

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Lumbar disc herniation (LDH) is a common cause of radicular pain, typically manifested as hyperalgesia, allodynia, and sometimes spontaneous pain [1]. It compromises patients’ life quality and work capability, and thus becomes a socioeconomic problem that needs immediate attention [1]. Currently, the mechanism of radicular pain is still obscure.
Toll-like receptor 4 (TLR4), which is involved in innate immune response, is a trans-membrane protein with both extracellular leucine-rich repeat domains and a cytoplasmic signaling domain [2,3]. When binding with an endogenous or exogenous ligand, TLR4 may induce pro-inflammatory cytokines released by activating the nuclear factor-kappa B (NF-κB) or p38 pathway [4,5]. Previous studies found that TLR4 antagonist may relieve hyperalgesia induced by nerve injury, chemotherapy drugs or diabetes [6-8] and a genetic defect of TLR4 or its accessory factor CD14 may inhibit activation of glia cells and inflammatory pain [9-11]. However, the role and mechanism of TLR4 in radicular pain from LDH is not clear.
The present study was aimed at confirming the role of the TLR4/NF-κB pathway on radicular pain, and exploring the mechanism of spinal microglia activation and subsequent inflammatory response.
Go to : 
MATERIALS AND METHODS
1. Animals
Sprague-Dawley rats (200-250 g, male) were provided by Guangdong Laboratory Animal Center. Rats were housed in separate cages under a 12-hour light/12-hour dark cycle with free access to food and water. The room temperature and humidity were constant. All animal experimental procedures were carried out in accordance with the guidelines of the International Association for the Study of Pain [12] and were approved by the Animal Care and Use Committee of Guangzhou Medical University (GD2019-143). All efforts were made to minimize animal number and their suffering.
2. Drug administration
TLR4 antagonist ethyl (6R)-6-[N-(2-chloro-4-fluorophenyl) sulfamoyl]-cyclohex-1-ene-1-carboxylate (TAK242, cat no. HY-11109; MedChemExpress, South Brunswick, NJ), which can effectively pass through the blood-brain barrier, was freshly prepared in saline with 5% dimethyl sulfoxide and 5% Tween 80, administered intraperitoneally 1 hour before surgery, and then once daily for 5 days with concentration based on previous studies [13].
For intrathecal administration of drugs, rats were implanted with intrathecal catheters 2 days before surgery according to previously reports [14]. Under anesthesia, a sterile polyethylene-10 (PE-10; Becton, Dickinson and Company, Franklin Lakes, NJ) tube filled with saline was inserted through the L5-L6 intervertebral space upwards until reaching the lumbar enlargement. Any rats with hind limb paresis or paralysis after surgery were eliminated. NF-κB inhibitor pyrrolidinedithiocarbamic acid (PDTC, 15 ng in 10 μL, P-8765; Sigma-Aldrich, St. Louis, MO) or vehicle was carefully injected from the distal inlet of the catheters for 5 consecutive days, beginning from 1 hour before surgery with the concentration based on previous studies [15,16]. Following the drug, sterile saline (7 μL) was given to ensure the drug reaching the subarachnoid space.
3. LDH model
LDH was modeled by autologous nucleus pulposus (NP) implantation described by previous reports [17,18]. The transverse processes were exposed by dissecting the paraspinous muscles from the spinous processes. Hemi-laminectomy was performed in the left L4-L5 segment and the lumbar nerve roots were exposed by carefully removing the facet joint. NP (about 10 mg) was collected from the coccygeal intervertebral discs which were exposed between two vertebral bodies ventrally, and was instantly relocated on the recently exposed nerve roots without any compression. For the sham group, rats experienced the same harvesting step, but the NPs were not implanted.
4. Pain behavioral tests
Mechanical pain thresholds were measured by employing a set of Von Frey filaments (0.41, 0.70, 1.20, 2.04, 3.63, 5.50, 8.51, and 15.14 g) with an up-down method described previously [19]. The first applied stimulus was 2.04 g, if there was no paw withdrawal, the next stronger stimulus was given. If the paw was withdrawn, a weaker stimulus was given. Stimuli were administered to the surface of the hindpaws, lasting 6-8 seconds. Brisk withdrawal or licking of the paw was regarded as a positive response.
