

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Pain, which is a complicated defensive mechanism and suffering experience, results from painful noxious stimuli that can be associated with tissue injuries. Moreover, it is a typical symptom of most diseases and medical conditions that can affect life quality and personal-social relations. It is estimated that about 3.9% to 48% of adults around the world, among them 31% of Americans, suffer chronic pain [1–4]. Inflammatory pain, a chronic pain by nature, is accompanied by tissue damage and inflammation during which inflammatory mediators are released from the damaged tissue. Specifically, their interaction with nociceptors not only speeds up pain transmission but also readies the nervous system for nociception [5,6].
Opioid receptors are distributed in the central nervous system (CNS) and peripheral neurons and are responsible for the analgesic effects of both endogenous and exogenous opioids during inflammatory and neuropathic pain, and also other painful stimuli [1,7,8]. Additionally, the GABAergic system in collaboration with other systems, such as the opioid receptors, muscarinic acetyl choline receptors, cannabinoid receptor type 1, and adenosine A1 receptors, participates in antinociception both at the spinal and supraspinal levels [9–11].
Due to the chronic nature, physio-psychological consequences, and complications, treatment of chronic pain is much more difficult and less effective than treatment of acute pain, and thus provides pain relief in less than 50% of cases. Additionally, due to the considerable side effects of common analgesics agents, such as opioid analgesics and nonsteroidal anti-inflammatory drugs (NSAIDs), especially in the long term e.g., drug tolerance and gastric ulcers, attention toward new analgesic agents with natural origins and fewer side effects have increased [1–3].
Artemisia annua is a plant of Asteraceae family which has been used over the centuries in Chinese traditional medicine for the treatment of malaria and fever. Artemisinin is a sesquiterpene lactone which was identified and isolated from A. anuua in 1970. Nowadays, it is indicated extensively for the treatment of malaria, and additionally, antifungal, antibacterial, anti-leishmania, anti-coccidian, antidiabetic, antispasmodic, anti-oxidative, anti-inflammatory and wound healing effects from different species of Artemisia and artemisinin have been shown [12–17].
Because of the previously mentioned effects and also the interaction of artemisinin with GABA receptors and its modulatory effects on neuropathic pain [12,18–20], this study was designed to study the analgesic effects of artemisinin on inflammatory pain, and possible role in the GABAergic and opioidergic systems.
Go to : 
