, 1.20, 2.00, 3.63, 5.50, 8.50, and 15.10 g) were sequentially applied to the plantar surface of the foot (in ascending or descending order) and gently pressed against the foot. Rapid withdrawal or flinching was interpreted as a positive response, in which case the next-lightest filament was tested; a negative response led to the application of the next-heaviest filament. Determination of the 50% withdrawal threshold was made using the up-down method, as previously described [25-27].
4. Immunohistochemistry
The histological findings in the 21 rats used in the behavioral tests were also analyzed (n = 7 rats per group), as previously described [12,18]. Briefly, under anesthesia, the rats were transcardially perfused with 4% buffered paraformaldehyde 14 days after final paclitaxel administration. The L5 DRG was collected and subsequently fixed in 4% paraformaldehyde, embedded in paraffin, and sectioned to a thickness of 4 µm. After blocking with 5% goat serum, 0.3% Triton X-100, and 1% bovine serum albumin in phosphate-buffered saline (PBS-T), double-label immunofluorescence was performed by incubating the paraffin sections overnight at 4℃ with antibodies against cleaved caspase-3 (Asp 175) (1:200; Cell Signaling Technology Inc., Danvers, MA, USA) and glial fibrillary acidic protein (GFAP) (1:200; Cell Signaling) or the N52 clone of neurofilament 200 (NF200) (1:200; Sigma-Aldrich), followed by incubation with Alexa Fluor 555 goat anti-rabbit IgG (red; 1:500; Invitrogen, Carlsbad, CA, USA) and Alexa Fluor 488 goat anti-mouse IgG (green; 1:500; Invitrogen) for 2 hours at 20℃. The immunostained DRG sections were observed using a confocal microscope. Two sections were used in each evaluation to count the number of NF200-immunoreactive (IR), GFAP-IR, and double-labeled cells at 400× magnification. The percentage of double-labeled cells in each DRG was calculated by dividing the number of double-labeled cells by the total number of NF200-IR or GFAP-IR cells within the field and multiplying the result by 100.
5. Statistical analysis
The data are presented as the means ± standard error. Behavioral data (withdrawal threshold to mechanical stimuli) were evaluated by 2-way repeated measures analysis of variance. The percentage of double-labeled cells among the DRG cells was analyzed using the Kruskal-Wallis test followed by the Mann-Whitney U-test to evaluate the between-group differences. P values less than 0.05 were considered significant.
Go to :
RESULTS
In the present study, normal weight gain was observed in all groups of animals during the testing periods (data not shown), and no animals died during paclitaxel therapy, similar to previous reports [22,28,29].
1. Effect of EP on paclitaxel-induced mechanical allodynia
The baseline threshold to evoke a withdrawal response to mechanical stimuli in all rats before drug administration was 15.10 g (maximal mechanical stimulus). However, as shown in Fig. 1, the withdrawal threshold to mechanical stimuli was reduced significantly at 7 and 14 days after paclitaxel administration (P = 0.023 for 7 days, P = 0.004 for 14 days). Post-treatment with EP did not significantly affect paclitaxel-induced allodynia (P = 0.071 for 7 days, P = 0.088 for 14 days), although it tended to slightly reduce sensitivities to mechanical stimuli after paclitaxel administration (Fig. 1).
