

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Injury to the peripheral nerves or in the central nervous system is responsible for the induction of neuropathic pain, which is characterized by allodynia and hyperalgesia. The presence of pain symptoms despite the healing of the injury, and being non-responsive to analgesics are the other characteristic features of neuropathic pain [1]. There have been drugs approved to symptomatically manage neuropathic pain; however, the therapeutic management of these patients have still not been optimized. Oleuropein is a polyphenolic compound, and is the main active constituent of olive oil [2]. Research studies have shown the therapeutic potential of oleuropein in several diseases, including depression, Parkinson’s disease, cancer, ischemia-reperfusion injury, metabolic disorders, etc. [2-4]. Apart from these, the inhibitory actions of oleuropein containing leaf extract of Ligustrum vulgare, in diabetic neuropathic pain, have been documented [5]. Moreover, Olea europaea-derived oleuropein is shown to exhibit anti-inflammatory and anti-nociceptive actions [6] and prevent anti-nociceptive tolerance against morphine [7].
H2S is a gaseous neurotransmitter with diverse physiological as well as pathophysiological functions [8,9]. Cystathionine-γ-lyase (CSE) and cystathionine-β-synthase (CBS) synthesize H2S in the body [10], which may impart useful effects in attenuating neuropathic pain in different experimental models of pain including chronic constriction injury (CCI) [11,12] and the oxaliplatin-induced pain model [13]. Orexins are neuropeptides that play a crucial role in sleep, wakefulness, and appetite [14]. Apart from these, studies have shown that orexins may exert analgesic actions in neuropathic pain models, including CCI [15], oxaliplatin [16], and diabetic pain models [17]. The protective role of H2S [11,12] and orexins [15] in neuropathic pain models have been mainly attributed to their actions on the neurons of the dorsal horn and spinal cord. Nuclear factor erythroid-2-related factor 2 (Nrf2), a transcription factor, is very important in maintaining the levels of antioxidants in the body [18]. The decrease in the levels of Nrf2 is associated with the development of neuropathic pain [12].
There have been studies showing that polyphenols may produce their beneficial effects by increasing the levels of H2S [19,20], orexin [21], and Nrf2 [22]. Moreover, there are direct studies showing that oleuropein (polyphenol) increases the expression of Nrf2 [23]. Considering the association of polyphenols with H2S, orexin, and Nrf2, it was hypothesized that oleuropein (polyphenol) may also affect these biochemical molecules to modulate pain. Therefore, the current study was designed to explore the therapeutic usefulness of oleuropein in CCI and vincristine-induced neuropathic pain. Moreover, the possible role of H2S, CSE, CBS, orexin, and Nrf2 was also explored in oleuropein-mediated analgesic actions in male rats.
Go to : 
MATERIALS AND METHODS
1. Animals, drugs, and chemicals
For this study, male Wistar rats (200-250 g) were used. The Institutional Animals Ethics Committee of Yidu Central Hospital of Weifang approved the experimental protocol (approval number: 201900876B097). Suvorexant was purchased from AdooQ BioScience, Irvine, CA; oleuropein and dimethyl sulfoxide (DMSO) were purchased from Sigma-Aldrich, St. Louis, MO. The doses of suvorexant (10 and 20 mg/kg) [24] and oleuropein (10 and 20 mg/kg) [25] were selected on the basis of previously published literature. Suvorexant was dissolved in 10% DMSO, and oleuropein was dissolved in water. Vincristine sulfate was dissolved in normal saline. The drugs oleuropein and suvorexant were administered to CCI, vincristine, and non-treated rats daily, by the oral route. The oral route was chosen to avoid the pain induction that may arise due to a daily injection of drugs. Since the direct administration of drugs by the oral route may also produce stress, drugs were administered by dissolving them in drinking water. To avoid variation in drinking water, drugs were dissolved in a specific volume of drinking water (200 mL of), and care was taken so that animals consumed the entire 200 mL of water.
2. CCI model
Thiopental sodium (40 mg/kg intraperitoneal [i.p.]) was used to anesthetize rats and the sciatic nerve was loosely ligated at four places with a gap of 1 mm, using silk 4-0 thread. Afterward, suturing was performed to cover the muscle and skin. The pain-related behavioral parameters were performed on the day before surgery and on the 14th day after surgery [26]. Amongst the different animal models, the CCI model is very frequently employed in experimental studies [27,28]. In this preclinical model, the ligation of sciatic nerve by four sutures mimics the clinical situation of carpal tunnel syndrome in humans [27,29]. There have been previous studies showing that pain manifestations in this model become predominant on the 14th day [29]. Accordingly, in this study, the pain testing was done on the 14th day following surgery.
3. Vincristine model
Vincristine (50 μg/kg i.p.) was administered for ten consecutive days to induce the development of neuropathic pain [30]. The pain testing was conducted on the 0th day (i.e., a day before starting vincristine administration) and on the 14th day of vincristine administration. Vincristine is an anticancer drug and is very frequently employed for the treatment of cancers [31]. Nevertheless, incidences of neuropathic pain in these patients limit the clinical utility of this anticancer drug [32]. Accordingly, vincristine-induced neuropathic pain is a widely employed preclinical model to explore new therapeutic modalities in pain management in vincristine treated patients [33].
4. Assessment of pain-related behavior
Three pain-related behavioral tests were performed to assess hyperalgesia and allodynia. The behavioral tests were performed in this order: the acetone drop test, the Von Frey hair test, and the pin-prick test, with a gap of 15 minutes between each test. All these tests were performed by placing the rats on an elevated cage with a wire mesh floor.
1) Acetone drop test
The characteristic feature of neuropathic pain is the development of cold allodynia against non-painful (non-noxious) cold stimuli, including acetone. Using micropipette, acetone (100 μL) was applied (from below of the wired mesh) on the plantar surface of the hind paw. The duration for which the animal withdrew its paw was noted in seconds. This test was done three times with an interval of 5 minutes, and the cumulative paw withdrawal duration was noted [34].
2) Von Frey hair test
This test was also performed in the same cage in which the acetone test was performed. In this test, the development of mechanical allodynia was assessed against non-painful stimuli [35] with slight modifications [36,37]. Twenty Von Frey hair filaments (Bio-VF-M; BiosebLab, Vitrolles, France) of varying stiffness, represented in terms of bending forces (0.008, 0.02, 0.04, 0.07, 0.16, 0.4, 0.6, 1, 1.4, 2, 4, 6, 8, 10, 15, 26, 60, 100, 180, and 300 g), were acquired. In this study, the cut off was maintained at 26 g [36] instead of 15.1 g [35]. Since the cut off was fixed at 26 g, the 60, 100, 180, and 300 g filaments were not used in this test. These filaments were touched to the mid-plantar region of the hind paw to induce paw withdrawal. These hairs were applied ten times in ascending order of stiffness i.e., the softest hair was applied first and hardest last. The threshold value i.e., the withdrawal threshold (in grams), was equal to the Von Frey hair stiffness, which evoked 50% paw withdrawal i.e., 5 times out of 10 trials.
