

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Transient receptor potential vanilloid 1 (TRPV1) is a non-selective cation channel involved in inducing the symptoms of pain in response to heat, protons, and capsaicin (CAPS) [1]. It is expressed in the dorsal root ganglia (DRGs), the cell bodies of the nociceptors extending to Aδ- and C-fibers [2]. It is well established that the activation of this channel is responsible for mechanical allodynia [3] and thermal hyperalgesia [4]. Several inflammatory mediators modulate TRPV1 indirectly through binding to their receptors and formation of second messengers that activate TRPV1, leading to its phosphorylation [5,6]. This stimulation increases the probability of activating the open state of this channel or its translocation to the plasma membrane [5,6].
Despite the fact that there are several analgesics in the market, many of them have side effects [7]. Accordingly, there is an increasing need for the development of new and safe drugs. Ononis spinosa is a spiny herb with trifoliate serrate leaflets, non-sticky hanging stems, and pink to violet flowers [8]. It is common in Asia, Europe, and Africa [9]. This plant belongs to Fabaceae family and is used for the treatment of inflammatory diseases, kidney stones [10], wound healing [11], and toothache [12]. Also, the anti-nociceptive effects of O. spinosa were reported previously using thermal and chemical models of pain [10,13].
Mechanical allodynia is defined as a painful sensation that results from innocuous stimuli such as touch [14]. It can be produced by the intraplantar (ipl) injection of CAPS (the pungent ingredient in chili pepper and the agonist of TRPV1 channel) into the hind paw of animal [2,3]. The nociceptive flexion reflex is quantified by applying an increasing mechanical force into the rat’s hind paw [15]. Since previous studies have not examined the effect of O. spinosa in any model of mechanical allodynia, the present study was designed to evaluate this effect in rats. In addition, the mechanism of action of O. spinosa was investigated using a TRPV1 antagonist and an antagonist for β2 adrenoreceptor. Molecular modelling was, also, used to predict the binding affinities of the active constituents in O. spinosa extract to TRPV1 receptor.
Go to : 
MATERIALS AND METHODS
1. Drugs
Butoxamine hydrochloride, a selective β2 adrenoreceptor antagonist, was purchased from Sigma-Aldrich, St. Louis. Diclofenac sodium, a non-steroidal anti-inflammatory drug (NSAID), was brought from Novartis, El-Salvador. CAPS, a TRPV1 agonist, and BCTC [4-[3-Chloro-2-pyridinyl]-N-[4-[1,1-dimethylethyl] phenyl]-1-piperazinecarboxamide], a TRPV1 antagonist, were brought from Tocris Bioscience, Bristol, United Kingdom. Methanol and ethanol were purchased from Scharlau, Spain. CAPS was dissolved in virgin olive oil, Al-Jazzazi Mill, Salt, Jordan, while BCTC was dissolved in absolute ethanol. Diclofenac sodium (2.5%), butoxamine, and O. spinosa extract were dissolved in sterile normal saline. All drugs were freshly prepared and injected (ipl) in 100 µL total volume.
2. Plant collection and extraction
O. spinosa was collected from Subaihi, Al-Balqa, Jordan in July 2017. The collected plant was identified by Prof. Barakat Abu-Irmaileh, Faculty of Agriculture at the University of Jordan. A voucher specimen was kept at the Graduate Studies Laboratory in Al-Ahliyya Amman University. Methanolic extract of O. spinosa leaves was obtained by maceration of groud leaves in 96% methanol. After filtration, the methanol was evaporated using a rotary evaporator at a temperature not exceeding 45°C, then was stored at –20°C. The extract was freshly prepared before use and was dissolved in sterile normal saline.
3. Experimental animals
All procedures were performed according to the Guide for the Care and Use of Laboratory Animals published by the National Institutes of Health and comply with the guidelines of the International Association for the Study of Pain. Ethical approval (No. AAU-2/4/2018) for conducting the work was obtained from Al-Ahliyya Amman University. Male Wistar rats (250-300 g) were brought from the animal house at Al-Ahliyya Amman University, Jordan, and were kept at 23 ± 2°C with an alternating 12 hour light-dark cycle. Food and water were provided ad libitum. The animals were allowed to adapt to the laboratory conditions for at least 2 hours before test.