Thermal pain thresholds were assessed by a plantar test (cat no 7370; Ugo Basile, Gemonio, Italy) [20]. The plantar surface of the rat’s foot was placed on a glass floor over a radiant heat source. Three values of paw withdrawal latency (PWL) were measured for each animal in each test course. The hindpaws were tested alternately with at least 5 minutes intervals between sequential tests. The three values of PWL per animal were averaged as the result of the test.
Baselines of mechanical and thermal thresholds were assessed repetitively 3 times (day -2, day -1, and day 0) before surgery. Those performing the behavioral tests were blinded to the groups of rats.
5. Western blotting analysis
The L4-L5 spinal cords were harvested and flooded with liquid nitrogen immediately, then the ipsilateral dorsal horn quadrants were separated and homogenized in Tris buffer (15 mmol/L, pH 7.6) containing protease inhibitor cocktail (1:100, cat no. AR1183; Boster Biological Technology, Wuhan, China) and phosphatase inhibitor (1:100, cat no. AR1182; Boster Biological Technology). After homogenization and sonication on ice, the samples were centrifuged at 14,000 × g for 20 minutes at a low temperature. The supernatant was extracted and stored at –80°C before Western blotting.
The total protein concentration of samples was calculated by bicinchoninic acid protein assay. Proteins were separated by gel electrophoresis (sodium dodecyl sulfate-polyacrylamide gel electrophoresis) and then transferred onto a polyvinylidene fluoride membrane (cat no. 1620264; Bio-Rad Laboratories, Inc., Hercules, CA). After blocking for 1 hour at room temperature, the membranes were incubated with TLR4 (mouse, 1:1,000, cat no. sc-293072; Santa Cruz Biotechnology, Inc., Dallas, TX) or β-actin (rabbit, 1:1,000, cat no. 4967; Cell Signaling Technology, Danvers, MA), phosphorylated-p65 (p-p65, rabbit, 1:1,000; cat no. 3033; Cell Signaling Technology), or p65 (rabbit, 1:1,000, cat no. 8242; Cell Signaling Technology) antibody overnight at 4°C. After being washed in phosphate-buffered saline (PBS) repeatedly, the membranes were incubated with goat anti-mouse or goat anti-rabbit horseradish peroxidase-conjugated immunoglobulin G (IgG) (1:5,000, cat no. ab205719 or ab205718; Abcam, Cambridge, UK) for 1 hour at room temperature, then were washed again, as above. The immune complex was detected by ECL kit (cat no. 35055; Pierce Biotechnology, Rockford, IL). The band intensities were quantified by densitometry with a computer-assisted imaging analysis system (IBAS 2.0; Kontron, Augsburg, Germany).
6. Immunofluorescence staining
Rats were sequentially perfused with saline and cold 4% paraformaldehyde (PFA, cat no. 158127; Sigma-Aldrich) in phosphate buffer (0.1 M, pH 7.4). The L4-L5 spinal cords were taken out, post-fixed with 4% PFA solution for 1-3 hours and then dehydrated in 30% sucrose solution for 2 days at 4°C. The spinal cords were sliced into sections transversely with 25 μm thickness by a cryostat (–20°C, CM1900; Leica Biosystems, Wetzlar, Germany) and processed for immunofluorescence staining.
Spinal sections were blocked with 3% donkey serum in 0.3% Triton X-100 (X100; Sigma-Aldrich) for 0.5-1 hours at room temperature and incubated with TLR4 (rabbit 1:400, cat no. AF7017; Affinity Bioscience Pty Ltd, Scoresby, Australia) or ionized calcium binding adaptor molecule-1 (Iba-1, mouse 1:500, cat no. sc32725; Santa Cruz Biotechnology, Inc., Dallas, TX) antibody overnight at 4°C, followed by incubation with goat anti-rabbit Cy3-conjugated IgG (1:400, cat no. BA1032; Boster Biological Technology) or goat anti-mouse fluoresceine isothiocyanate (FITC)-conjugated IgG (cat no. BA1101; Boster Biological Technology) for 1 hour at room temperature. For double immunofluorescence staining, primary antibodies for TLR4 (1:400) were incubated together with anti-neuronal specific nuclear protein (NeuN, neuronal marker, mouse 1:500, cat no. ab104224; Abcam) or anti-glial fibrillary acidic protein (GFAP, astrocyte marker, mouse 1:500, cat no. sc33673; Santa Cruz Biotechnology, Inc.) or anti-Iba-1 (microglia marker, mouse 1:500, cat no. sc32725; Santa Cruz Biotechnology, Inc.) respectively, followed by a mixture of goat anti-mouse FITC- and goat anti-rabbit Cy3-conjugated IgG (1:400). The stained sections were examined with a fluorescence microscope (Leica Biosystems), and images were captured with a charge coupled device (CCD) spot camera (Leica Biosystems). To avoid error, the main parameters such as gain value and exposure time in image acquisition were standardized across sessions.
