

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Pregabalin is an anticonvulsant analgesic agent that binds to the alpha-2-delta subunit of voltage sensitive Ca2+ channels in the presynaptic area [1-5]. As for the site of action, intrathecal pregabalin reduces presynaptic neurotransmitter release without altering postsynaptic receptors [3]. However, gabapentin and pregabalin reduced the K+-evoked glutamate release without affecting basal glutamate levels [6]. And the actions of pregabalin may involve both extracellular and intracellular drug target sites and modulation of a variety of neuronal conductances, by direct interactions and through intracellular signaling involving protein kinase A [7].
Opioid receptor agonists open K channels in neurons through the activation of G-proteins. The analgesic effect of morphine can be prevented by the inhibition of KATP channels and GIRK channels [8]. These 2 types of K channel activations lead to cell membrane hyperpolarization and are involved in opioid induced antinociception [9]. In addition, morphine activates presynaptic opioid G-protein-coupled mu-opioid receptors or inhibits calcium currents, thereby reducing neurotransmitter release [10].
Gabapentin or pregabalin reduce excitatory neurotransmitter release by binding to the alpha-2-delta subunit of voltage sensitive Ca2+ channel even though they are not Ca2+ blockers [2]. Pregabalin reduces the excitatory properties of cultured DRG neurons by modulating voltage-activated Ca2+ and K+ channels [7]. On the other hand, the anti-allodynic effect of gabapentin was produced by the activation of Ca2+ activated and ATP-sensitive K+ channels [11]. These results led us to postulate that an association between KATP channels and the analgesic effect of pregabalin may exist. In addition, if KATP channels are associated with pregabalin action, the evaluation of KATP channel expression in the spinal cord after intrathecal injection of pregabalin will yield important information on the mechanism of the analgesic effect of pregabalin.
The aim of this study was to evaluate whether the analgesic action of intrathecal pregabalin is associated with KATP channels in the rat formalin test. In addition, the effect of pregabalin on KATP channel expression in the spinal cord dorsal horn was also evaluated.
Go to : 
MATERIALS AND METHODS
This study was approved by the Institutional Animal Care Committee. Adult male Sprague-Dawley rats weighing 250-300 g were housed in individual cages on a 12 h night/day cycle at a constant temperature of 20-22℃. Food and water were available ad libitum.
1. Catheter implantation
For intrathecal (IT) drug administration, rats were chronically implanted with catheters as previously described [12]. Briefly, under enflurane anesthesia, the rat were placed in a stereotaxic head holder. The occipital muscles were separated and retracted caudal to expose the cisternal membrane at the base of the skull. A polyethylene tube (PE-10 catheter) was passed caudally from the cisterna magna to the level of the lumbar enlargement. After bleeder ligation, the tissues were sutured layer by layer. Rats with evidence of neuromuscular dysfunction were promptly sacrificed. After 5-7 days of the catheterization, rats without neurological deficit and infection were considered for data analysis.
2. Neurobehavioral test
1) Drugs: Forty five rats were divided into 3 groups according to the IT administered drug. In the control (CON) group (N = 5), normal saline 10 µl was administered as a placebo and flushed with normal saline 10 µl. In the pregabalin (PGB) group (N = 20), pregabalin 0.3, 1, 3 or 10 µg dissolved in normal saline 10 µl were administered and flushed with normal saline 10 µl. In the glibenclamide (GBC) group (N = 20), IT injection of glibenclamide 100 µg dissolved in dimethyl sulfoxide (DMSO) 5 µl was followed by pregabalin 0.3, 1, 3, or 10 µg dissolved in normal saline 5 µl and flushed with normal saline 10 µl.
