

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Peripheral nerve injury may result in neuropathic pain, which is characterized by spontaneous pain, hyperalgesia, and allodynia, that is thought to be secondary to increased excitability of the neurons. Although debate is still ongoing regarding the etiology of neuropathic pain, increasing amounts of data have provided greater insight into the role of proinflammatory cytokines such as tumor necrosis factor-α (TNF-α) in the mechanism of neuropathic pain.
In the standard model of chronic constriction injury or crush injury of the sciatic nerve in rats, TNF-α has been detected at the injury site, and shows temporal up-regulation [1,2]. Elevated levels of TNF-α immunoreactivity in the dorsal root ganglia (DRG) are also observed in chronic constriction injury [3], a finding that can be partially explained by retrograde axonal transport [4].
Apoptosis has also been demonstrated in the DRG following peripheral nerve injury, and TNF-α expression is known to be associated with DRG apoptosis via the caspase signaling pathway [5]. Caspase-3 expression is one of the key factors in apoptosis, as it is responsible for the proteolytic cleavage of many key cellular proteins, and cell bodies and satellite cells expressing caspase-3 might progress to apoptosis [6]. A recent study compared crush injury distal to the DRG with injury proximal to the DRG in rats, and found that distal crush injury resulted in a greater amount of neuronal apoptosis and enhanced TNF-α expression and caspase-3 levels [7], lending additional support to a TNF-α-apoptosis-caspase signaling paradigm for peripheral neuropathic pain [8].
The aim of the present study was to evaluate TNF-α expression, activated caspase-3 in astrocyte and neuronal cells in the DRG, and DRG apoptosis after spinal nerve ligation (SNL) in rats.
Go to : 
MATERIALS AND METHODS
The experimental procedures were approved by the Animal Care Committee. Twenty-eight male Sprague-Dawley rats (weight 160-180 g) were housed three per cage in a temperature-controlled (21.0 ± 1.0℃) vivarium with water and food available ad libitum and allowed to acclimate for three days in a 12-h light/dark cycle.
1. Surgical procedure for animal model
The animals were randomly divided into two experimental groups (14 in each group). One group was subjected to spinal nerve ligation injury (SNL group), and the other group was subjected to no operation (naïve group). SNL injury was performed as previously described [1]. Briefly, a midline incision above the lumbar spine and deep dissection through the paraspinal muscles exposed the left L6 transverse process, and the process was then removed. The left L5 and L6 spinal nerves were isolated, and both nerves were ligated tightly with 6-0 black silk distal to the DRG and proximal to the formation of the sciatic nerve.
2. Behavioral tests
Animals were placed in individual plastic cages with wire mesh bottoms and allowed to acclimate for 20 min, and behavioral testing was done.
To test for mechanical allodynia, a von Frey filament was applied vertically to the plantar surface of the hind paw ipsilateral to the SNL injury. A series of eight calibrated von Frey filaments (0.41, 0.70, 1.20, 2.00, 3.63, 5.50, 8.50, and 15.10 g) was applied serially to the paw in ascending order of strength with sufficient force to cause gentle bending against the paw, and held for six seconds. A sharp withdrawal or paw flinching was considered a positive response. The mechanical stimulus producing a 50% likelihood of withdrawal was determined by using the up-down method as previously described [9].
To test for cold allodynia, acetone was applied to the plantar surface of the hind paw ipsilateral to the SNL injury, five times to each paw at intervals of 5 min. A prompt foot withdrawal response to the acetone application was interpreted as a sign of cold allodynia. The frequency of paw withdrawal was expressed as a percentage (the number of paw withdrawals divided by the total number of trials, times 100).
Baseline testing was done before surgery to verify that the animals had normal responses. All animals were tested on postsurgical days 4 and 7. All testing was done by a single examiner blinded to the experimental groups.
3. Double-labeling immunohistochemistry study
After behavioral testing, 7 rats of both SNL and naïve groups were subjected to histologic findings. The animals were anesthetized with zoletil 12.5 mg and xylazine 3 mg intraperitoneally and perfused transcardially with 4% buffered paraformaldehyde on postoperative day 7. The left L5 DRG was removed, postfixed in 4% paraformaldehyde, and subsequently embedded in paraffin.
