

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Complex regional pain syndrome (CRPS) is the disease appearing after tissue injury or nerve injury, such as trauma, fracture, and burn, and causing neuropathic pain, impairment of the autonomic nervous system and of motor function. Reactive oxygen species (ROS), secretion of cytokine, immunological and inflammatory responses, abnormal responses of the autonomic nerve system, and changes in the central nervous system have been suggested as the cause of CRPS, but the detailed mechanism has not yet been clarified [1].
Recently, Coderre et al. [2] reported a chronic post-ischemia pain (CPIP) model as the necessary model to investigate the pathophysiology of CRPS. This model, which is to induce four weeks of allodynia and hyperalgesia by causing ischemia/reperfusion (IR) injury on the hindpaw of a rat, has a similar pain pattern to that of ischemic injury-related CRPS caused after fractures, arthroscopic surgery, and overly tight casting. Coderre et al. [2] reported that ROS is correlated with the generation of allodynia by decreasing allodynia with the ROS scavenger in this model. In addition, Kwak et al. [3] showed the role of a more characteristic ROS, such as superoxide and nitric oxide (NO), on the generation of postischemic allodynia by using several drugs to block free-radical reaction pathways in a CPIP model. These results correspond with the results from the conventional animal pain model. In other words, in the peripheral nerve injury (spinal nerve ligation) model, it was reported that free radical scavengers, such as phenyl-N-t-butyl nitrone (PBN) and 5,5-dimethyl-1-pyrroline-N-oxide (DMPO), decrease mechanical allodynia [4], and a systemically or intrathecally injected antioxidants reduce the formalin-induced nociceptive response in the hindpaws of mice [5]. In addition, after peripheral nerve injury (sciatic nerve transaction, SNT), the ROS generation in the spinal cord increases while the SOD activity decreases [6], and the mitochondrial ROS inside the spinal dorsal horn neurons increases in the spinal nerve ligation model [7]. These results show the important role of the ROS, increased inside the spinal cord or peripheral tissue, in generating and maintaining chronic persistent pain after nerve injury, inflammatory, or ischemic injury.
In this research, by means of a neuropathic pain model and an inflammatory pain model, the enhancement of N-methyl-D-aspartate (NMDA) receptor phosphorylation by ROS has drawn attention as an important mechanism of central sensitization [8-11]. From this, it is suggested that the development of the allodynia mechanism, expressed in the CPIP model, may result in the increased ROS in spinal cord or peripheral tissue enhances NMDA receptor phosphorylation to induce central sensitization.
Therefore, in this study, we have performed two animal experiments to examine whether the increased ROS is involved in central sensitization through NMDA receptor phosphorylation in CPIP model. First, in order to investigate the role of superoxide produced by xanthine oxidase (XO), which has drawn attention as a main ROS that causes post-IR injury tissue damage in the CPIP model, the degree of NMDA receptor phosphorylation in the spinal cord was measured in the rats, which were given allopurinol (an XO inhibitor) to examine the involvement of superoxide mediated by XO in central sensitization by NMDA receptor phosphorylation. Second, in order to clarify the role of superoxide and nitric oxide, which produce peroxynitrite (known to have strong post-IR injury pro-inflammatory action and cytotoxicity), we have used superoxide dismutase (SOD), which dismutates superoxide and N-nitro-L-arginine methyl ester (L-NAME) (a NO synthase inhibitor), to investigate whether superoxide and NO are involved in the phosphorylation of NMDA receptor and the expression of allodynia that is related with post-ischemia injury.
Go to : 
MATERIALS AND METHODS
All the housing conditions and experimental procedures were approved by the Institutional Animal Care and Use Committee and were in accordance with the National Institutes of Health guidelines on laboratory animal welfare.
Male Sprague-Dawley rats (280-320 g) were housed in the animal room, where temperature, humidity, light, and noise were automatically maintained, and they received water and food ad libitum. They were placed in the animal room for three days prior to the experiment in order to adjust to the environment.
The CPIP model induced by hindpaw IR injury (created by Coderre et al.) [2] was applied to the animals for the experiment. After the rats were anesthetized with sodium pentobarbital, a 7/32 inch of Nitrile 70 Durometer O-ring (O ring West, Seattle, USA) was placed at the rat's left hindpaw just proximal to the ankle joint for three hours to induce ischemia and then the O-ring was removed for reperfusion. The location of the O-ring was standardized as the area near to the medial malleolus. For the sham group (n = 4), the sham operation that did not induce ischemia at the hindpaw was performed by cutting one side of the O-ring.