7. Enzyme linked immunosorbent assay (ELISA)
The dorsal quadrants of the L4-L5 spinal dorsal horn were rapidly harvested and homogenized in PBS followed by centrifugation at 4°C for 15 min at 14,000 × g. The supernatants were collected to detect the concentrations of TNF-α, IL-1β, IL-6, and IL-10 using corresponding ELISA kits (cat no. EK0526, EK0393, EK0412, EK0418; Boster Biological Technology). According to the manufacturer’s instructions, the absorbance was detected at 450 nm (A450) and a standard curve was delineated based on the absorbance of standards.
8. Statistical analysis
All data reported were means ± standard error of means, and were analyzed with SPSS 13.0 (SPSS Inc., Chicago, IL). Western blotting, immunofluorescence and ELISA data were analyzed by one-way analysis of variance followed by Tukey’s post hoc analysis. Data of behavioral tests were analyzed with the unpaired Student’s t-test. The criterion of statistical significance was P < 0.05.
Go to :
RESULTS
1. Spinal TLR4 and p-p65 expression was increased after NP implantation
Western blotting analysis found spinal TLR4 and p-p65 expression was significantly increased from day 3 NP implantation and lasted up to day 14 (Fig. 1A, B: *P = 0.002, **P < 0.001, Fig. 1C, D: *P = 0.001, **P < 0.001). Immunofluorescence staining also confirmed the up-regulation of TLR4 expression (*P = 0.001, **P < 0.001; Fig. 2E-I). More interestingly, the up-regulation of TLR4 mainly appeared in the ipsilateral spine (Fig. 2A-C). Negative control sections, in which only the secondary antibody but not the primary antibody was incubated, were used for testing the antibody specificity (Fig. 2D). Double immunofluorescence staining showed that the TLR4 primarily coexisted with microglia (Fig. 2J-L), but not with astrocytes (Fig. 2M-O) or neurons (Fig. 2P-R).