2) Formalin test: After 10 min of IT drug administration, 5% formalin solution 50 µl was injected intradermally to the dorsal surface of the left hind paw. Nociceptive behavior was determined by the number of flinches of the left hind paw during 60 min. Flinching per minute was averaged over 5 min. The initial 10 min was classified as phase 1 and the next 11-60 min as phase 2. Maximum possible effect (%MPE) was calculated as follows:
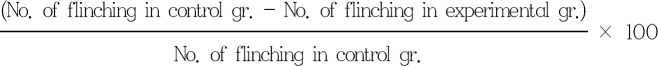
3. Immunohistochemistry
1) Perfusion fixation: Immunohistochemitry was done separately with a different set of 13 rats. In the naïve group (N = 4), rats were deeply anesthetized 1 hr after administration of intrathecal normal saline 20 µl without formalin test. In both the normal saline group (N = 4) and pregabalin group (N = 5), the formalin test was done 10 min after IT administration of normal saline 20 µl or pregabalin 10 µg dissolved in normal saline 10 µl and flushed with normal saline 10 µl. Rats were deeply anesthetized and fixed by vascular perfusion with paraformaldehyde 1 hr after drug injection. The spinal cord was dissected and placed in 4% paraformaldehyde for an additional 2 h. Then the spinal cord was transferred to a 30% sucrose solution and incubated for 1 day at 4℃ for cytoprotection. A microtome (Leica Microsystems, Wetzlar, Germany) was used to cut 40 µm floating sections stored in phosphate buffer. Sections were rinsed 3 times in PBS then blocked in 4% goat serum and 0.3% Triton X-100. Antigen was retrieved by the heat induced epitope retrieval method after rehydration. Inactivation of endogenous peroxidase was conducted with 0.3% H2O2 incubated for 5 min at room temperature. After 3 more PBS rinses, sections were incubated for 1 hour at room temperature and over-night with the primary antibodies (1:100 Kir 6.1 (H-80)) diluted in blocking buffer at 4℃. Sections were rinsed again with PBS and incubated for 20 min at room temperature with secondary antibodies (anti-rabbit IgG, Envision kit, DAKO: Santa Cruz Biotechnology, Inc. Santa Cruz, USA). The slices were counterstained with hematoxylin and dried before they were mounted with media.
2) Imaging: Images were taken using a SP1 confocal microscope and image capturing software (Leica Microsystems, Wetzlar, Germany). Quantifications were done using the Metamorph Imaging System software (Universal Imaging Corporation). Average pixel intensities within a region that approximated the laminae I-II of the ipsilateral spinal cord dorsal horn were calculated and normalized to a region of a comparable area in the contralateral dorsal horn. Threshold intensities were determined for each image above with most of the background noise suppressed. A rectangular box of constant size was placed over the entire width of the dorsal horn and the area of pixels within the box with an intensity value above the threshold was determined for both the injured and uninjured side. The results are expressed as the percentage change in staining ipsilateral to the injury compared to the uninjured side. The calculated values of percent change for each section were then averaged to yield a value for each animal, and a mean and standard error were determined per group. The experimenter was blinded to the treatment group during image collection and quantification.
4. Statistical analysis
All of the data were presented as the mean ± SEM. The dose-response data were expressed as %MPE in each phase. The dose response data was analyzed using one-way ANOVA with Scheffe for post hoc. The ED50 values were analyzed using dose-response analysis and the difference in the ED50 between the glibenclamide group and pregabalin group was analyzed with the student's t-test. One-way ANOVA was used for comparison of the KATP channel expression in the spinal cord among groups. If the P value was less than 0.05, the results were considered statistically significant.
Go to :
RESULTS
1. Neurobehavioral test
1) After IT injection of pregabalin and/or glibenclamide, all animals showed no change in motor function such as the placing/stepping reflex and righting reflex tests.
2) A subcutaneous injection of formalin resulted in a highly reliable biphasic display of flinching of the injected paw (Fig. 1A). IT injection of pregabalin decreased the sum of flinches in a dose dependent manner in phase 2 (P < 0.05), but not in phase 1. Additionally, a fixed dose of glibenclamide reversed the analgesic effects of pregabalin (Fig. 1B).

3) The %MPE curve and ED50 of pregabalin shifted to the right in a log dose response curve compared to that of the glibenclamide pretreated rats (P < 0.01)(Fig. 2).

2. Immunohistochemistry
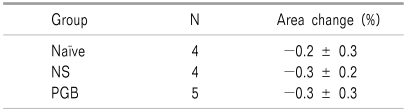
The results of the immunohistochemistry on the spinal cord dorsal horn showed no difference among the naïve group (no formalin test), normal saline group (IT normal saline 20 µl 10 min before formalin injection) and pregabalin group (IT pregabalin 10 µg administered 10 min before formalin injection) in pixel intensities for KATP channels (Fig. 3, Table 1).