4. Treatment of animals
The animals were divided into 8 groups (with 8 rats in each group). Mechanical allodynia was induced by the ipl injection of 40 µg CAPS into the left hind paw of the animals. Group I received ipl injection of vehicle only (control group). Group II received ipl injection of CAPS. Group III received ipl injection of 100 µg plant extract into the left hind paw 20 minutes before the ipl injection of CAPS. Group IV, the positive control group, received 2.5% diclofenac sodium 20 minutes prior to CAPS injection. The dose and time of injecting diclofenac sodium was chosen pursuant to the study of Nozadze et al. [16]. Groups V and VI received 300 µg BCTC and 150 µg butoxamine 30 minutes prior to the injection of plant extract, respectively followed by CAPS injection. Groups VII and VIII received the antagonists BCTC and butoxamine 30 minutes before CAPS injection, respectively. All drugs, vehicle, and plant extract were injected in a volume of 100 µL/paw into the left paw of the rats.
5. Measurement of CAPS-induced mechanical allodynia
The experimental rats were allowed to acclimatize for 1 hour in wire mesh-floor cages covered with perforated lids. Baseline withdrawal thresholds were assessed in the first day to ensure that the animals have equivalent baseline readings before starting the test. Paw withdrawal threshold (PWT) of the ipsilateral (left hind paw) and the contralateral (right hind paw) hind paws were measured using handmade von Frey filament (pursuant to Ferrier et al. [17]) 30, 90, and 150 minutes after CAPS injection. Briefly, a thin filament was used to press against the plantar surface of the animal’s hind paw. The filament was pressed once against the hind paw of the animal at each measured time point. A sharply withdrawn paw was regarded as a positive response. A cut-off mass unit of 80 g was used in this experiment according to Ferrier et al. [17]. Also, the behavior of animals was observed and recorded.
6. Molecular docking
The X-ray crystallographic structure of TRPV1 (PDB ID: 5IS0) was selected from Protein Data Bank (https://www.rcsb.org/) [18]. The crystal structure is co-crystallised with the TRPV1 antagonist, capsazepine [19]. The molecular docking tool, AutoDock 4.2 (The Scripps Research Institute, La Jolla, CA) was used to study the intermolecular interactions and binding energies of the proposed compounds found in O. spinosa leaf extract.
7. Ligand preparation
The two dimensional (2D) chemical structures of the major compounds found in O. spinosa leaf extract were generated by ACD/ChemSketch software [20]. All compounds were saved as mol files by ChemSketch software then converted to pdb files after minimization utilizing BIOVIA DS Visualizer [21]. The pdb files were uploaded in AutoDockTools and the gasteiger charges were added to each compound. These files were converted to AutoDock pdbq format, indicating all possible rotatable bonds in the structure.
8. Protein preparation
The X-ray crystallographic structure of TRPV1 (PDB ID: 5IS0) was downloaded from the Protein Data Bank. According to the co-crystallised ligand (capsazepine), the intermolecular interactions involved chains B and C, while chains D and E were removed from the crystal structure prior to docking. Polar hydrogens as well as Kollman charges were added on the amino acids. The charged protein was solvated using addsol utility in AutoDockTools.
9. Docking simulation
In Abbas et al. [22], the compounds in the methanolic leaf extract of O. spinosa were identified using liquid chromatography–mass spectrometry (LC-MS) analysis. These compounds were used to determine the docking simulation in this study. The grid parameter file was prepared using AutoDockTools to limit the search area, and a set of grid maps was created, using AutoGrid 4 (The Scripps Research Institute). A grid box was then utilized to select the area of protein structure to be mapped. The box size was set to 22.5, 22.5, and 22.5 Å (x, y, and z, respectively). Docking simulations were carried out using the Lamarckian genetic algorithm parameters and 100 runs per simulation. After the completion of docking simulation, the output information of the docked coordinates and free energy of binding in the docking log file (dlg) were obtained from AutoDock. The 2D intermolecular interactions between TRPV1 (PDB ID: 5IS0) and campesterol, stigmasterol ononin and capsazepine were prepared by PoseView [23,24].
10. Statistical analysis
The normality test was conducted for all groups using Shapiro–Wilk test. The statistical significance of differences between groups was assessed by one-way analysis of variance, followed by Tukey’s post-hoc test using GraphPad Prism version 7 (GraphPad Software, San Diego, CA). P < 0.05 was considered significant. Data were presented as means ± standard error of the mean.