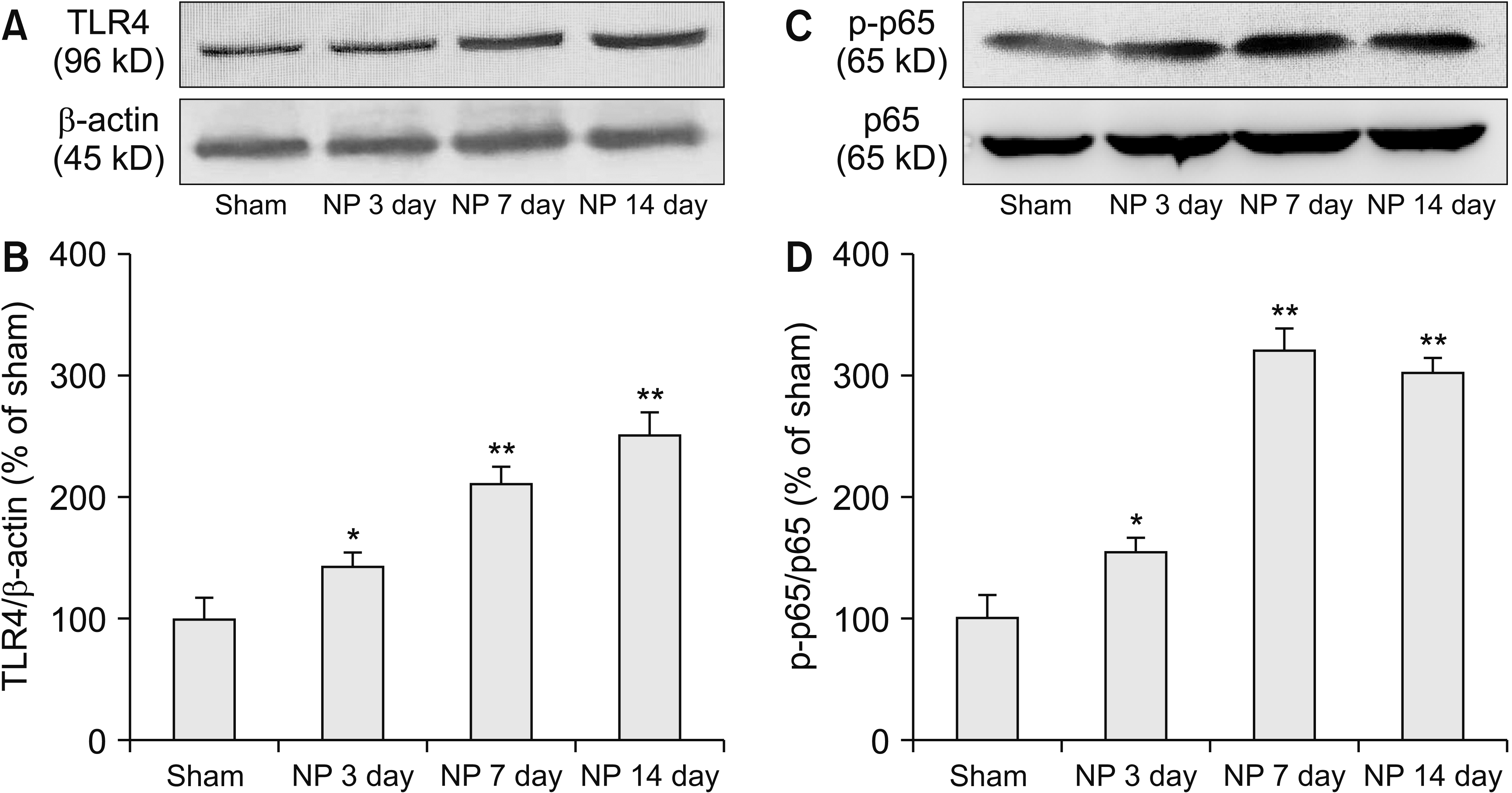 | Fig. 1The protein level of (A, B) toll-like receptor 4 (TLR4) and (C, D) phosphorylated-p65 (p-p65) in spinal dorsal horn of rats with nucleus pulposus (NP) implantation (*P = 0.002 or 0.001, **P < 0.001, n = 5/group). Error bars indicate standard error of means.
|
 | Fig. 2(A-C) Expression of toll-like receptor 4 (TLR4) in bilateral spinal dorsal horn of rats with nucleus pulposus (NP). (D) Specificity of antibody is tested by negative control (n = 2). (E-I) Expression of TLR4 in ipsilateral spinal dorsal horn of different groups (*P = 0.001, **P < 0.001, n = 4/group). (J-R) Double immunofluorescence staining of TLR4 (red) with cellular markers (green): ionized calcium binding adaptor molecule-1 (Iba-1, microglia marker), glial fibrillary acidic protein (GFAP, astrocyte marker) and neuronal specific nuclear protein (NeuN, neuronal marker) respectively. (H) The histogram shows Iba-1 positive area of spinal dorsal horn in different groups. Scale bar (A) = 400 μm, Scale bars (B, C) = 200 μm, Scale bars (D-H, J-R) = 100 μm. Error bars indicate standard error of means.
|
2. TLR4 antagonist and NF-κB inhibitor alleviated pain behaviors of rats with NP
The role of TLR4 and NF-κB in pain behaviors were tested by successive delivery of TLR4 antagonist TAK242 (intraperitoneal [i.p.], 3 mg/kg) or NF-κB inhibitor PDTC (intrathecal [i.t.], 15 ng in 10 μL), beginning from 1 hour before implantation and once daily for 5 days. NP implantation significantly decreased the mechanical PWT and thermal PWL of the ipsilateral, but not contralateral, hindpaws (*P = 0.001, **P < 0.001; Fig. 3A, B) which indicating that NP implantation induced unilateral mechanical allodynia and thermal hyperalgesia. Both TAK242 and PDTC increased the PWT and PWL of rats with NP implantation (vs. vehicle, #P = 0.002; Fig. 3C-F). Therefore, TLR4 and NF-κB may be involved in radicular pain from NP implantation.