MATERIALS AND METHODS
1. Animals
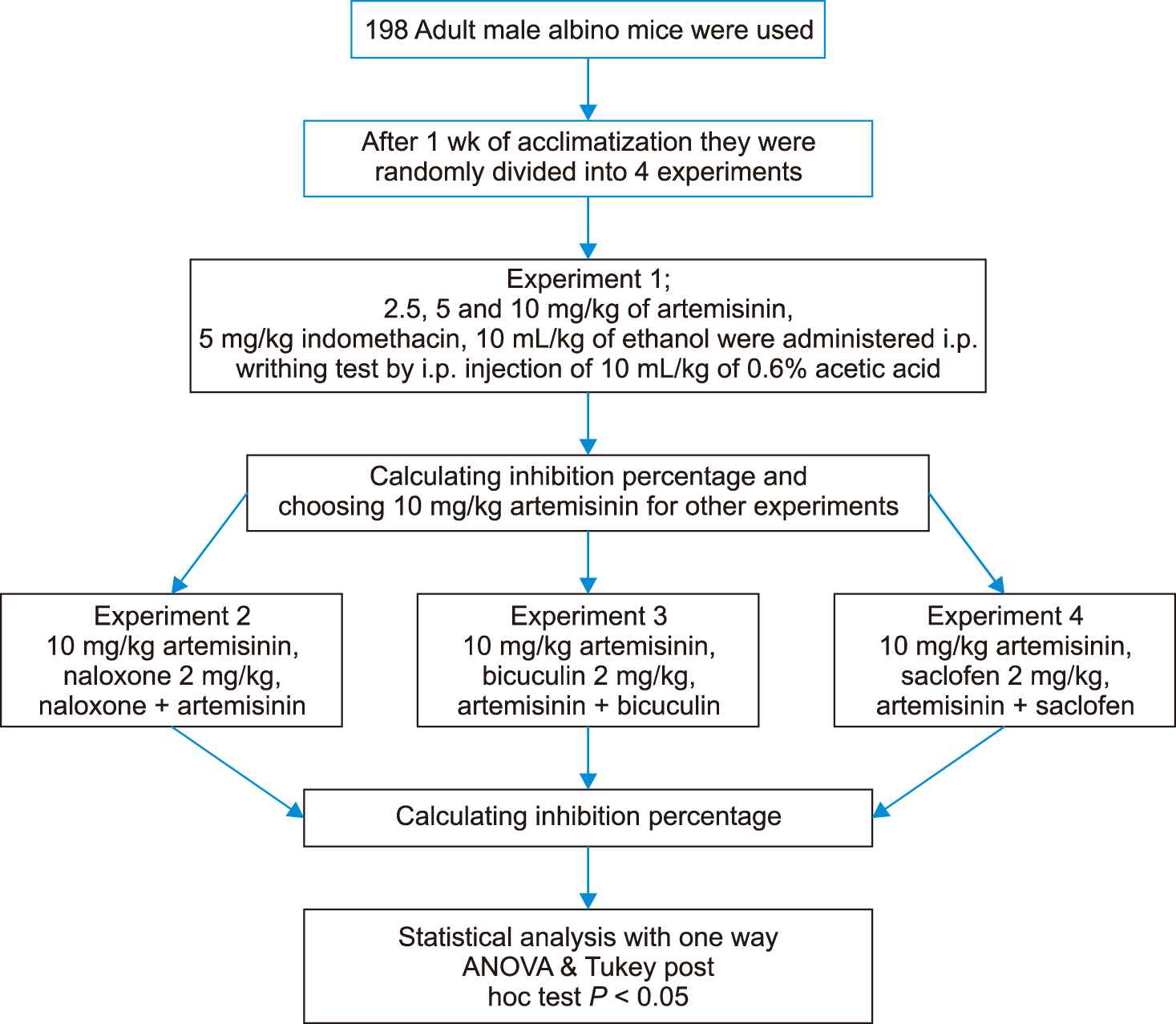
In this study 198 adult male albino N-MRI mice (Pasteur Institute, Tehran, Iran) weighting 25–30 g were prepared and kept under standard laboratory conditions in accordance with the European community guidelines for laboratory animals (23°C ± 1°C ambient temperature, 12 hr dark/light cycle, and 55%–56% relative humidity) in the standard cages with ad libitum access to chow pellets and clean water. After one week of acclimatization, animals were divided randomly into 9 groups (n = 6) each with three replicates in 4 experiments. Experimental procedures were done according to the guide for the care and use of laboratory animals to investigate experimental pain in animals [21], approved by approved by the university ethical committee (559, 2017.06.20). To avoid the possible effects of the animals’ circadian rhythm, all experiments were done from 9 am to 3 pm.
2. Drugs
Artemisinin (Alexis Biochemicals, San Diego, CA) and bicuculline (Sigma-Aldrich, St. Louis, MO) were dissolved in ethanol (Merck, Darmstadt, Germany) so that the final concentration of ethanol was 1.6%, whereas acetic acid (Sigma-Aldrich), indomethacin (Sigma-Aldrich), naloxone (Sigma-Aldrich) and saclofen (Tocris Bioscience, Bristol, UK) were dissolved in saline. All drugs were injected intraperitoneally, and the injection volume was 0.5 mL.
3. Antinociception evaluation
For evaluating the possible antinociceptive effects of artemisinin in the first experiment, 2.5, 5, and 10 mg/kg of artemisinin, 5 mg/kg indomethacin (positive control) and 10 mL/kg of ethanol were administered in each group (Fig. 1). A writhing test was done 30 minutes later by i.p. injection of 10 mL/kg of 0.6% acetic acid. The total number of writhings were recorded for 30 minutes and anti-nociceptive activity was expressed as the inhibition of the percentage of writhings based on the ratio of: (control mean – treatment mean) × 100/control mean [22,23]. Then, the most efficient dose of artemisinin, which statistically was comparable to indomethacin, was chosen for the rest of the experiments. Writhing or abdominal contractions are an excessive extension of the abdominal region in combination with the extension of the hind limbs [22,23]. In the 2nd, 3rd and 4th experiments, first, mice were pretreated with an opioid receptor antagonist (naloxone 2 mg/kg), GABAA receptor antagonist (bicuculline 2 mg/kg), and GABAB receptor antagonist (saclofen 2 mg/kg), respectively. After 15 minutes, artemisinin (10 mg/kg) was administered, and 30 minutes later, the writhing test was done and the inhibition percentage was calculated (Fig. 1). The doses of the drugs used were chosen based on the literature review and preliminary pilot study [23,24].