Go to :
RESULTS
1. Effect of different treatments on CAPS-induced mechanical allodynia
The injection of CAPS into the left hind paw of rats induced mechanical allodynia at all-time points (measured 30, 90, or 150 min after CAPS injection) and decreased the PWT significantly compared to control group that received the vehicle (Fig. 1). Pre-injecting 100 µg O. spinosa methanolic extract into the same hind paw was effective in alleviating CAPS-induced mechanical allodynia at all-time points (Fig. 1). O. spinosa increased the PWT by 5.35-fold compared to the 3.59-fold increase caused by diclofenac sodium. In addition, the anti-allodynic effect of O. spinosa in CAPS-induced mechanical allodynia decreased when the animals received 150 µg butoxamine or 300 µg BCTC prior to the ipl injection of the plant extract. This effect was exhibited at all time-points, 30, 90, and 150 minutes after CAPS injection (Fig. 1). Besides, the animals that received CAPS injection displayed guarding behavior for their paw compared to other groups. When PWT was measured in the right hind paw (contralateral to CAPS injection) in all groups, no effect was found at any time point (Fig. 2).
 | Fig. 1Mechanical paw withdrawal threshold (PWT) of the left hind paw (ipsilateral to the injections) measured 30 (A), 90 (B) and 150 minutes (C) after intraplantar injection of the vehicle (control) or capsaicin (CAPS), P < 0.05. Data are presented as means ± standard error of the mean. BCTC: [4-[3-Chloro-2-pyridinyl]-N-[4-[1,1-dimethylethyl] phenyl]-1-piperazinecarboxamide]. aSignificant compared to control. bSignificant compared to CAPS-treated group. cSignificant compared to O. spinosa/CAPS-treated group.
|
 | Fig. 2Mechanical paw withdrawal threshold (PWT) of the right hind paw (contralateral to the injections) measured 30 (A), 90 (B) and 150 minutes (C) after intraplantar injection of capsaicin (CAPS) or vehicle. Data are presented as means ± standard error of the mean. BCTC: [4-[3-Chloro-2-pyridinyl]-N-[4-[1,1-dimethylethyl] phenyl]-1-piperazinecarboxamide].
|
2. Molecular docking
All the proposed compounds in this study were successfully docked against TRPV1 (Table 1). The compounds that had the lowest binding energies are listed in Table 2 and Fig. 3. These compounds include campesterol, stigmasterol, and ononin with –11.73, –11.02, and –8.91 kcal/mol binding energies, respectively. The results showed a conserved intermolecular interaction in these compounds, as well as the TRPV1 antagonist, capsazepine. The amino acids Leu515 and Leu553 were found to participate in the hydrophobic interaction with campesterol, stigmasterol, ononin, and capsazepine. Importantly Arg557 was shown to participate in the hydrogen bond interactions with previously mentioned compounds, except with capsazepine. Interestingly, the free energy of binding for the docked compounds is greatly affected by the number of residues that participate in hydrophobic interactions.