DRG samples embedded in paraffin were sectioned (4 µm), and placed on slides. Sections were deparaffinized and rehydrated in a graded ethanol series. After blocking with 5% goat serum, 0.3% Triton X-100, and 1% BSA in PBS-T, double-label immunofluorescence was performed by incubating the paraffin sections with cleaved caspase-3 (Asp 175) antibody (1:200, #9661, Cell Signaling Technology Inc., Danvers, MA, USA) and GFAP antibody (1:200, #3670, Cell Signaling) or NeuN antibody (1:200, MAB377, Chemicon, Billerica, MA, USA), which were applied overnight at 4℃, followed by incubation with Alexa Fluor 555 goat anti-rabbit IgG (red; 1:500; Invitrogen) and Alexa Fluor 488 goat anti-mouse IgG (green; 1:500; Invitrogen, Carlsbad, CA, USA) for 2 hours at room temperature. Sections were rinsed in PBS-T and mounted with DAPI. The immunoreactions in the DRG were observed using a confocal microscope. Two slices of the DRG for each evaluation were used to determine the number of NeuN-immunoreactive (IR) or GFAP-IR cells in the DRG. Co-labeled cells with a clearly definable greenish cell border and reddish nucleus were counted as a marker of apoptotic cells at 400× magnification for each section. The number of co-labeled cells was divided by the total number of NeuN-IR or GFAP-IR cells within the field and multiplied by 100.
4. ELISA immunoassay for TNF-α
ELISA immunoassay findings were analyzed using 7 rats of both SNL and naïve groups that were not used for behavioral testing. The animals were decapitated rapidly under anesthesia on the 7th day of experiment, and the DRGs of the left L5 and L6 segments were removed and frozen in liquid nitrogen. Samples were homogenized in ice-cold RIPA buffer (R0278, Sigma, St. Louis, MO, USA) and protease inhibitor cocktail (P8340, Sigma, St. Louis, MO, USA), and centrifuged (13,000 rpm for 20 min) to obtain protein extracts. Commercially available ELISA kits were used for assessing TNF-α (R&D Systems, MN, USA, sensitivity: 5 pg/ml). The Bradford protein assay was used to measure total protein concentration in the tissue supernatant. Microplates were measured using a VICTOR3 V Multilabel Counter (1420, Perkin Elmer), and data were standardized as picograms of TNF-α to 200 µg of total protein in the supernatant.
5. Statistical analysis
All statistical analyses were performed using SPSS for Windows Version 12.0 (SPSS Inc., Chicago, IL, USA). Data are presented as mean ± SD or SEM. The differences between the groups were compared using the t-test. P values less than 0.05 were considered statistically significant.
Go to :
RESULTS
1. Behavioral tests
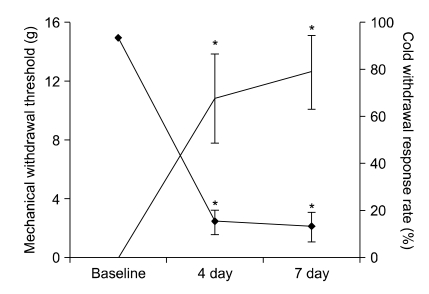
The mechanical withdrawal threshold and prompt paw withdrawal response to acetone application are shown in Fig. 1. SNL injury produced significant mechanical and cold allodynia after 4 and 7 days (P < 0.05) compared to baseline.
2. Immunohistochemistry of NeuN, GFAP and caspase-3 in the DRG
As shown in Fig. 2, the NeuN-IR cells (DRG neurons, green) and GFAP-IR cells (satellite cells, green) were observed in the DRG (A, D, G, J). The GFAP-IR satellite cells were localized around the DRG neurons. Caspase-3 immunoreactivity (red) was observed in neurons (B, E), astrocytes (H, K). After SNL, activated caspase-3 co-localized with NeuN (C, F) and GFAP (I, L). Co-localized caspase-3 expression was significantly increased in DRG astrocytes and neurons at postoperative day 7 compared with naïve DRG. (P < 0.05, Fig. 3).
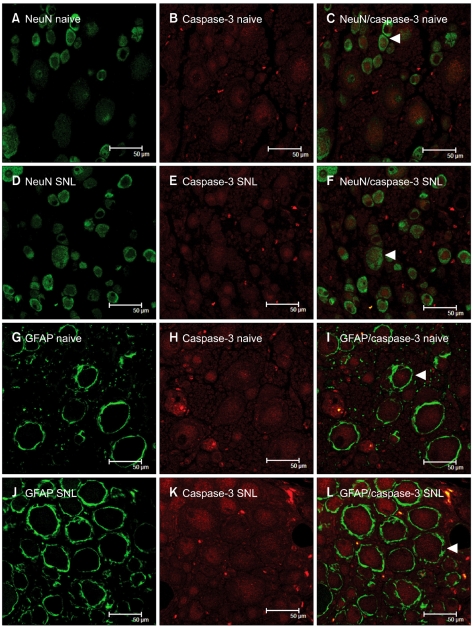 | Fig. 2Double immunofluorescence staining of activated caspase-3 and cell markers in the rat dorsal root ganglion (DRG). Tissue samples from the naïve and spinal nerve ligation (SNL) groups were immunostained with caspase-3 (B, E, H, K), an indicator of apoptosis, and cell markers, such as NeuN (A, D) and GFAP (G, J), and the co-localization of caspase-3 was visualized in a merged image (C, F, I, L). (A-F) Immunostaining for caspase-3 with NeuN in naïve and SNL DRG; (G-L) Immunostaining for caspase-3 with GFAP in naïve and SNL DRG. Note that NeuN is a marker of neurons, and GFAP is a marker of activated astrocytes. Examples of apoptotic cells are marked with white arrows.