3) Pin-prick test (mechanical hyperalgesia)
In the same elevated cage, the pointed end of a traumatic needle was applied to the plantar surface of the hind paw to assess mechanical hyperalgesia. The pain stimulus was applied to the plantar surface with a sufficient intensity to just evoke a paw withdrawal response, without inducing injury to the paw. This duration of paw withdrawal was recorded in seconds, and the minimum paw withdrawal duration was 0.5 seconds [38].
5. Biochemical parameters
After euthenizing the animals, the entire sciatic nerve (from the hip to the foot) was isolated. It was homogenized in phosphate-buffered saline, pH 7.4, followed by centrifugation at 2,000 g, for 15 minutes. Thereafter, the sediments were discarded and the supernatant was selected for biochemical testing.
1) Quantification of H2S
The H2S levels were quantified in the nerve homogenate using reverse-phase chromatography. Thirty μL of the sample was added to 70 μL of 100 mM Tris-HCl buffer (pH 9.5), and thereafter, sulfide was derivatized with excess monobromobimane to form sulfide-dibimane. After 30 minutes, the reaction was stopped by adding 50 μL of 200 mM 5-sulfosalicylic acid. The concentration of sulfide-dibimane was measured using a fluorescence detector (excitation wavelength: 390 nm and emission wavelength: 475 nm) [39].
2) Quantification of CSE, CBS, orexin, and Nrf2
The levels of CSE, orexin, and Nrf2 were quantified in the nerve homogenate using commercially available enzyme-linked immunosorbent assay kits, following the instruction manual. The levels of CBS were quantified using a fluorometric assay kit in which cysteine and homocysteine were added in the homogenate to generate H2S. This was followed by the reaction of hydrogen sulfide with the azido-functional group to yield fluorescence, which was detected using an excitation wavelength of 368 nm and an emission wavelength of 460 nm. Concurrently, the protein content in the homogenates was determined using Folin-Lowery’s method [40]. All biochemical estimations were done in triplicate.
6. Study design
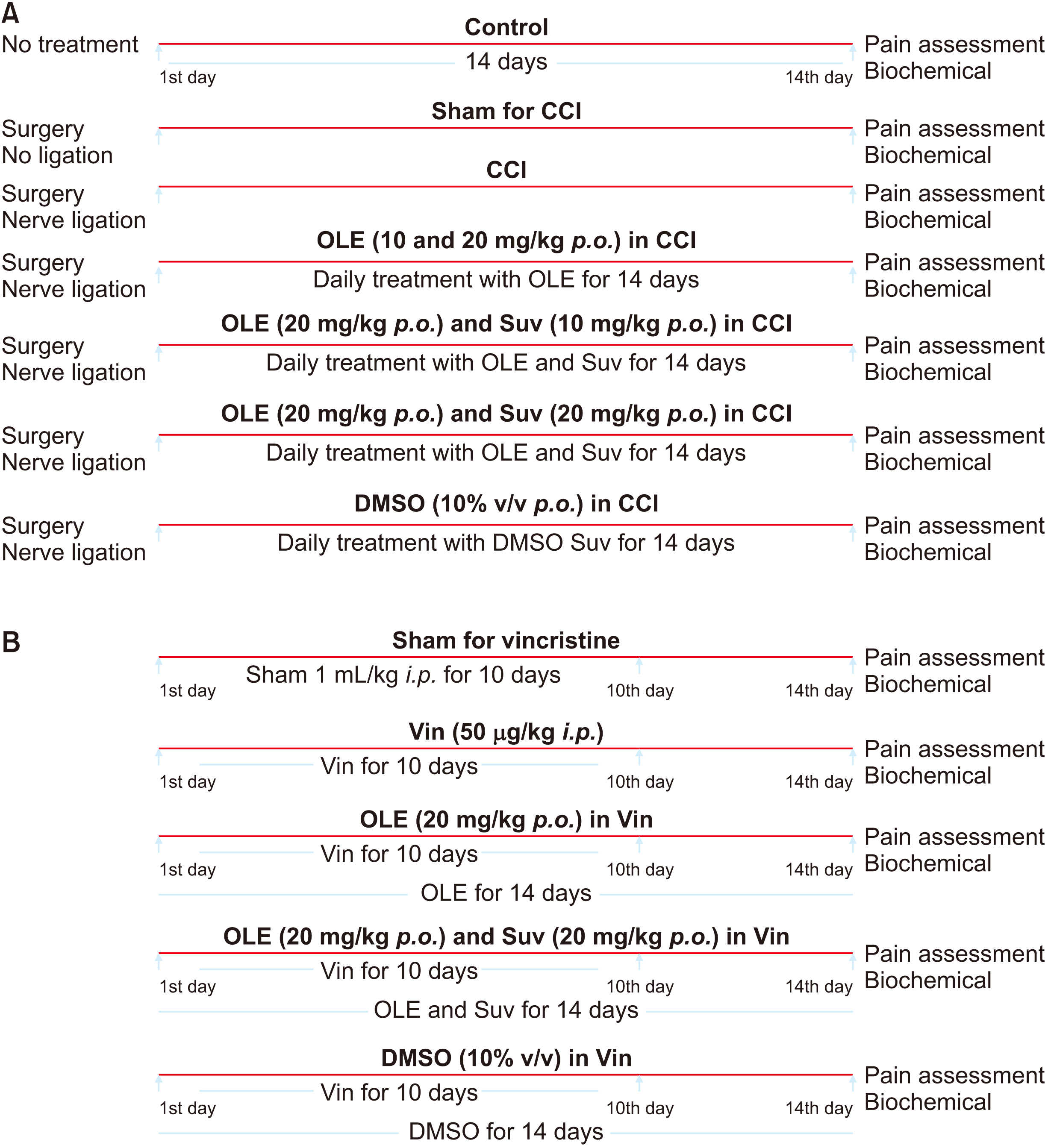
In this experimental study, fifteen groups (n = 10) were used to meet the aim of the study. These groups included a non-treated control (not subjected to any treatment); a sham for CCI (just exposure of the sciatic nerve, no ligation); CCI (the sciatic nerve exposed and ligated); oleuropein (10 mg/kg) in CCI (treatment with oleuropein per oral [p.o.] for 14 days); oleuropein (20 mg/kg) in CCI; suvorexant (10 mg/kg) and oleuropein (20 mg/kg) in CCI (co-administration of suvorexant and oleuropein for 14 days); suvorexant (20 mg/kg) and oleuropein (20 mg/kg) in CCI; a sham for vincristine (administration of saline 1 mL/kg, a vehicle of vincristine for 10 days); a vincristine control (vincristine 50 µg/kg for ten days); oleuropein (20 mg/kg) in vincristine (treatment with oleuropein p.o. for 14 days); suvorexant (20 mg/kg) and oleuropein (20 mg/kg) in vincristine (treatment with suvorexant and oleuropein for 14 days); DMSO (10% v/v) in CCI (administration of DMSO, a solvent of suvorexant in CCI-subjected rats); DMSO (10% v/v) in vincristine (administration of DMSO, a solvent of suvorexant in vincristine-administered rats); oleuropein (20 mg/kg) per se (administration of oleuropein in normal rats); and suvorexant (20 mg/kg) per se (administration of suvorexant in normal rats) (Fig. 1).