 | Fig. 3(A, B) Nucleus pulposus (NP) implantation decreased mechanical and thermal thresholds in ipsilateral but not contralateral hindpaws. Toll-like receptor 4 antagonist TAK242 (TAK) (C, D) and nuclear factor-kappa B inhibitor pyrrolidinedithiocarbamic acid (PDTC) (E, F) alleviate mechanical and thermal pain behaviors of rats with NP (vs. sham, *P = 0.001, **P < 0.001; vs. NP + vehicle [Veh], #P = 0.002; n = 10/group). Error bars indicate standard error of means. PWT: paw withdrawal threshold, PWL: paw withdrawal latency, ipsi: ipsilateral hindpaws, contra: contralateral hindpaws.
|
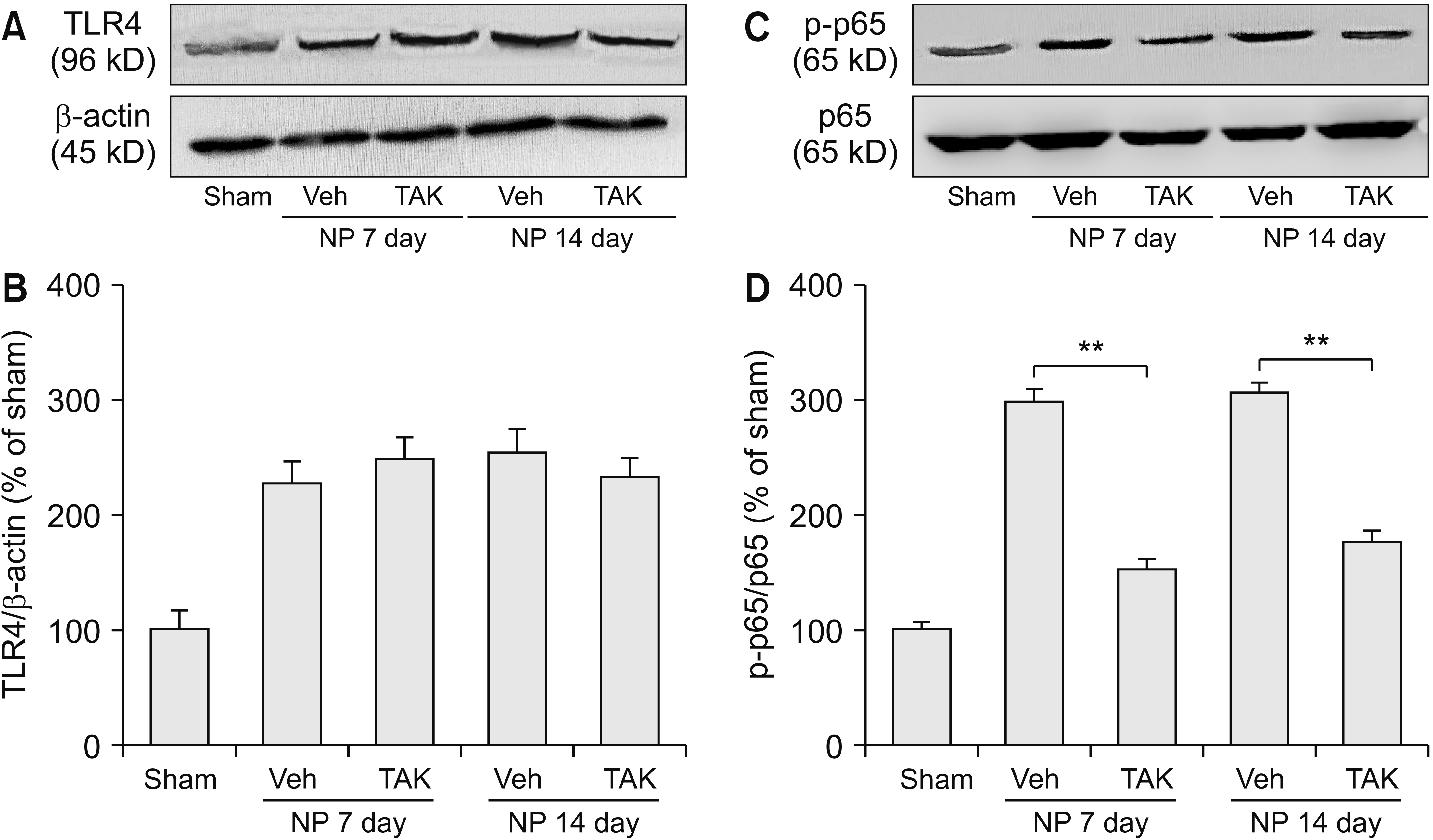
3. TLR4 antagonist TAK242 inhibited NF-κB activation
TLR4 antagonist TAK242 (i.p., 3 mg/kg) was delivered for 5 days and spinal tissues were harvested on day 7 and day 14 after surgery for western blotting analysis. TAK242 evidently weakened spinal p-p65 expression on day 7 and day 14 (**P < 0.001; Fig. 4C, D), without changing TLR4 protein levels (Fig. 4A, B). It indicated NF-κB may be a downstream molecule of TLR4 in the radicular pain model.