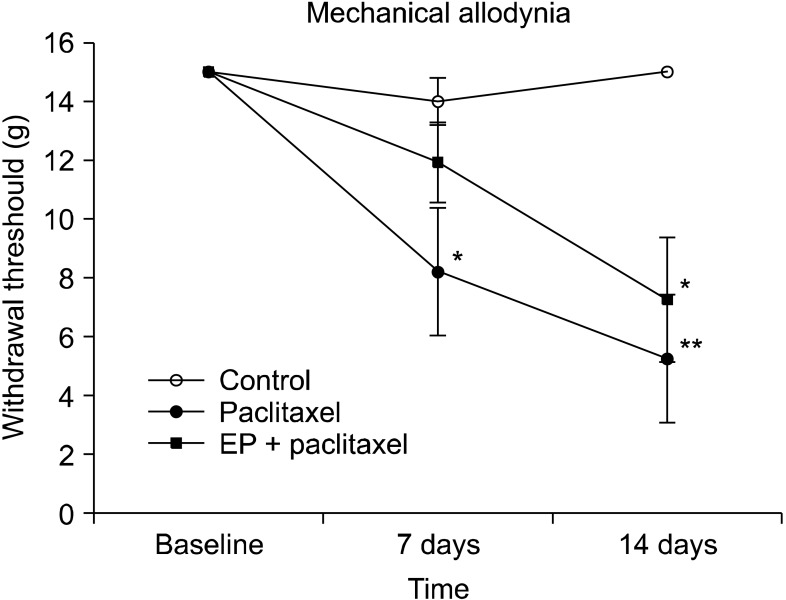 | Fig. 1The effect of ethyl pyruvate (EP) on paclitaxel-induced mechanical allodynia in rats. Groups of rats were injected i.p. with either vehicle (control) or paclitaxel (2 mg/kg on 4 alternate days). Another group of rats was treated i.p. with EP (50 mg/kg on 6 consecutive days) after final paclitaxel administration. Mechanical allodynia was tested before and at 7 and 14 days after final paclitaxel administration by application of von Frey filaments to the surface of the hind paw. The vertical bars indicate the standard error of the mean. The number of animals used for each group was 7 (*P < 0.05 and **P < 0.01 compared with the control group).
|
2. Effect of EP on NF200 and caspase-3 IRs in paclitaxel-induced neuropathy
The immunohistochemical analysis at 14 days after paclitaxel administration showed an increase in caspase-3 IR in the DRG cells, which was co-localized with NF200- positive neurons (Fig. 2A). Moreover, post-treatment with EP decreased the paclitaxel-induced caspase-3 IR, shown Fig. 2A. The percentage of NF200-positive DRG neurons co-localized with caspase-3 at 14 days after paclitaxel administration was significantly increased compared with the vehicle-treated control group (P < 0.001, Fig. 2B). The paclitaxel-induced caspase-3 IR in the DRG neurons was inhibited by post-treatment with EP (P < 0.001, Fig. 2B).
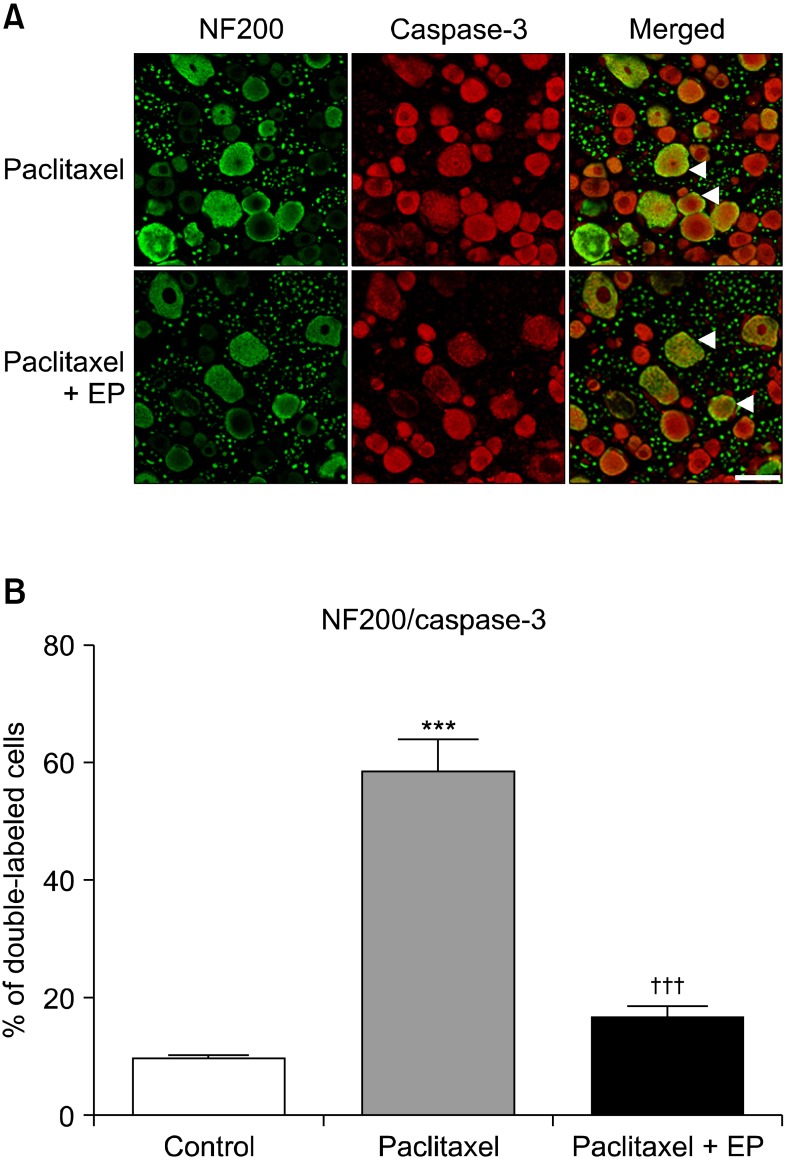 | Fig. 2The representative double immunofluorescence staining and quantitative analysis of activated caspase-3 and neurofilament 200 (NF200) in the rat dorsal root ganglion (DRG). Groups of rats were injected i.p. with either vehicle (control) or paclitaxel (2 mg/kg on 4 alternate days). Another group of rats was treated i.p. with either vehicle or ethyl pyruvate (EP, 50 mg/kg on 6 consecutive days) after final paclitaxel administration. Fourteen days after paclitaxel treatment, L5 DRG samples were immunostained with caspase-3, an indicator of apoptosis, and NF200, a marker of myelinated neurons. The co-localization of caspase-3 was visualized in a merged image. The arrow heads indicate where activated caspase-3 is co-localized with NF200. The scale bar represents 50 µm (A). The NF200 positive neurons that co-localized with caspase-3 were counted and are expressed as a percentage of the total neurons per section (***P < 0.001 compared with the control group, and †††P < 0.001 compared with paclitaxel group). Error bars represent SEM (B).
|
3. Effect of EP on GFAP and caspase-3 IRs in paclitaxel-induced neuropathy
An immunohistochemical analysis showed that paclitaxel administration did not alter the GFAP IRs in the DRG compared with the vehicle-treated control group (data not shown). Additionally, post-treatment with EP had no effect on the GFAP and caspase-3 IRs located in the DRG (Fig. 3).
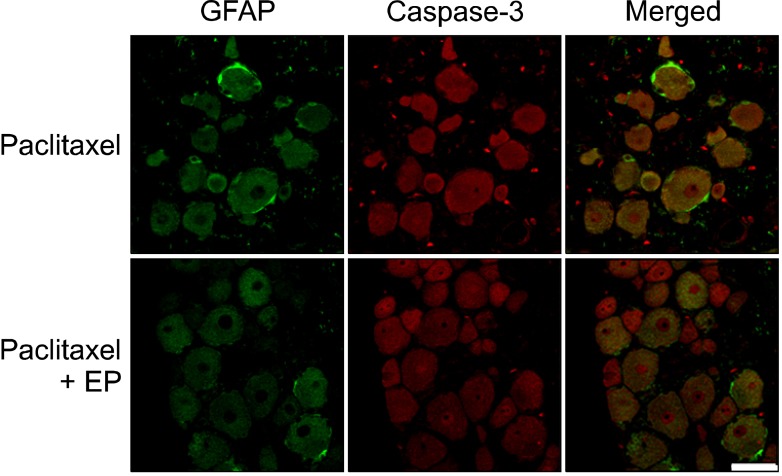 | Fig. 3The representative double immunofluorescence staining of activated caspase-3 and glial fibrillary acidic protein (GFAP) in the rat dorsal root ganglion (DRG). Groups of rats were injected i.p. with either vehicle (control) or paclitaxel (2 mg/kg on 4 alternate days). Another group of rats was treated i.p. with either vehicle or ethyl pyruvate (EP, 50 mg/kg on 6 consecutive days) after final paclitaxel administration. Fourteen days after paclitaxel treatment, L5 DRG samples were immunostained with caspase-3, an indicator of apoptosis, and GFAP, a marker of activated astrocytes. Neither paclitaxel nor post-treatment with EP affected GFAP- or caspase-3 IRs in DRG. The scale bar represents 50 µm.