4. Statistical analysis
Statistical analysis of data was carried out with SPSS ver. 19.0 (SPSS Inc., Chicago, IL). Data were presented as mean ± standard error of mean and analyzed by using one-way analysis of variance followed by the Tukey’s post hoc test (P < 0.05).
Go to :
RESULTS
1. Antinociceptive effects of artemisinin
Administration of 2.5, 5, and 10 mg/kg of artemisinin significantly decreased pain response in comparison to control, to 28.6%, 48.15%, and 63.08%, respectively in a dose dependent manner (P = 0.031, 0.081, and 0.0005, respectively). Indomethacin administration also decreased pain response in comparison to control to 59.35% (P = 0.0007) which was not significantly different with a 10 mg/kg dose of artemisinin (P = 0.902). Thus, a 10 mg/kg dose of artemisinin was used in the rest of experiments (Table 1).
Table 1
Effect of the Artemisinin on Acetic Acid-Induced Writhing Test in Mice (n = 6)
| Treatment | Dose (i.p.) | Writhing count | Inhibition (%) | P value |
|---|---|---|---|---|
| Ethanol (control) | 10 mL/kg | 87.14 ± 4.26 | - | - |
| Artemisinin | 2.5 mg/kg | 62.21 ± 3.71* | 28.60* | 0.031 |
| Artemisinin | 5 mg/kg | 45.18 ± 3.27* | 48.15* | 0.018 |
| Artemisinin | 10 mg/kg | 32.17 ± 2.14* | 63.08* | 0.0005 |
| Indomethacin | 5 mg/kg | 35.42 ± 2.19* | 59.35* | 0.0007 |
![]()
2. Effect of naloxone on antinociceptive effects of artemisinin
Administration of artemisinin (10 mg/kg) induced a significant inhibition in pain response of 63.08% in comparison to control (P = 0.0005). Whereas naloxone did not produce any significant antinociceptive response (P = 0.391), its co-administration with artemisinin (10 mg/kg) induced a significant inhibition in pain response of 57.91% (P = 0.0009), which was not significantly different with artemisinin (10 mg/kg) alone (P = 0.409). This shows that opioid receptor blockade by naloxone has no effect on the antinociceptive effects of artemisinin. Therefore, it seems that, the antinociceptive response of artemisinin is not mediated via opioid receptors (Table 2).
Table 2
Effect of Opioid Receptor Antagonist Naloxone on Artemisinin Antinociception in the Acetic Acid-Induced Writhing Test in Mice (n = 6)
| Treatment | Dose (mg/kg, i.p.) | Writhing count (mean ± SEM) | Inhibition (%) |
|---|---|---|---|
| Artemisinin | 10 | 32.17 ± 2.14 | 63.08 |
| Naloxone | 2 | 75.21 ± 4.52* | 0* |
| Naloxone + artemisinin | 2 + 10 | 34.78 ± 2.25 | 57.91 |
![]()
3. Effect of bicuculline on antinociceptive effects of artemisinin
Table 3 shows that bicuculline does not produce any antinociceptive response (P = 0.561), but bicuculline-artemisinin (10 mg/kg) co-administration significantly reduced the pain response to 26.7% (P = 0.021), which was significantly lower than that of artemisinin (63.08%) alone (P = 0.0005). Therefore, it can be concluded that GABAA receptor blockade by bicuculline diminishes the antinociceptive effects of artemisinin. As a result, the antinociceptive effect of artemisinin may be mediated via GABAA receptors.