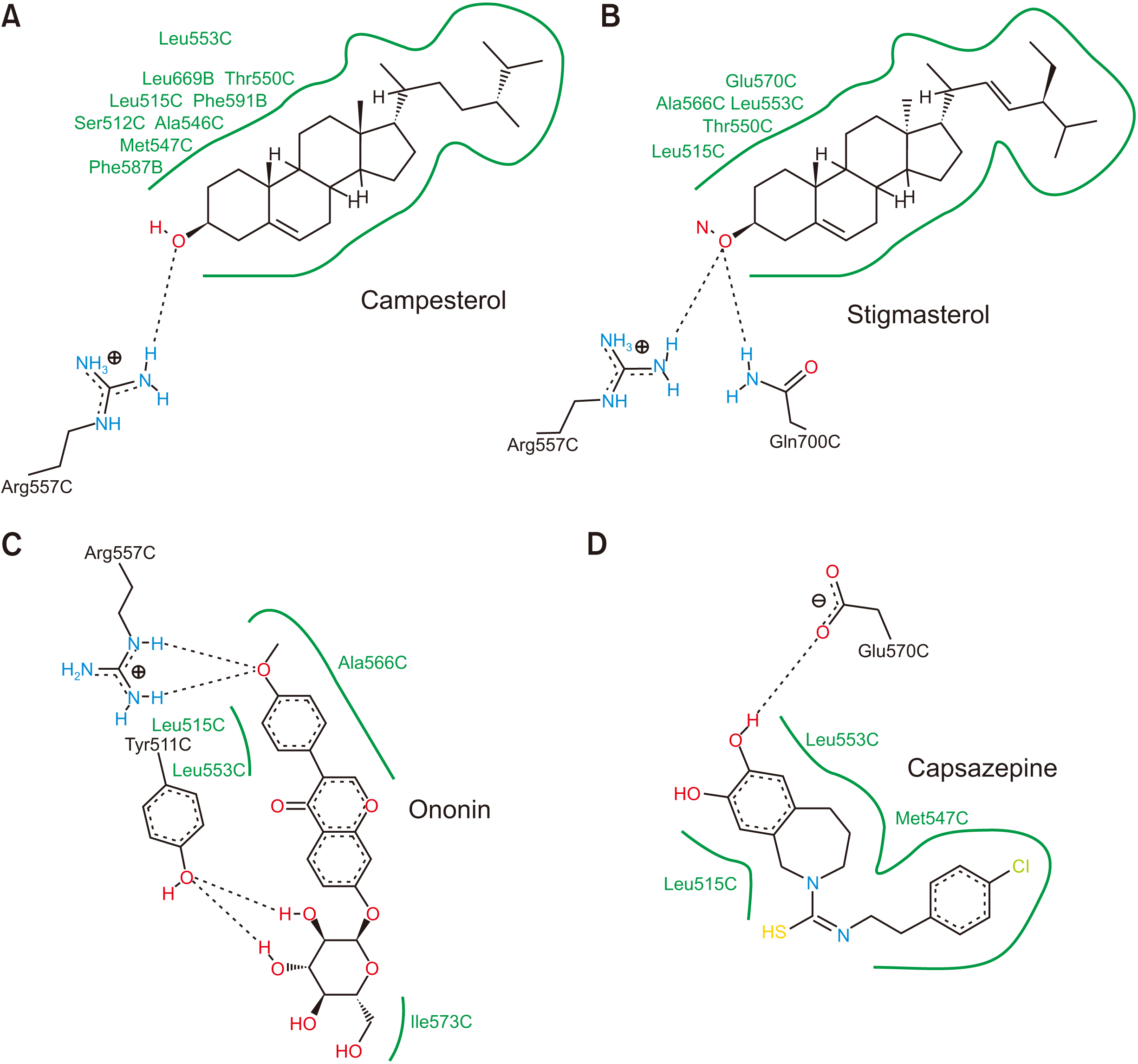 | Fig. 3Representations of the two dimensional intermolecular interactions between transient receptor potential vanilloid 1 (PDB ID: 5IS0) and (A) campesterol, (B) stigmasterol, (C) ononin, and (D) capsazepine.
|
Table 1
Free energy of binding among compounds found in Ononis spinosa leaf extract and the co-crystallised ligand (capsazepine)
![]()
Table 2
Top compounds in Ononis spinosa leaf extract having the lowest free energy of binding and with co-crystallised ligand (capsazepine)
![]()
Go to :
DISCUSSION
It is well established that the activation of TRPV1 channel is responsible for mechanical allodynia [2]. In this study, the anti-allodynic activity of O. spinosa leaf extract was assessed in CAPS-induced mechanical allodynia in male Wistar rats. To the best of our knowledge, this is the first research reporting the effect of O. spinosa in alleviating CAPS-induced mechanical allodynia.
Ipsilateral injection of O. spinosa methanolic extract into the left hind paw, 20 minutes prior to CAPS injection, increased PWT in the animals by 5.35-fold. Of note, the anti-allodynic activity of O. spinosa was stronger than the effect of the NSAID diclofenac sodium that was used as a positive control. In this study, the anti-allodynic action of O. spinosa in CAPS-induced mechanical allodynia model was attenuated using BCTC, a TRPV1 antagonist, indicating that its mechanism of action involves TRPV1 modulation. Furthermore, the decrease in PWT observed in the group that received BCTC/O. spinosa/CAPS compared to the BCTC/CAPS-treated group suggests that the plant extract can be an antagonist that binds allosterically to TRPV1 channel or an inverse agonist for TRPV1. Another possibility is that the effect of the plant extract is implicated in influencing the availability of the channel on the plasma membrane, and this effect can be direct or indirect. Further research is needed to clarify these points. Additionally, the ipsilateral pretreatment with the β2 adrenoreceptor antagonist, butoxamine, decreased the effect of O. spinosa on CAPS-induced mechanical allodynia, suggesting that the mechanism involves β2 adrenoreceptor. No effect was found on the contralateral hind paw meaning that the anti-allodynic effect of O. spinosa extract was local and not systemic.
Interestingly, previous studies have investigated the interaction between β2 adrenoreceptors and TRPV1 [25,26]. It is well established that β-adrenoreceptors are coupled to G stimulatory protein that enhances adenylate cyclase activity and intracellular cyclic adenosine monophosphate (cAMP), leading to the activation of protein kinase A, PKA (a cAMP-dependent protein kinase) [27]. Therefore, it is expected that β2 adrenoreceptor blockers will deactivate PKA, thereby reducing TRPV1 activity and its availability in the plasma membrane, finally elevating PWT in the CAPS-induced mechanical allodynia model. This may explain the findings in the present study in which a β2 blocker (butoxamine) decreased the anti-allodynic action of O. spinosa in CAPS-induced mechanical allodynia.