|
Go to :
DISCUSSION
Systemic administration of paclitaxel can induce neuropathy, mechanical allodynia, or thermal hyperalgesia [2,3]. Intraperitoneal injection of paclitaxel at a dose of 2 mg/kg on 4 alternate days is believed to be a reliable model of peripheral neuropathy [22,29]. The pathophysiology of paclitaxel-induced neuropathic pain appears to involve changes in the primary sensory nerve, DRG, and spinal cord levels [8,9,21,28,30,31]. For example, in a previous study, myelinated A-fiber hypersensitization was observed at 1 and 2 weeks after paclitaxel treatment [21]. Another study found significant increases of atypical (swollen and vacuolated) mitochondria in both C-fibers and myelinated axons of paclitaxel-treated nerves [28]. Paclitaxel may also induce apoptosis in DRG cells and cortical neurons [7,8,32,33]. Melli et al. [7] found that paclitaxel caused early mitochondrial impairment, with loss of membrane potential and induction of autophagic vacuoles, in the primary cultures of DRG sensory neurons [7]. Another in vitro studies have shown that the neurotoxic and apoptotic effects of paclitaxel were dose- and time-dependent in DRG explants and cultured neuronal cells [8,32,33].
On the other hand, it is relatively well known that apoptosis may be evident in DRG neurons following nerve injury, which may induce allodynia [10-12]. Caspase-3 can induce the proteolytic cleavage of many key proteins, and its expression is regarded as a distinct indicator of apoptosis [34,35]. In the present study, we found increased caspase-3 IR in DRG cells, which was co-localized with NF200-positive myelinated neurons 14 days after paclitaxel administration. Moreover, post-treatment with EP decreased the paclitaxel-induced caspase-3 IR. However, EP did not affect the paclitaxel-induced mechanical allodynia significantly, although post-treatment with EP tended to slightly reduce the withdrawal threshold to mechanical stimuli after paclitaxel administration. Although we did not investigate the dose-dependent effects of EP in the present study, the possibility of an anti-allodynic effect from EP administered at doses higher than 50 mg/kg cannot be ruled out. Recently, we reported that EP has a strong anti-allodynic effect that acts through the inhibition of TNF-α expression and apoptosis in the DRG after spinal nerve ligation injury [18]. Further studies to determine the exact time and dose of EP administration are needed.
Interestingly, the present study showed that paclitaxel administration (2 mg/kg on 4 alternate days) did not alter GFAP IRs in the DRG. Contrary to our present findings, a previous study on cellular pathology in DRG after paclitaxel treatment reported that the GFAP IR levels in the DRG were increased in paclitaxel-treated rats compared with vehicle-treated rats from days 6 through 10 after final administration of paclitaxel [9]. Because these researchers injected paclitaxel intravenously into the tail vein of rats at a dose of 18 mg/kg twice (on days 0 and 3), this discrepancy may be explained in part by the differences in duration and dose of paclitaxel administration. Therefore, these findings suggest that astrocytes located in the DRG may have a time-specific or dose-specific involvement in paclitaxel-induced neuropathic pain. In addition, paclitaxel evokes the release of pro-inflammatory cytokines, including TNF-α [36,37]. Pro-inflammatory cytokines have an important role in the pathogenesis of painful peripheral neuropathies and neuropathic pain (for a review, see [38,39]). Several studies have reported the involvement of spinal glial cells and cytokines in the pathogenesis of paclitaxel-induced neuropathic pain [40,41]. Burgos and colleagues observed a strong microglial and astroglial activation and induction of pro-inflammatory cytokines in the lumbar spinal cord after paclitaxel administration [41]. In the present study, apoptosis of DRG neurons, but not mechanical allodynia, was inhibited by post-treatment with EP. Therefore, this result cannot exclude the possibility that spinal glial cells have a more important role in the development of paclitaxel-induced neuropathic pain than DRG astrocytes, although we did not examine the involvement of glial cells located in the spinal cord and the pro-inflammatory cytokine levels.
In conclusion, inhibition of apoptosis of DRG neurons by EP may not be critical in paclitaxel-induced mechanical allodynia. Further studies are needed to elucidate the exact time and dose of EP administration in the paclitaxel-induced neuropathic pain model.
Go to :
 XML Download
XML Download