Table 3
Effect of GABAA Antagonist Bicuculline on Artemisinin Anti-Nociception in the Acetic Acid-Induced Writhing Test in Mice (n = 6)
| Treatment | Dose (mg/kg, i.p.) | Writhing count (mean ± SEM) | Inhibition (%) |
|---|---|---|---|
| Artemisinin | 10 | 32.17 ± 2.14 | 63.08 |
| Bicuculline | 2 | 78.52 ± 9.45* | 0* |
| Bicuculline + artemisinin | 2 + 10 | 54.37 ± 6.13* | 26.70* |
![]()
4. Effect of saclofen on antinociceptive effects of artemisinin
Administration of 10 mg/kg of artemisinin induces significant inhibition of pain response (P = 0.0005), but saclofen does not produce any antinociceptive response (P = 0.531) (Table 4). Considering the fact that the antinociceptive response to the co-administration of saclofen + artemisinin (57.27%) was not significantly different than with artemisinin alone (63.08%; P = 0.481), it can be concluded that GABAB receptors are not involved in the antinociceptive effects of artemisinin (Table 4).
Table 4
Effect of GABAB Antagonist Saclofen on Artemisinin Antinociception in the Acetic Acid-Induced Writhing Test in Mice (n =6)
| Treatment | Dose (mg/kg, i.p.) | Writhing count (mean ± SEM) | Inhibition (%) |
|---|---|---|---|
| Artemisinin | 10 | 32.17 ± 2.14 | 63.08 |
| Saclofen | 2 | 71.45 ± 8.05* | 0* |
| Saclofen + artemisinin | 2 + 10 | 34.29 ± 5.11 | 57.27 |
![]()
Go to :
DISCUSSION
To the best of the present researchers’ knowledge, this is the first study carried out to investigate the interaction of GABAergic and opioidergic systems on the antinociceptive response to artemisinin during inflammatory pain. Results showed a dose dependent antinociceptive response to artemisinin which decreased following pretreatment with bicuculline GABAA antagonist. Meanwhile, neither saclofen nor naloxone could reverse the antinociceptive effect of artemisinin, which indicates that only GABAA receptors are involved in the antinociceptive action of artemisinin.
The acetic acid writhing test is an animal model of inflammatory pain that has been created for the assessment of the anti-inflammatory/analgesic potency of different compounds [13,16,22]. Intraperitoneal administration of acetic acid induces chemical peritonitis, releases pro-inflammatory mediators, activates peripheral nociceptors on the sensory nerve fibers, and induces abdominal constriction via the synthesis of prostaglandins, especially PGE2, which leads to pain and hyperalgesia. Previous studies have shown that inflammatory pain is responsive to NSAIDs and opioid analgesics [5,6,13,25–29].
Naloxone is a nonselective opioid antagonist for μ, κ and δ receptors which was used to study the probable role of the opioidergic system on the analgesic effects of artemisinin [13,16]. Because naloxone administration could not inhibit the antinociceptive effects of artemisinin, it seems that opioid receptors are not involved in the analgesic effect of artemisinin [1,13,25].
Over the centuries, herbal remedies were the only source for treatment of diseases, including pain, which had an effect due to the presence of herbal metabolites. Sesquiterpene lactones are terpenoids compounds, believed to be the most abundant active herbal metabolites with different biological properties, including anti-inflammatory and analgesic properties [23,30]. It has been reported that artemisinin has prominent immunosuppressive qualities, demonstrated by an inhibition of delayed type hypersensitivity reaction which, interestingly, was more potent than cyclosporine A [17].
Photochemical analysis revealed that different species of Artemisia are rich sources of flavonoid. In the 1980s, the relationship between flavonoid and GABAA receptors was discovered and showed that flavonoids can act on GABAA receptors at a very low concentration as positive allosteric agents. Flavonoids also possess anxiolytic, sedative, and anticonvulsant effects and prevent neurodegenerative lesions of some neural disorders e.g., Parkinson’s and Alzheimer’s which, at least in part, can be mediated by GABAA receptors [18,29]. Inhibition of NF-κB may be involved in the anti-inflammatory activity of the sesquiterpene lactones, because it is a key protein involved in the regulation of cytokines, inflammatory mediators and immune responses [31]. Previously, the profound anti-inflammatory effects of some sesquiterpene lactones, including artemisinin, obtained from different Artemisia species, have been shown [15,18,31,32]. It was reported that the anti-inflammatory effects of Artemisia vestita may be due to the presence of flavone derivatives [12]. Nerolidol is a cyclic sesquitepene compound which exerts analgesic activity via an anti-inflammatory mechanism by suppressing TNF-α, polymorphonuclear leukocytes and proinflammatory cytokines [25]. Dinari et al. [33] reported that the analgesic and anti-inflammatory effects of a methanolic extract of Artemisia aucheri is mainly due to the flavonoids which inhibit the nitric oxide synthase (NOS) enzyme and decrease intracellular calcium as well as calcium dependent PLA2, leading to a decrease in arachidonic acid release, COX activity and, finally, synthesis of inflammatory mediators. Similarly, the analgesic and anti-inflammatory effects of the Artemisia herba-alba essential oil were attributed to the decrease in exudate volume and proteins, as well as the number of inflammatory cells and inhibition of NOS enzyme [13,18].