4. TAK242 and PDTC inhibited spinal microglia activation and inflammatory response
Spinal tissues were harvested on day 7 after surgery for assessing the effect of drugs on microglia activation and inflammatory response. Microglia-specific marker Iba-1 positive areas in the NP and NP+ vehicle group were significantly increased (vs. sham, **P < 0.001; Fig. 5B, C, D, F, H); TAK242 and PDTC distinctly reduced Iba-1 positive areas (vs. vehicle, ##P < 0.001; Fig. 5D-H). Antibody specificity was also tested (Fig. 5A). Spinal expression of pro- and anti-inflammatory cytokines in different groups was detected by ELISA kits. IL-1β, IL-6, and TNF-α expression was decreased by TAK242 and PDTC (**P < 0.001; Fig. 6A-C), and both drugs increased anti-inflammatory cytokine IL-10 expression (**P < 0.001; Fig. 6D). The data indicated that TLR4/NF-κB activation may promote spinal microglia activation and inflammatory response.
 | Fig. 5TAK242 (TAK) or pyrrolidinedithiocarbamic acid (PDTC) reduces microglia specific marker ionized calcium binding adapter molecule-1 (Iba-1) expression in rats with nucleus pulposus (NP) (vs. sham, **P < 0.001; vs. NP + vehicle [veh], ##P < 0.001; n = 4/group). Scale bars (A-G) =100 μm. (H) The histogram shows the Iba-1 positive area of spinal dorsal horn in different groups. Error bars indicate standard error of means.
|
 | Fig. 6Both TAK242 (TAK) and pyrrolidinedithiocarbamic acid (PDTC) reduce spinal expression of interleukin (IL)-1β, IL-6, and tumor necrosis factor-α (TNF-α), increase IL-10 expression of rats with nucleus pulposus (NP) (**P < 0.001; n = 4/group). Error bars indicate standard error of means. Veh: vehicle.
|
Go to :
DISCUSSION
In the present study, LDH was induced by autologous NP implantation. Spinal expression of TLR4 and p-p65 was significantly increased after NP implantation, lasting up to 14 days. TLR4 was mainly expressed in spinal microglia, but not in astrocytes or neurons. Successive delivery of TLR4 antagonist TAK242 decreased spinal p-p65 expression. TAK242 or NF-κB inhibitor PDTC alleviated pain behaviors and inhibited both spinal microglia activation and spinal inflammatory response. The results demonstrated that the TLR4/NF-κB pathway may be involved in radicular pain through promoting spinal microglia activation and inflammatory response.
Typical symptoms of radicular pain from LDH are unilateral long-lasting low back pain and pain radiating into the leg and foot [21,22]. Initially, it was believed that herniated discs may mechanically compress the nerve roots or spinal cord, which was the source of pain [23]. But this poorly explained the fact that the degree of compression to the nerve roots was not positively correlated with the severity of pain sensation for some patients [23], thus other mechanisms other than compression may be involved in radicular pain. Data from our reports as well as our peers’ reports revealed that levels of spinal pro-inflammatory cytokines and neuroinflammation marker 18 kDa translocator protein were increased after LDH, which demonstrated that neuroinflammation response may be a potential reason of radicular pain [16,24,25]. However, the mechanism of neuroinflammation needs to be further investigated.