The experimental group was divided into small groups in which allopurinol 4 mg/kg (LA Group, n = 4), allopurinol 40 mg/kg (HA Group, n = 4), superoxide dismutase 4,000 U/kg (SOD Group, n = 4), N-nitro-L-arginine methyl ester 10 mg/kg (L-NAME Group, n = 4), and SOD 4,000 U/kg and L-NAME 10 mg/kg (SOD + L-NAME Group, n = 4) were intraperitoneally injected respectively, immediately after the O-ring placement, one day after reperfusion and two days after reperfusion. All the chemicals were from Sigma Chemical Co. (St. Louis, MO) and resolved in normal saline just before the injection. For the vehicle group, the same amount of normal saline (500 µl) was injected.
The high dose of allopurinol 40 mg/kg used in the experiment was decided based on the amount used in the previous studies related to IR injury [12-15] and one tenth of it was decided as the low dose; 50 mg/kg allopurinol for rats is one third of the dose for humans in mg/m2 basis [16]. The dose of L-NAME and SOD were also decided based on the conventional research articles related to neuropathic pain model and IR injury [17,18].
Behavioral tests were blindly performed by the tester, who was given no information about the experimental treatment on the rats. In order to observe the behaviors, the rats were placed in a transparent acryl box installed on a wire net. After 15 minutes of adaptation time, the mechanical allodynia was observed.
Dynamic plantar aesthesiometer (DPA, Ugo Basile, Comerio, Italy), operated in an automated von Frey method, was used for the measurement of mechanical allodynia. After the animals for the experiment were completely adapted to the wire net, von Frey filament (straight metal filament, 0.5 mm diameter) was placed at the plantar surface of the ipsilateral hindpaw and the force (Max. 50 g) was increased gradually until withdrawal response, in order to measure the force needed to cause the withdrawal response. The force was applied for four times with a minimum 10 second interval in order to measure the paw withdrawal threshold. The mechanical allodynia was examined and the average threshold was calculated for the contralateral hindpaw in the same manner. The measurement data before the IR injury was set as the baseline value and the ipsilateral and contralateral data were measured on the third day after reperfusion when mechanical allodynia was the maximum [3].
After the mechanical allodynia was measured on the third day from reperfusion, the L4-6 spinal cord was extracted by lumbar laminectomy, separated into left (ipsilateral) and right (contralateral) cord, immediately frozen with liquid nitrogen, and then dissolved in the dissolving buffer solution (20 mM Tris-HCl pH 8.0, 150 mM NaCl, 1 mM EDTA, 2 mM Na3Vo4, 0.5 mM DTT, 10% Glycerol, 1% Nonidet P-40, protease inhibitor cocktail). After centrifugation (12,000 rpm, 4℃, 20 minutes), the supernatants were separated and the proteins were quantified by the Bradford method (Bio-Rad, Hercules, USA). The amount of 50 µg of the whole extracted proteins was mixed with the buffer solution, including a gel loading buffer (0.5 M Tris-HCl, glycerol, 10% SDS, 0.5% bromophenol blue). It was boiled for five minutes at 100℃ and moved to the nitrocellulose membrane after 10% SDS-polyacrylamide gel electrophoresis. The membrane reacted with tris buffered saline (50 mM Tris pH 7.4, 10 mM NaCl), including 3% skim milk, for one hour at room temperature (15-30℃) and then reacted with a phosphorylated NMDA-receptor subunit 1 (pNR1) and antibody (Upstate biotechnology, Temecula, USA) at 4℃ overnight.
The membrane was washed with tris buffered saline (50 mM Tris pH 7.4, 10 mM NaCl) and reacted with a monoclonal secondary antibody conjugated with peroxidase diluted at the ratio of 1:2,000 for one hour at room temperature (15-30℃) to identify the proteins with the ECL system (Amersham Biosciences, Buckinghamshire, England).
The measured data was indicated as the mean ± SEM and analyzed by a paired t-test and analysis of variance (ANOVA) with an SPSS 12.0 program. The post-hoc test was performed with Tukey's HSD method.