7. Statistical Analysis
GraphPad Prism 7 (GraphPad Software Inc, La Jolla, CA) was employed to statistically analyze the data of this study and the data were reported as mean ± standard deviation. The statistical analysis was done using one way analysis of variance. Thereafter, Tukey’s multiple comparison test was used for post hoc analysis. A P value < 0.05 was considered to be statistically significant.
Go to :
RESULTS
1. Development of pain in CCI and vincristine models
CCI and vincristine injection resulted in marked development of pain manifestations as observed on the 14th day, including cold allodynia assessed in the form of an increase in paw withdrawal duration in the acetone drop test (Fig. 2). Moreover, there was a significant development of mechanical allodynia assessed in the Von Frey hair test (Fig. 3) and mechanical hyperalgesia assessed in the pin-prick test (Fig. 4).
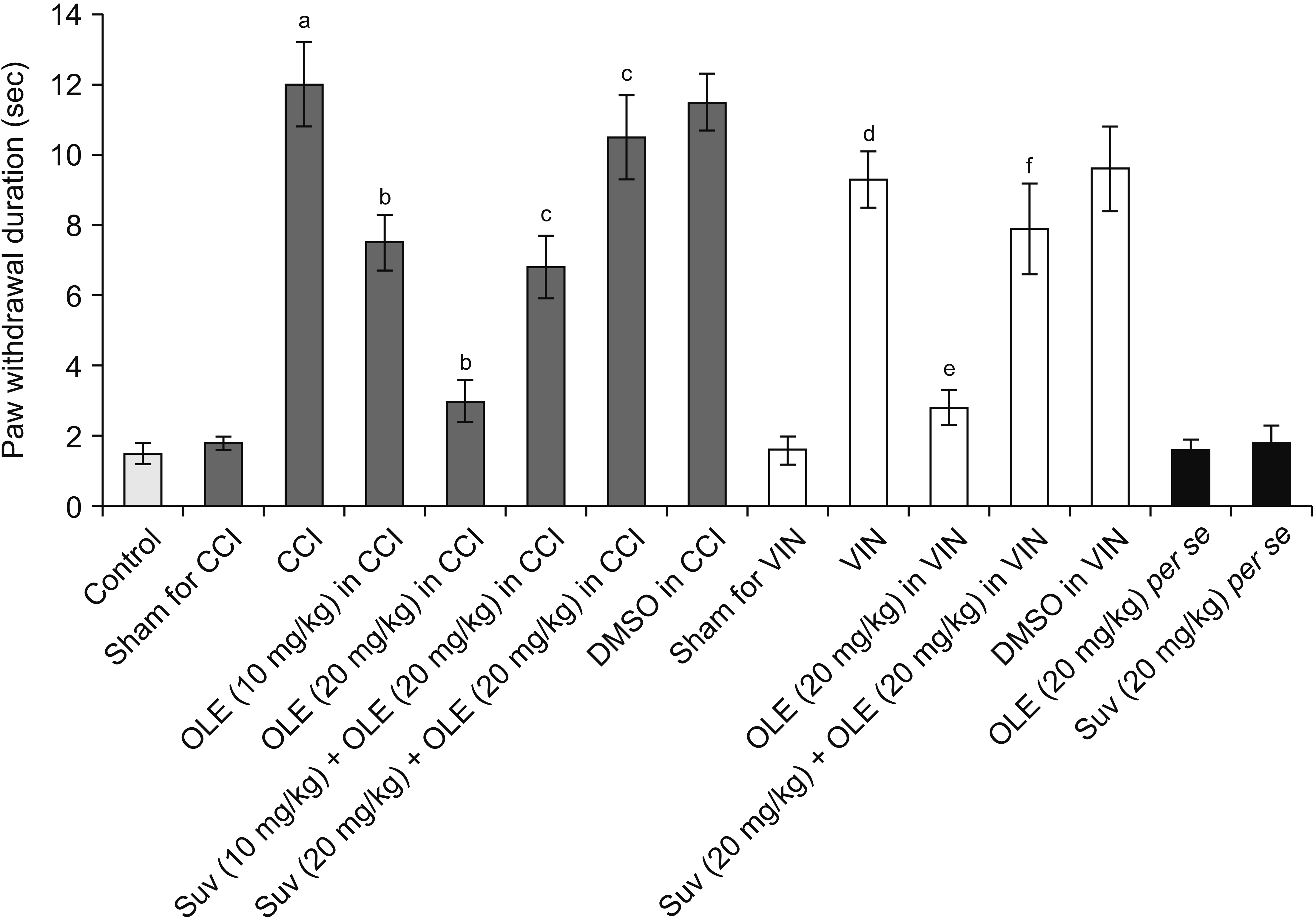 | Fig. 2Effect of different interventions on the cold allodynia in the acetone drop test in chronic constriction injury (CCI) and vincristine models. Values are in mean ± standard deviation. N = 10, F(14, 135) = 342.5. OLE: oleuropein, Suv: suvorexant, DMSO: dimethyl sulfoxide, VIN: vincristine-induced neuropathic pain. aP < 0.05 vs. sham for CCI, bP < 0.05 vs. CCI, cP < 0.05 vs. OLE (20 mg/kg) in CCI, dP < 0.05 vs. sham for VIN, eP < 0.05 VIN, fP < 0.05 OLE (20 mg/kg) in VIN.