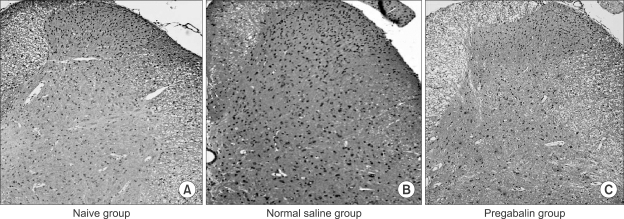 | Fig. 3Results of Immunohistochemistry of KATP channel on the spinal cord dorsal horn (magnification of 10×). The results of immunohistochemistry on the spinal cord dorsal horn shows no difference among the naïve group (A, no formalin test), normal saline group (B, IT normal saline 10 min before formalin injection) and pregabalin group (C, IT pregabalin 10 µg administered 10 min before formalin injection) in pixel intensities of KATP channel.
|
Go to :
DISCUSSION
In the present study, IT pregabalin in a dose-dependent manner decreased the number of flinching in phase 2 of the formalin test and the log dose response curve IT pregabalin was shifted to the right when glibenclamide was added. There was no change in KATP channel expression in the spinal cord dorsal horn after IT pregabailn treatment in the formalin induced pain model.
In this study, the antinociceptive effect of IT pregabalin was inhibited by glibenclamide, which suggests KATP channel activation as the antinociceptive mechanism of IT pregabalin. This is in agreement with a previous study, which reported that the antiallodynic effect of gabapentin in a spinal nerve ligation model was associated with ATP-sensitive K channels [11]. The KATP channel is known to exert its antinociceptive effect by regulating the excitability of peripheral neurons and inhibiting neurotransmitters such as substance P in the spinal cord [13-15]. This may explain the preemptive analgesic effect of pregabalin in this study.
A recent review demonstrated that although perioperative pregabalin administration did not reduce pain intensity, opioid consumption was significantly decreased during the first 24 h after surgery [16]. Gabapentin and pregabalin modulate the pain pathway by targeting the presynaptic Ca2+ channel alpha-2-delta subunit, inhibiting K+-induced Ca2+ influx and reducing the release of presynaptic glutamate and aspartate [17]. It has also been reported that the KATP channel opening effect of pregabalin is one of the mechanism for reducing neuronal excitability [18]. In Chung's model, an IT KATP inhibitor such as glibenclamide and a Ca2+ activated K+ channel inhibitor such as charybdotoxin and apamin reduced the antiallodynic effect of gabapentin [11]. A subsequent study showed that this effect was associated with the NO-cyclic GMP-PKG-K+ channel pathway [19]. Nitric oxide (NO) is synthesized during the pain process and it activates guanylate cyclase, which increases cGMP and opens the K+ channel [20]. Administration of a (Rp)-analogue of c-AMP prevents an increase in K+ current induced by pregabalin, which means that the intracellular response of pregabalin is associated with protein kinase A [7]. Another potassium channel suspected to be associated with pain modulation is the big potassium (BK) channel, which is a large conductance Ca2+ and voltage activated K+ channel. Non-inactivating BK channels with low activity can be activated by protein kinase A [21]. BK channel also have an important role in pain modulation and found in the dorsal root ganglion [22]. Therefore, BK channels can be a possible target of pregabalin but further research is needed to clarify the mechanism.
Opening of the KATP channels results in an efflux of K+ and occurs by membrane hyperpolarization and reduction in excitability. Therefore, the up-regulation of KATP channels have a protective effect when ischemia or reperfusion cause tissue injury by Ca2+ overload and can be a promising target for pain control. However, our results showed that there is no change in channel expression for KATP in the spinal cord under the rat formalin test. This result is different from Yin's report that Kir 6.2 mRNA expression was significantly increased after sciatic nerve injury [23]. This may have resulted from the difference between the inflammatory pain model and the neuropathic pain model in terms of KATP channel involvement in the pain modulation process.
Our study has the following limitations. Animals for the immunohisochemistry analyses were sacrificed 1 hr after the formalin test and there was no difference in gene expression in our study. As with other studies the change in gene expression can be delayed up to 24 hours, and the result such that the expression of the KATP channels did not change at 1 hour after the formalin test.
The antinociceptive effect of pregabalin in the rat formalin test was indirectly associated with the activation of the KATP channels. However, pregabalin did not induce KATP channel expression in the spinal cord dorsal horn 1 hr after the formalin test.
Go to :
 XML Download
XML Download