TLR4 is a transmembrane protein that is predominantly expressed in microglia of the central nervous system [26,27]. Involvement of TLR4 initiates intracellular signaling pathways, leading to the synthesis and secretion of inflammatory cytokines and chemokines, typically by activating NF-κB and subsequent gene transcription and protein synthesis [4,28]. TLR4 mediated inflammatory response is involved in multiple neurodegenerative diseases such as Parkinson’s disease, Alzheimer disease, and depression [29-32]. The role of TLR4 in neuropathic pain has attracted more attention in recent years. It has been reported that TLR4 antagonist may relieve hyperalgesia induced by nerve injury, chemotherapy drugs, or diabetes [6-8], and a genetic defect of TLR4 or its accessory factor CD14 may inhibit glia cells activation, as well as moderate inflammatory pain [9-11]. In the present study, the role and mechanism of TLR4 on radicular pain is first reported in a rat model of LDH. We find that TLR4 and its downstream molecule NF-κB are upregulated in the ipsilateral spinal dorsal horn of rats with NP implantation (Figs. 1, 2). Successively delivery of TLR4 antagonist or NF-κB inhibitor for 5 days significantly elevate mechanical and thermal pain thresholds that are reduced by NP implantation, and the analgesia effect lasted up to 14 days (Fig. 3). These results clearly confirm the engagement of the TLR4/NF-κB pathway in radicular pain. Interestingly, western blotting analysis finds that intraperitoneal delivery of TLR4 antagonist TAK242 significantly reduces p-p65 expression, but doesn’t change the TLR4 level itself (Fig. 4). As proved by other scientists, TAK242, with high permeability to the blood brain barrier, has evident analgesia effect after both intraperitoneal and intrathecal administration [33,34]. The present result also reveals that TAK242 acts as an antagonist that may block downstream signal of TLR4, but not an inhibitor which inhibits or hydrolyzes TLR4 itself. And this viewpoint is consistent with other reports which revealed that TAK242 decreased TLR4 downstream molecules without changed TLR4 expression in brain tissues in the rat model of migranee [34]. As TLR4 is exclusively located in microglia, we deduce that this pathway may effect microglia activation.
Microglia, acting as resident macrophages in the central nervous system, could release pro-inflammatory cytokines such as TNF-α, IL-1β, and IL-6 when activated in neuropathic pain conditions [35]. Data from our team and other teams has proved microglia activation contributes to radicular pain from LDH [17,36,37]. However, the mechanism of microglia activation is still obscure. The morphological specifics of microglial activation were cell body hypertrophy, thickened and retracted processes, increased cell numbers, and increased staining of microglial specific markers, such as Iba-1 and CD11b [38]. Previously, we proved that spinal microglia were activated mainly in the ipsilateral spine of rats with NP implantation, paralleled with pain behaviors which always appear in the ipsilateral hindpaws [17,37]. In the present study, TAK242 or PDTC obviously alleviate pain behaviors and synchronously decrease the Iba-1 positive area in the ipsilateral spinal dorsal horn, demonstrating that the TLR4/NF-κB pathway may be an important reason for pain development and microglia activation.
What’s more, we find spinal levels of pro-inflammatory cytokines (IL-1β, IL-6, and TNF-α) are reduced and anti-inflammatory cytokine IL-10 expression is elevated by TAK242 and PDTC. The pro-inflammatory cytokines may directly sensitize neurons in the dorsal root ganglion or spinal dorsal horn, which exaggerates pain sensation [16,39-41]. On the contrary, IL-10 down-regulates sodium channels in dorsal root ganglion neurons [42]. Clinical studies found that the serum levels of TNF-α, IL-6, and IL-8 in patients with radicular pain were significantly higher [43-45], and anti-inflammatory cytokine IL-10, but not IL-4, was lower than that of healthy subjects [43]. In an animal model of LDH, expression of IL-1β, TNF-α, and IL-6 in the spinal dorsal horn or dorsal root ganglia was increased [17,46]. The mechanical allodynia was attenuated by inhibition of pro-inflammatory cytokines or using the neutralizing antibody in rats with NP implantation or nerve root compression [47-49]. These reports suggested the imbalance between anti- and pro-inflammatory cytokines may be a crucial reason for radicular pain and restoring balance from upstream molecules may the benefit of relieving radicular pain. Together with the present study, we may deduce that targeting spinal microglia activation and inflammatory response by inhibiting upstream signals such as TLR4 may benefit radicular pain relief.
In summary, our study demonstrates that the spinal TLR4/NF-κB pathway is involved in radicular pain by activating microglia and promoting inflammatory response.
Go to :
 XML Download
XML Download