|
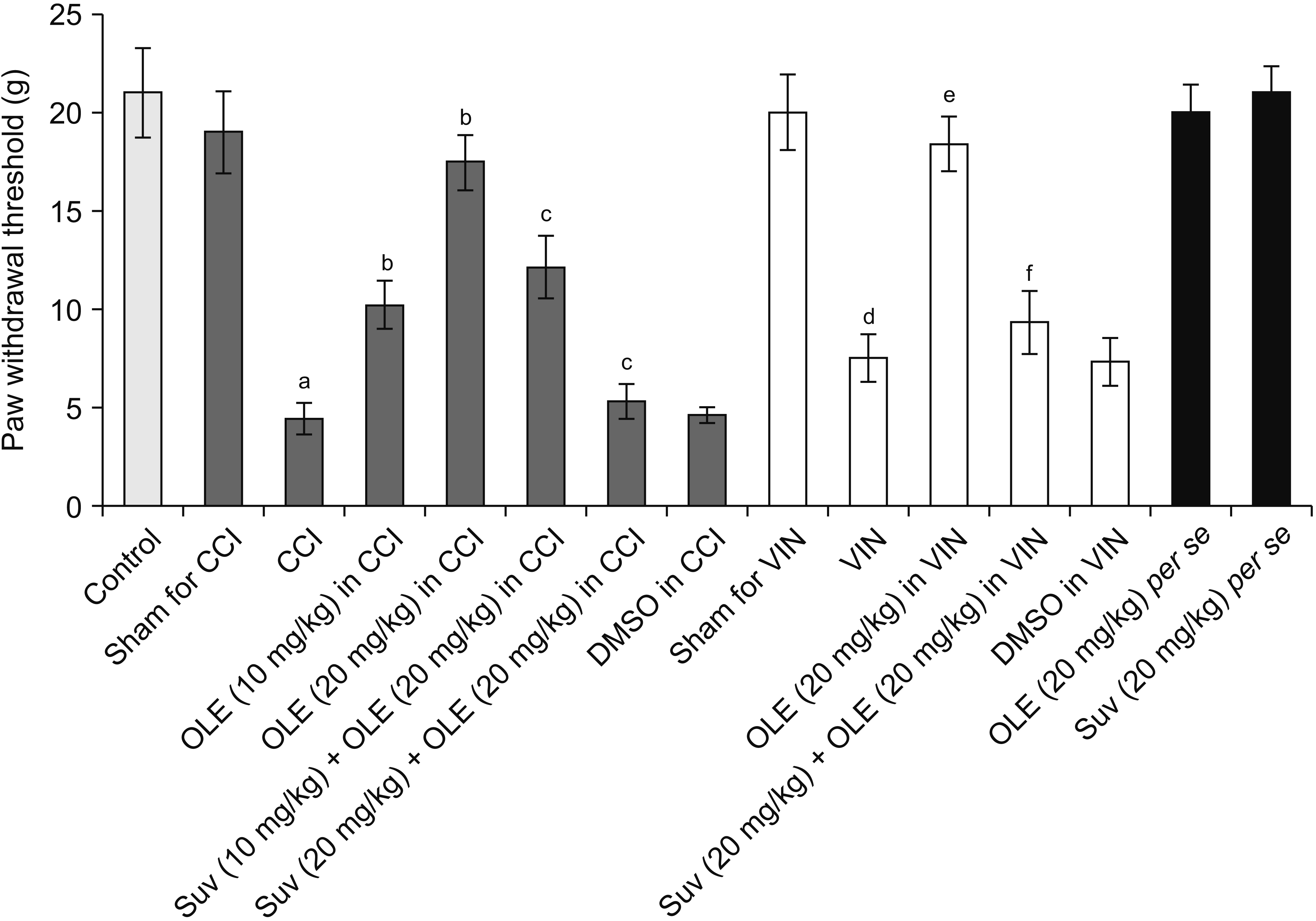 | Fig. 3Effect of different interventions on mechanical allodynia in the Von Frey filament test in chronic constriction injury (CCI) and vincristine models. Values are in mean ± standard deviation. N = 10, F(14, 135) = 372.7. OLE: oleuropein, Suv: suvorexant, DMSO: dimethyl sulfoxide, VIN: vincristine-induced neuropathic pain. aP < 0.05 vs. sham for CCI, bP < 0.05 vs. CCI, cP < 0.05 vs. OLE (20 mg/kg) in CCI, dP < 0.05 vs. sham for VIN, eP < 0.05 VIN, fP < 0.05 OLE (20 mg/kg) in VIN.
|
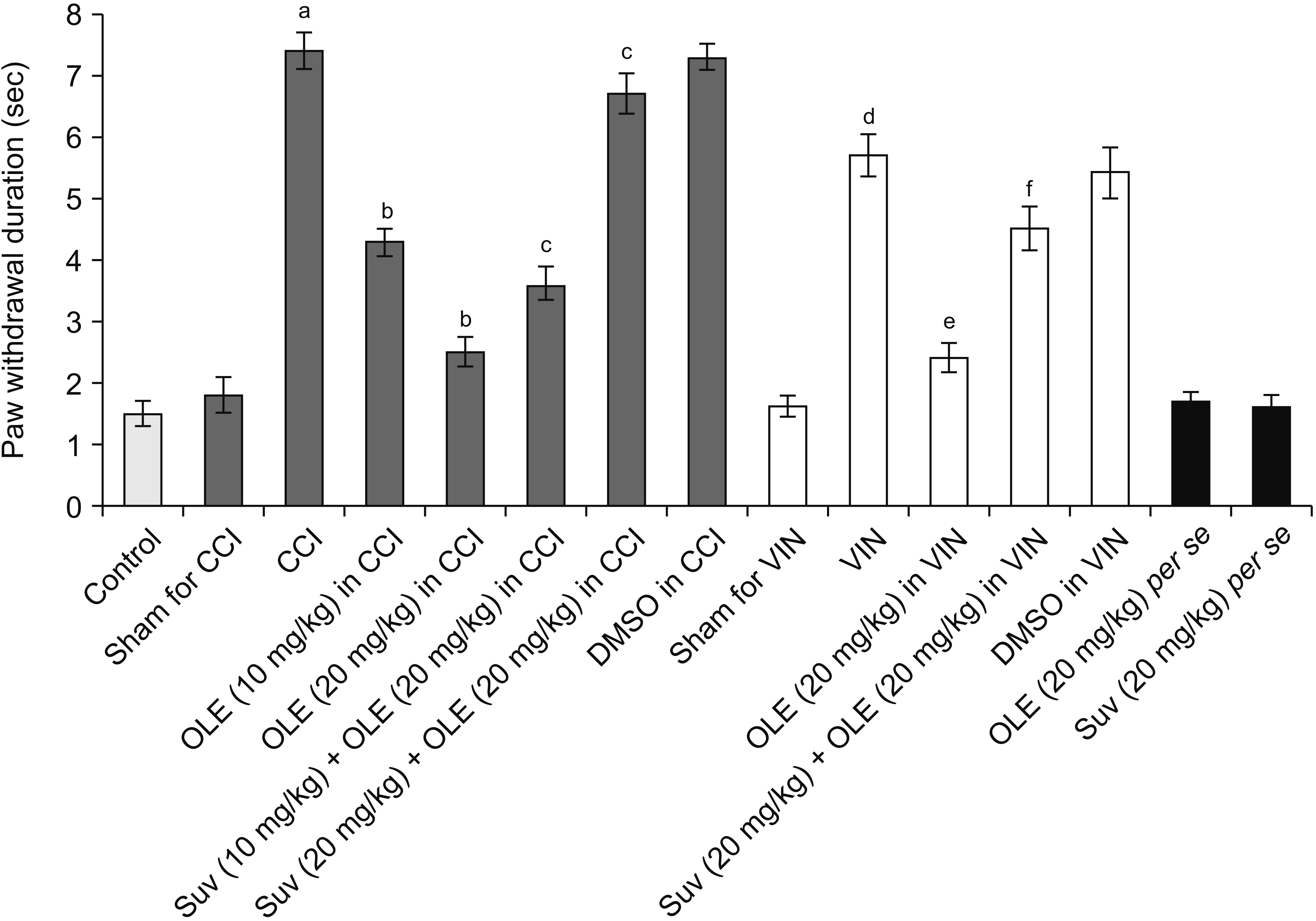 | Fig. 4Effect of different interventions on mechanical hyperalgesia in the pin-prick test in chronic constriction injury (CCI) and vincristine models. Values are in mean ± standard deviation. N = 10, F(14, 135) = 313.8. OLE: oleuropein, Suv: suvorexant, DMSO: dimethyl sulfoxide, VIN: vincristine-induced neuropathic pain. aP < 0.05 vs. sham for CCI, bP < 0.05 vs. CCI, cP < 0.05 vs. OLE (20 mg/kg) in CCI, dP < 0.05 vs. sham for VIN, eP < 0.05 VIN, fP < 0.05 OLE (20 mg/kg) in VIN.
|
2. Attenuation of behavioral manifestations of neuropathic pain in CCI-subjected and vincristine-administered rats with the treatment of oleuropein
Fourteen days of treatment of CCI-subjected rats with oleuropein (10 and 20 mg/kg) reduced the pain manifestations, including a decrease in cold allodynia (Fig. 2), mechanical allodynia (Fig. 3), and mechanical hyperalgesia (Fig. 4). Similarly, the treatment of vincristine-administered rats with oleuropein (20 mg/kg) for 14 days significantly attenuated the pain symptoms. From the results of the CCI-treated group, oleuropein (20 mg/kg) was obtained for the vincristine group.