|
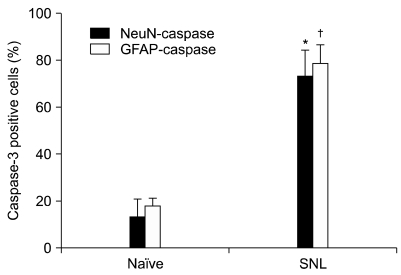 | Fig. 3Quantitative analysis of different phenotypic-specific markers in positive cells expressing caspase-3 (%) in naïve and SNL groups. The change in caspase-3 was striking in astrocytes and neurons. *,†Indicate significant difference at P < 0.05 compared with naïve group. Error bars represent SEM.
|
3. Apoptotic cells in the DRG, and ELISA immunoassay for TNF-α
Apoptotic cells were observed in the left L5 DRGs in the SNL group. The number of apoptotic cells in the SNL group (13.7 ± 1.1) was significantly higher than in the naïve group (3.5 ± 0.8, P < 0.001) on postoperative day 7. TNF-α protein levels were also increased in the DRG following SNL injury (12.7 ± 3.2 pg/100 µg, P < 0.001), when compared with naïve rats (4.1 ± 1.4 pg/100 µg).
Go to :
DISCUSSION
In the study presented here, we showed that tightly ligating the L5 and L6 spinal nerves in rats produces an approximately eightfold increase in TNF-α as measured by ELISA performed 1 week after SNL, and that caspase-3 is activated and translocated to the astrocytes and neurons in the DRG. Because the DRG was not injured directly, the apoptosis resulting from the SNL was thought to be secondary to the nerve injury. There is compelling evidence demonstrating that hyperalgesia and allodynia associated with peripheral nerve injury reflect, at least in part, changes in the excitability of primary afferent neurons [10,11]. One of the possible explanations for the pain behaviors is that proinflammatory cytokines produced at the injured site of the primary afferent neurons may activate apoptotic cascade reactions in the DRG neurons [8].
TNF-α is a pleiotropic proinflammatory cytokine that is a principal mediator of the early degenerative changes in peripheral nerve injury. TNF-α is produced during Wallerian degeneration [12] and contributes to both inflammatory [13] and neuropathic [14] hyperalgesia. DRG neurons robustly increase their expression of TNF-α after peripheral nerve injury [15]. Experimentally, various animal models of peripheral neuropathic pain have been developed; however, there are considerable differences in the magnitude and duration of response components among models [16]. In the current study, TNF-α protein levels were increased in the DRG after SNL injury. These results are consistent with previous results from other peripheral nerve injury models showing an increase in DRG expression of TNF-α.
Apoptosis plays an important role in neuropathic pain. Peripheral nerve injury produces apoptosis in the dorsal horn of the spinal cord [17-19], and DRG apoptosis is also observed after peripheral nerve injury [6,20]. Caspase-3 expression is one of the key factors in apoptosis, as it is largely responsible for the proteolytic cleavage of many key cellular proteins. The results of a study showing that inhibition of caspases reduces pain behavior suggest that caspase signaling pathways are involved in pain; upregulation of the death receptor ligand TNF-α also produces pain via caspase pathway [8]. In the histologic findings of the present study, the percentage of neurons or satellite cells co-localized with activated caspase-3, as well as the number of apoptotic cells, were significantly higher in the SNL group than in the naïve rats. Neurons and astrocytes expressing caspase-3 might progress to apoptosis.
In the current study, we demonstrated that pain behaviors following spinal nerve injury were related to increased levels of TNF-α and apoptosis in the DRG. However, this study has several limitations. First, because of the short period of observation following spinal nerve injury, it is not clear whether increases in TNF-α and apoptosis affect the maintenance of the pain behaviors after 7 days. Previous studies showed that the time of peak TNF-α expression and of apoptosis is about 7 days after nerve injury [6,21]. Therefore, we decided to perform the ELISA immunoassay for TNF-α and the histologic studies at postoperative day 7. The second limitation of our study is that we did not investigate pain behaviors in the sham operation group. A previous study, however, demonstrated that operation per se did not induce any pain-related behaviors [22]. Lastly, we did not compare apoptosis in the spinal cord between the two groups. Apoptosis has been found in the spinal cord after spinal nerve injury, and is associated with pain-related behavior [18]. Future studies must investigate the changes of the spinal cord.
In conclusion, spinal nerve ligation injury produces significant mechanical and cold allodynia, as well as TNF-α elevation and apoptosis in the DRG. Considering that inhibition of TNF-α by selective antibody therapy attenuates mechanical allodynia in animals with spinal nerve ligation injury, further work is warranted to investigate the effect of such approaches in different study designs.
Go to :
 XML Download
XML Download