Notably, there are several receptors in the noradrenergic system whereby the effect of the noradrenergic system depends on the subtype of the receptors involved. Noradrenaline reduced the amplitudes of the inward CAPS currents in DRG neurons by either yohimbine (an antagonist for α2 adrenergic receptor) or propranolol (a β adrenoreceptor antagonist) [25]. Previous reports showed the involvement of PKA and protein kinase C (PKC) in the inhibitory effect of α2 adrenergic receptors on TRPV1 [25]. However, it was found that α2 adrenergic receptors inhibited the function of TRPV1 in DRG neurons in a mechanism that involves calmodulin-dependent kinase II [28].
Earlier reports showed the involvement of β2 adrenoreceptors in pain modulation, generally. In more detail, inhibition of catechol-O-methyltransferase, the enzyme that degrades catecholamines, increased pain sensitivity through activation of both β2- and β3-adrenergic receptors [29]. Also, epinephrine caused mechanical hyperalgesia mediated by β adrenoreceptor and an in vitro sensitization of rat nociceptors [15]. Furthermore, the interaction between β2 adrenoreceptors and TRPV1 was reported in DRG neurons in a model of visceral hypersensitivity [30]. Catecholamines produced a significant reduction in TRPV1 desensitization in cultured DRG neurons, and this effect was mediated by α1-, α2-, and β2-adrenoreceptors [26]. According to Coutaux et al. [31], epinephrine and norepinephrine do not normally activate nociceptors, but sensitize them under specific conditions, generating hyperalgesia. This sensitizing effect is probably mediated by protein kinases, mainly PKA and, to a lesser extent, PKC [15,31]. Therefore, the ability of butoxamine, in the present study, to abolish the anti-nociceptive effect of O. spinosa is most probably through kinases rather than a direct effect on TRPV1 channel.
Importantly, the anti-nociceptive effect of O. spinosa was investigated previously in thermal models including tail flick and hot plate tests, as well as chemical pain models such as formalin and writhing tests [10,13]. However, it should be kept in mind that there are differences between the mechanisms of mechanical allodynia and thermal hyperalgesia at different levels [32]. In more detail, the neurons that transmit the signals from mechanoreceptors synapse at the superficial lamina of the spinal cord, while the signals from thermoreceptors synapse at the superficial and deep laminae of the spinal cord [32].
The results of the behavioral assay used in this study showed the efficacy of O. spinosa in blocking TRPV1, and the substance or group of substances responsible for that effect were specified. The molecular docking tool (AutoDock) selects the compounds that have the potential to bind proteins and is therefore a forward-looking plan towards improving and developing active small molecules [33]. Molecular modelling aids in the drug discovery process and reduces experimental cost and time [33].
O. spinosa belongs to the family Fabaceae that is rich in isoflavonoids (phytoestrogens) [34]. In fact, the crude extract includes different constituents that may function on different receptors and exert synergistic, additive, or antagonistic effects. In a previous study conducted in our laboratory, the ingredients of the methanolic leaf extract of O. spinosa were determined using LC-MS analysis [22]. Among these ingredients, the present study shows that several compounds were found to interact with TRPV1 based on molecular docking. Campesterol, stigmasterol, and ononin exhibited the strongest effect in this interaction, whereby ononin alone comprises almost a fifth of the compounds (20.5%) in O. spinosa extract [22]. According to Peres et al. [35], the phytosterols campesterol and stigmasterol had analgesic effects in a writhing test. Of note, acetic acid injection in the writhing test causes abdominal constriction and trunk twisting [36]. This test involves the activation of acid-sensitive ion channels and TRPV1 channel in sensory primary fibers [37]. In another study, it was found that stigmasterol markedly attenuated the mechanical allodynia caused by surgical incision, partial sciatic nerve ligation, and complete Freund’s adjuvant [38]. All these studies justify the effect of O. spinosa on CAPS-induced mechanical allodynia.
The present study sheds the light on the importance of O. spinosa extract in modulating TRPV1 action in mechanical allodynia. However, it raises many questions that need further investigation: 1. What are the extract’s constituents that block β2 adrenoreceptor? 2. What are the effects of campesterol, stigmasterol, and ononin on TRPV1 in electrophysiological studies? 3. What are the functions of these compounds in binding studies with TRPV1, are they antagonists or inverse agonists? Future research may answer these questions.
In conclusion, O. spinosa alleviated CAPS-induced mechanical allodynia through 2 mechanisms: the direct modulation of TRPV1 and the involvement of β2 adrenoreceptor signaling. Further clinical studies are needed to confirm the effectiveness and safety of O. spinosa as an analgesic in human.
Go to :
 XML Download
XML Download