Bicuculline is a competitive antagonist of GABAA receptors which significantly diminished the antinociceptive effects of artemisinin, whereas the competitive antagonist of GABAB receptors, saclofen, had no inhibitory activity in this regard. Consequently, it can be concluded that GABAA receptors mediate the antinociceptive effects of artemisinin.
Hispidulin and cirsilineol are among the flavonoid compounds isolated from Artemisia herba-alba showing GABAA benzodiazepine receptor activity with IC50 values of 8 and 104 μm respectively [18]. Recently, it was shown that artemisinin interacting with Gephyrin protein as a part of the GABAA receptor complex, activates GABAA receptors and increases GABA signaling. Gephyrin exerts a structural function in the delivery of GABAA and glycine receptors to the cell membrane, playing an important role in their inhibitory action. This effect of artemisinin was antagonized with GABAA antagonists, which confirms the mediating role of the GABAA receptor [19].
In the chronic constriction injury model of neuropathic pain, artemisinin decreases expression of P2X4 receptors and glial fibrillary acidic protein in the satellite glial cells of the dorsal root ganglia, inhibiting nociceptive signaling and inducing analgesia [12]. GABA is the cardinal inhibitory neurotransmitter in the mammalian CNS which involves different functions, such as pain modulation. In fact, GABA exerts its action via ionotropic GABAA and metabotropic BABAB receptors [8–10,25,34–36]. GABAA receptors seem to be the most important and abundant inhibitory receptors in the CNS [11,19]. GABAA signaling is a complimentary part of nociceptive transmission, but during inflammation, GABAergic inhibition of dorsal horn C-fibers is intensified [30,34–37]. In the limbic system, the rostral anterior cingulate cortex participates in the supraspinal processing of nociceptive signals from the periphery, where GABAA binding sites are much more numerous than those for GABAB. It is believed that GABAA receptors mediate synaptic transmission via muscarinic M1 receptors into the ACC region, inducing antinociceptive activity [8,34]. Locally released GABA in the central nucleus of the amygdala is associated with collecting and processing pain information and exerting an inhibitory control mediated via GABAA receptors mainly via increasing chloride conductance which induces neural hyperpolarization, subsequently decreasing neural excitability [9,38].
GABA adjusts pain communication at the dorsal horn via GABAA and GABAB receptors established on the primary afferent terminals in addition to dorsal horn neurons including inhibitory interneurons in the laminae I-III of the dorsal horn. GABAA receptors are recognized on the dorsal root ganglion (DRG) neurons as the primary afferents to the CNS and interneurons in the dorsal horn of the spinal cord, and possess great importance in the gating of afferent sensory signals to the spinal cord where they act both pre- and postsynaptically. At the presynaptic level, GABAA receptor inhibition of afferent sensory inputs is the main tool for blockade of somatosensory signaling to the CNS, while their postsynaptic inhibitory action is assumed as the key element in the regulation of the nociceptive threshold, especially during tissue injury and inflammation. Previously upregulation of GABA content and synthesizing enzymes in inflamed tissues has been shown. It has been reported that inflammation induced by carrageenan or complete Freud’s adjuvant increases the amount of immunoreactive cells and GABA discharge in the spinal cord. Similarly, in the formalin induced pain model, pretreatment with bicuculline inhibited nociceptive behavior, which indicates the role of GABAA receptors in the antinociceptive response to inflammatory pain [39–43].
In conclusion, the results of this study showed dose dependent antinociceptive effects from artemisinin during an inflammatory pain model, which seems to be mediated by GABAA receptors.
Go to :
 XML Download
XML Download