3. Oleuropein-mediated modulation of biochemical parameters in CCI-subjected and vincristine-administered rats
Significant alterations in the biochemical parameters were observed in the homogenates of the sciatic nerve in both neuropathic pain models. There was a decrease in the H2S levels along with the reduction in the CSE, CBS, and orexin. Treatment with oleuropein for 14 days led to significant elevation in H2S (Fig. 5), CSE (Fig. 6), CBS (Fig. 7), orexin (Fig. 8), and Nrf2 (Fig. 9) in the sciatic nerve of CCI-subjected and vincristine-administered rats.
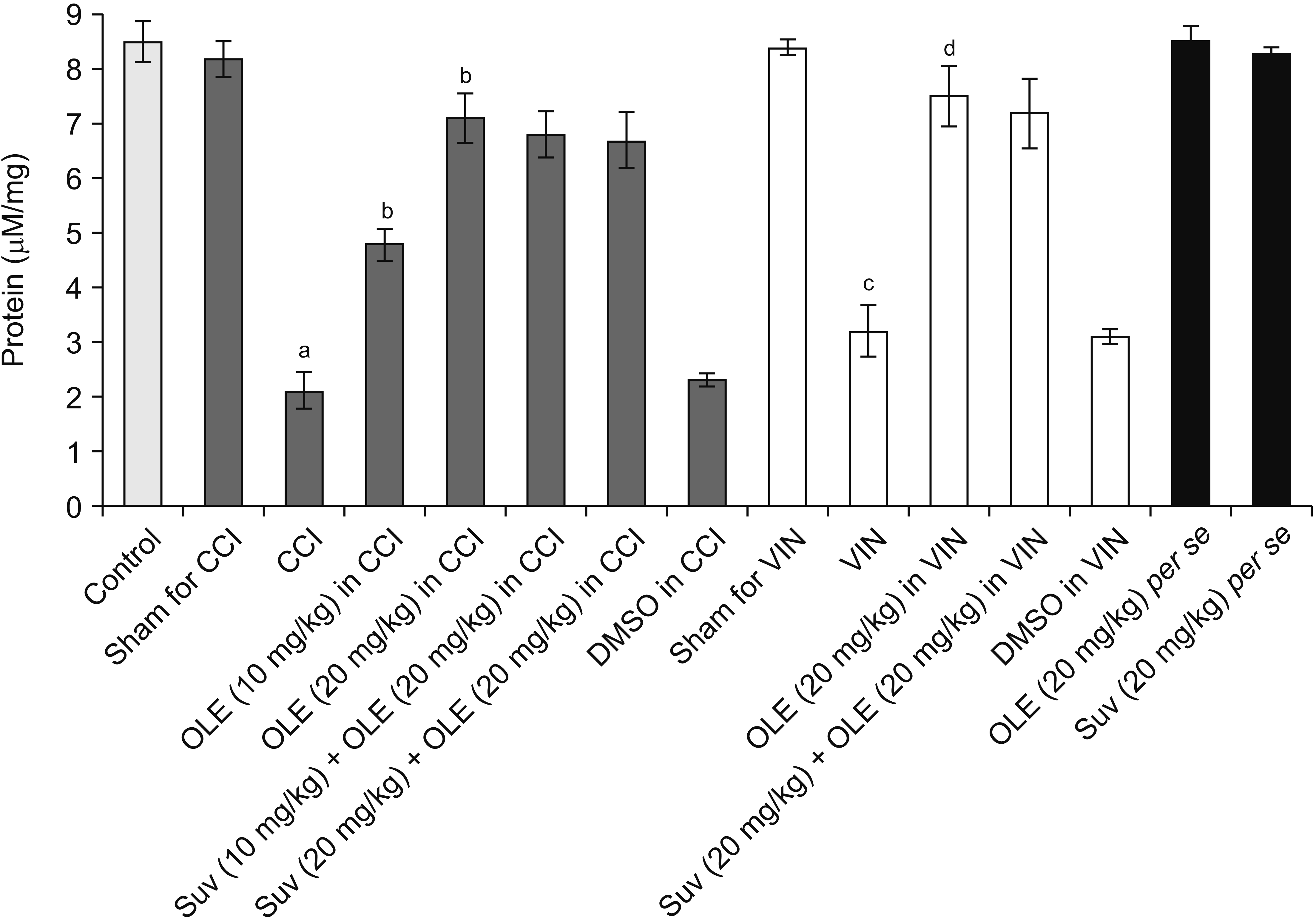 | Fig. 5Effect of different interventions on H2S levels in the sciatic nerve in chronic constriction injury (CCI) and vincristine models. Values are in mean ± standard deviation. N = 10, F(14, 135) = 140.5. OLE: oleuropein, Suv: suvorexant, DMSO: dimethyl sulfoxide, VIN: vincristine-induced neuropathic pain. aP < 0.05 vs. sham for CCI, bP < 0.05 vs. CCI, cP < 0.05 vs. sham for VIN, dP < 0.05 VIN.
|
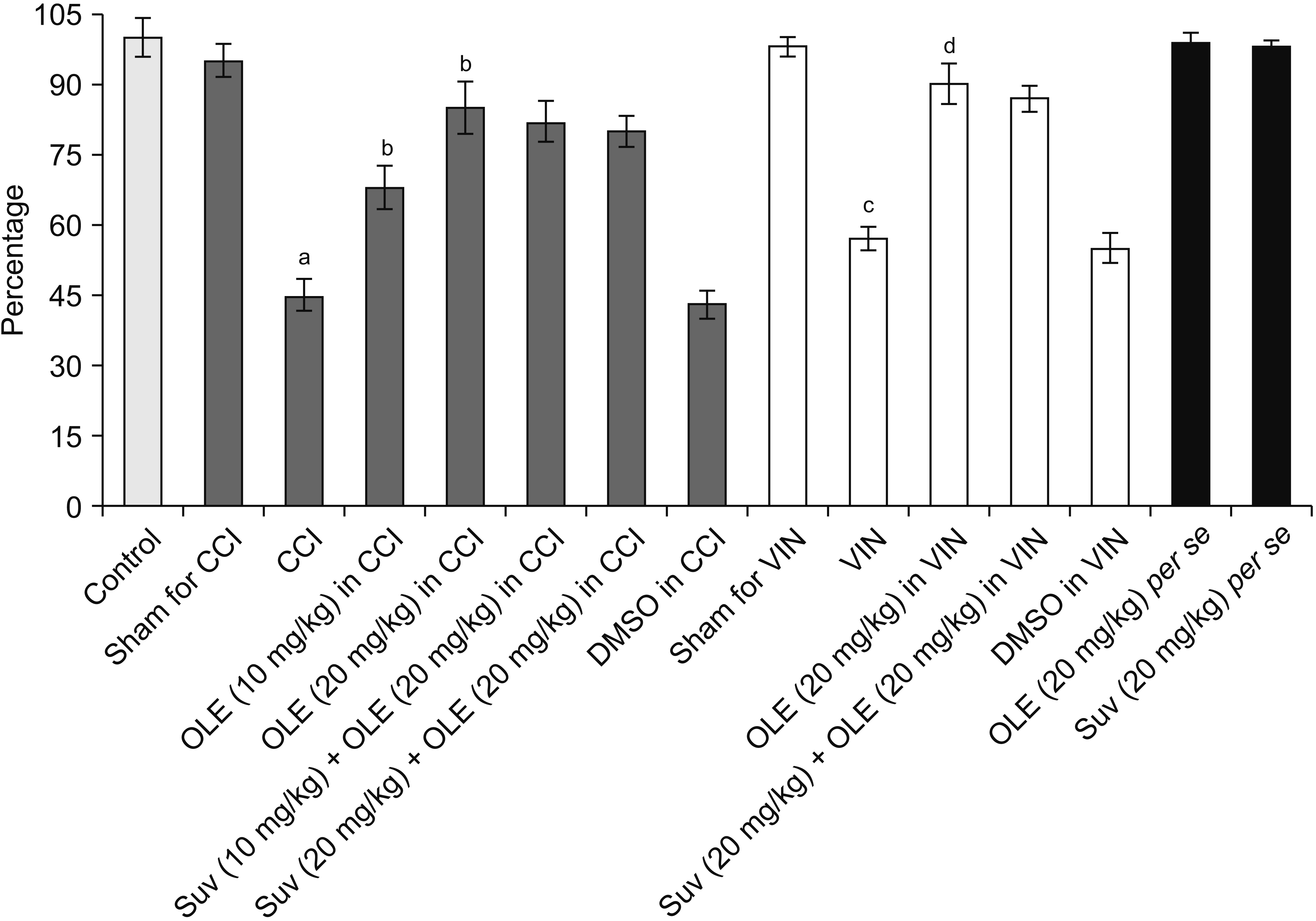 | Fig. 6Effect of different interventions on cystathionine-γ-lyase levels in the sciatic nerve in chronic constriction injury (CCI) and vincristine models. Values are in mean ± standard deviation. N = 10, F(14, 135) = 120.2. OLE: oleuropein, Suv: suvorexant, DMSO: dimethyl sulfoxide, VIN: vincristine-induced neuropathic pain. aP < 0.05 vs. sham for CCI, bP < 0.05 vs. CCI, cP < 0.05 vs. sham for VIN, dP < 0.05 VIN.
|
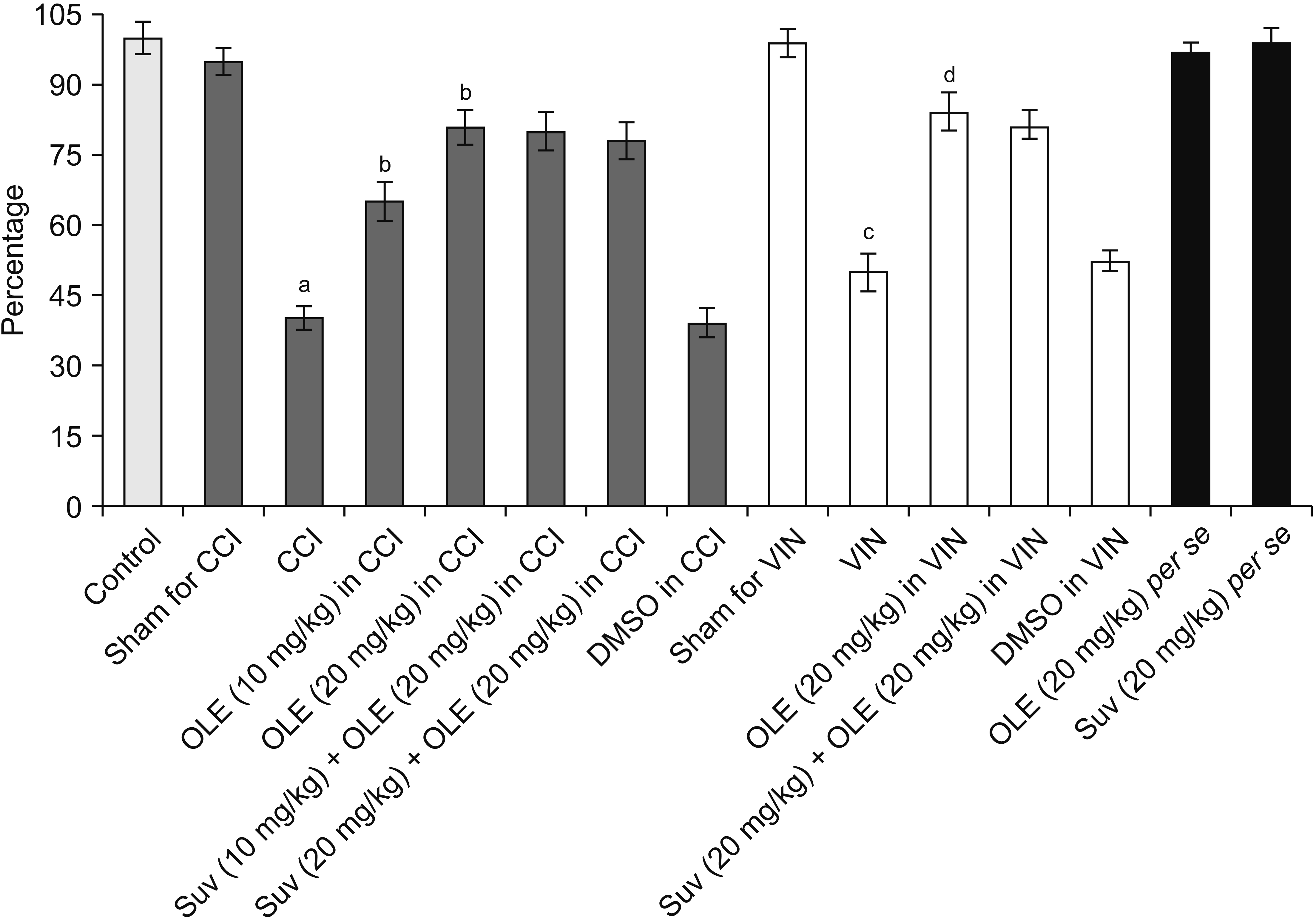 | Fig. 7Effect of different interventions in cystathionine-β-synthase levels in the sciatic nerve in chronic constriction injury (CCI) and vincristine models. Values are in mean ± standard deviation. N = 10, F(14, 135) = 132.7. OLE: oleuropein, Suv: suvorexant, DMSO: dimethyl sulfoxide, VIN: vincristine-induced neuropathic pain. aP < 0.05 vs. sham for CCI, bP < 0.05 vs. CCI, cP < 0.05 vs. sham for VIN, dP < 0.05 VIN.
|
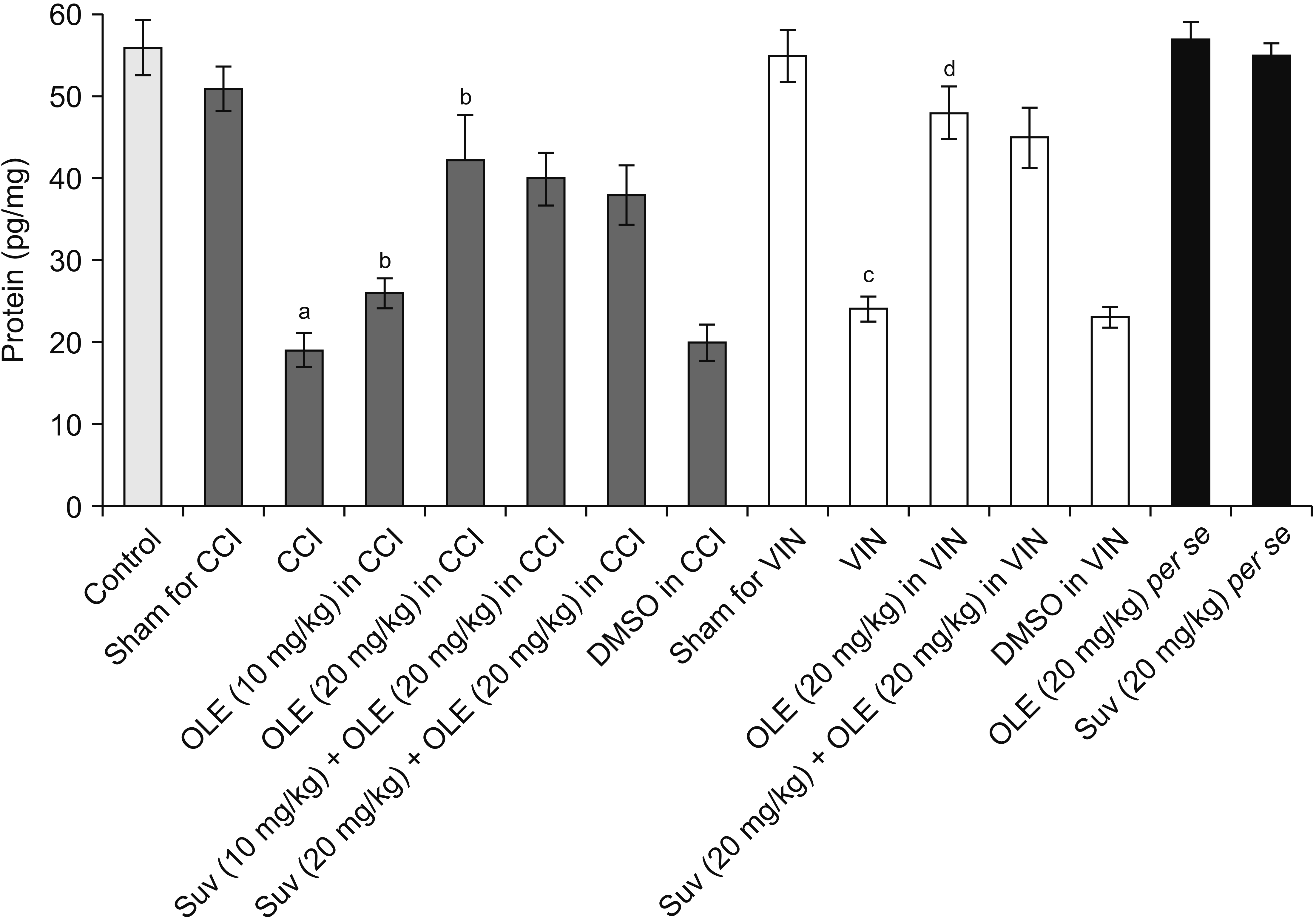 | Fig. 8Effect of different interventions on orexin levels in the sciatic nerve in chronic constriction injury (CCI) and vincristine models. Values are in mean ± standard deviation. N = 10, F(14, 135) = 154.4. OLE: oleuropein, Suv: suvorexant, DMSO: dimethyl sulfoxide, VIN: vincristine-induced neuropathic pain. aP < 0.05 vs. sham for CCI, bP < 0.05 vs. CCI, cP < 0.05 vs. sham for VIN, dP < 0.05 VIN.
|
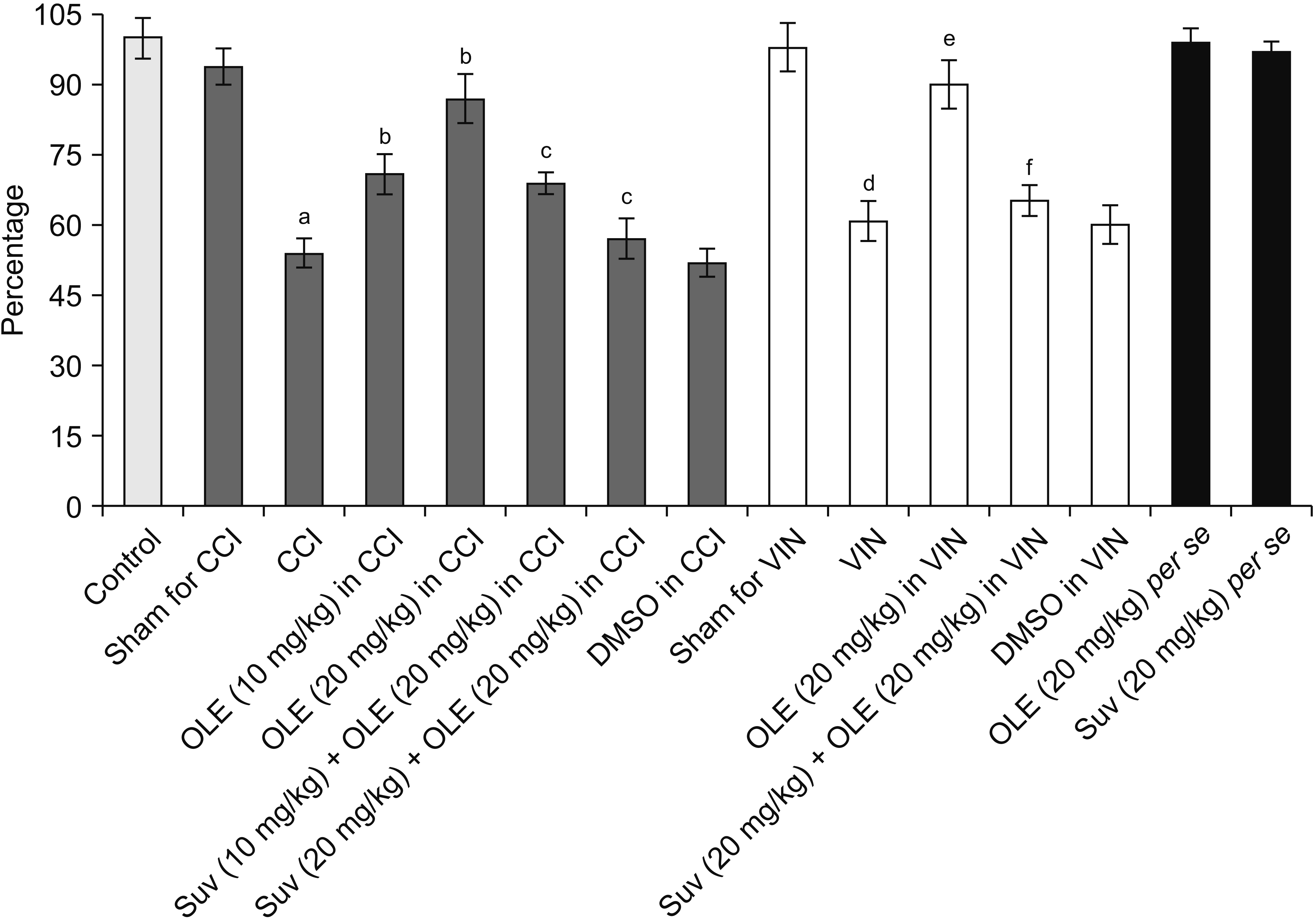 | Fig. 9Effect of different interventions on nuclear factor erythroid-2-related factor 2 levels in the sciatic nerve in chronic constriction injury (CCI) and vincristine models. Values are in mean ± standard deviation. N = 10, F(14, 135) = 144.2. OLE: oleuropein, Suv: suvorexant, DMSO: dimethyl sulfoxide, VIN: vincristine-induced neuropathic pain. aP < 0.05 vs. sham for CCI, bP < 0.05 vs. CCI, cP < 0.05 vs. OLE (20 mg/kg) in CCI, dP < 0.05 vs. sham for VIN, eP < 0.05 VIN, fP < 0.05 OLE (20 mg/kg) in VIN.
|
4. Attenuation of effects of oleuropein in CCI and vincristine models in the presence of suvorexant
Co-administration of suvorexant, an orexin receptor antagonist, significantly attenuated oleuropein-mediated pain attenuating actions in CCI-subjected and vincristine-administered rats. Suvorexant also attenuated oleuropein-mediated restoration of Nrf2 levels in the sciatic nerve (Fig. 9) in CCI-subjected and vincristine-administered rats, without influencing H2S (Fig. 5), CSE (Fig. 6), CBS (Fig. 7), or orexin (Fig. 8).
Go to :
DISCUSSION
In this experimental study, oleuropein exhibited neuropathic pain-attenuating actions and attenuated CCI-induced development of cold allodynia, mechanical allodynia, and mechanical hyperalgesia in male rats. Previous studies have reported the therapeutic potential of oleuropein in preventing cognitive decline [34], cancer, pulmonary inflammation [41], and ischemia-reperfusion injury. Moreover, there have been studies showing the usefulness of oleuropein in diabetic, inflammatory, and nociceptive pain [5,7]. However, this is the first experimental study suggesting the analgesic actions of oleuropein in nerve injury-induced pain. The therapeutic potential of oleuropein in neuropathic pain was further supported by the results showing the attenuation of pain manifestations in oleuropein-treated vincristine-administered male rats.
Along with the development of neuropathic pain, there were significant biochemical changes in the sciatic nerve, including a decrease in the levels of H2S, CSE, and CBS. CSE and CBS synthesize hydrogen sulfide [42], which plays an important role in the pathophysiology of neuropathic pain [13,43]. In this current experimental investigation, treatment with oleuropein restored the levels of H2S, CSE, and CBS in the sciatic nerve in CCI-induced and vincristine-administered rats. It suggests that oleuropein-mediated therapeutic effects in neuropathic pain may be due to an increase in the expression of CSE and CBS, along with the increase in H2S. In this present study, the biochemical alterations were measured in the sciatic nerve (in the periphery), as nerve injury-induced peripheral sensitization is very important in inducing long-lasting pain in neuropathy [44-47]. Therefore, it may be proposed that oleuropein-mediated normalization of the biochemical milieu in the injured nerve may prevent peripheral nerve sensitization in the form of a decrease in abnormal, spontaneous, and ectopic activity. However, there is also a critical role for central changes (neurons of the dorsal horn and others) in inducing neuropathic pain in nerve injury-subjected rats [48]. Accordingly, there is a need for more experimental studies to explore the role of central changes in oleuropein-mediated analgesic actions.
In this investigation, the expression of orexin and Nrf2 were also attenuated in the sciatic nerve in CCI and vincristine models. However, oleuropein restored the expression of orexin and Nrf2 in neuropathic pain models, suggesting that restoration of orexin and Nrf2 may be critical in the pain attenuating action of oleuropein. A decrease in the expression of orexin [16] and Nrf2 [12] is critical in the pathogenesis of neuropathic pain. Oleuropein increases the expression of Nrf2 to produce beneficial effects in different diseases [49]. However, this is the first study showing that oleuropein may produce analgesic actions by increasing the expression of orexin and Nrf2. The key role of orexin in oleuropein-mediated beneficial effects in neuropathic pain models was supported by this investigation, showing that co-administration of suvorexant, an orexin receptor antagonist [50], attenuated the analgesic actions of oleuropein in CCI and vincristine models.
Administration of suvorexant also abolished the oleuropein-mediated increase in the expression of Nrf2, suggesting that oleuropein may increase the expression of orexin, which may subsequently increase the expression of Nrf2. Nrf2 has been reported as the downstream mediator of orexin [41]. However, administration of an orexin receptor blocker (suvorexant) did not attenuate the levels of orexin in oleuropein-treated rats. Generally, the receptor blockers attenuate the biological actions of agonists by preventing their binding to receptor sites, rather than attenuating their biosynthesis. Therefore, it is possible that suvorexant may have attenuated the actions of orexin by preventing its binding to orexin receptors. Nevertheless, more studies may be done to precisely elucidate the reason for the non-modulation of orexin levels in the presence of suvorexant. Moreover, suvorexant did not alter the expression of H2S, CSE, and CBS. In other words, the blockade of the actions of the orexin receptors did not affect H2S, CSE, and CBS, suggesting that orexin is the downstream mediator of H2S. Accordingly, it may be hypothesized that oleuropein may up-regulate the expression of CSE and CBS to increase the H2S levels, which may be followed by an increase in the expression of orexin and Nrf2 to ameliorate neuropathic pain in rats.
Despite the increase in levels of H2S, the lack of any beneficial effects from oleuropein in suvorexant-treated rats also suggests that the rise in the peripheral H2S levels may not be contributing in a significant manner to attenuating pain manifestations. In other words, oleuropein-mediated modulation of orexin and Nrf2 may be more important mechanisms in comparison to the increase in H2S levels. However, the previous studies showing the role of H2S in pain modulation and the present study results showing an increase in the expression of H2S biosynthetic enzymes, along with an increase in the H2S levels in oleuropein-treated rats, also suggest the important role of H2S in oleuropein-mediated pain attenuating actions.
It may be concluded that oleuropein has therapeutic potential to attenuate the pain manifestations in CCI and vincristine-induced neuropathic pain, possibly by restoring the CSE, CBS, and H2S, which may subsequently increase the expression of orexin and Nrf2 to ameliorate behavioral manifestations of pain. Future experimental investigations are needed to delineate the precise role of H2S in oleuropein-mediated pain attenuating actions by employing exogenous H2S donors and selective pharmacological inhibitors of CSE and CBS. Moreover, further studies are also required to clearly delineate the interrelationship between H2S, Nrf2, and orexin in oleuropein-mediated neuropathic pain attenuating actions in rats.
Go to :
 XML Download
XML Download