Go to :
RESULTS
1. Hindpaw mechanical allodynia
The pre-IR mechanical withdrawal threshold (baseline) measured at contralateral hindpaw and ipsilateral hindpaw had no significant difference between the groups. Although there was no difference with the baseline in the mechanical withdrawal threshold measured in the sham group on the third day after reperfusion, the vehicle group showed significant decrease from the baseline, and the measurement at the ipsilateral hindpaw showed an even lower mechanical withdrawal threshold than that of the contralateral hindpaw (paired T-test, P < 0.05, Fig. 1).
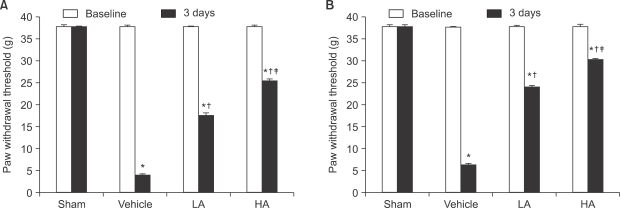 | Fig. 1The mechanical thresholds in the ipsilateral (A) and contralateral (B) hindpaws are measured on 1 hour before ischemia (baseline) and 3 days after reperfusion (3 days). The data are presented as average ± SEM. Sham: non invasive sham control, Vehicle: normal saline i.p., LA: allopurinol 4 mg/kg i.p., HA: allopurinol 40 mg/kg i.p., *P < 0.05 compared with sham group, †P < 0.05 compared with vehicle group, ‡P < 0.05 compared with LA group.
|
When compared to the vehicle group, the LA group and HA group on the third day after reperfusion showed significant increase in the ipsilateral mechanical withdrawal threshold (F3,12 = 10,984.1, P < 0.05, Fig. 1A), as well as the contralateral mechanical withdrawal threshold (F3,12 = 13,519.6, P < 0.05, Fig. 1B). The HA group had a more significant increase than the LA group regarding the ipsilateral mechanical withdrawal threshold (F3,12 = 10,984.1, P < 0.05, Fig. 1A), as well as the contralateral mechanical withdrawal threshold (F3,12 = 13,519.6, P < 0.05, Fig. 1B).
When compared to the vehicle group, the ipsilateral mechanical withdrawal threshold of the SOD group, L-NAME group, and SOD + L-NAME group on the third day after reperfusion had significant increase (F4,15 = 8,414.5, P < 0.05, Fig. 2A), as well as the contralateral mechanical withdrawal threshold (F4,15 = 8,406.0, P < 0.05, Fig. 2B). The ipsilateral mechanical withdrawal threshold of the SOD + L-NAME group on the third day after reperfusion had significant increase when compared to the SOD group and L-NAME group, and the ipsilateral mechanical withdrawal threshold of the SOD group showed significant increase compared to the L-NAME group (F4,15= 8,414.5, P < 0.05, Fig. 2A).
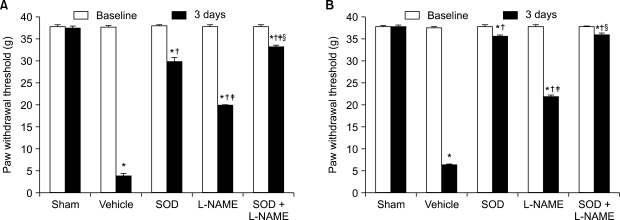 | Fig. 2The mechanical thresholds in the ipsilateral (A) and contralateral (B) hindpaws are measured on 1 hour before ischemia (baseline) and 3 days after reperfusion (3 days). The data are presented as average ± SEM. Sham: non invasive sham control, Vehicle: normal saline i.p., SOD: superoxide dismutase (SOD) 4,000 U/kg i.p., L-NAME: N-nitro-L-arginine methyl ester 10 mg/kg i.p., SOD + L-NAME: SOD 4,000 U/kg plus L-NAME 10 mg/kg i.p., *P < 0.05 compared with sham group, †P < 0.05 compared with vehicle group, ‡P < 0.05 compared with SOD group, §P < 0.05 compared with L-NAME group.
|
The contralateral mechanical withdrawal threshold of the SOD + L-NAME group and SOD group on the third day after reperfusion showed significant increase when compared to the L-NAME group (F4,15 = 8,406.0, P < 0.05, Fig. 2B), while there was no significant difference between the SOD + L-NAME group and SOD group.
2. Measurement of phosphorylated NMDA-receptor subunit 1 (pNR1)
The ipsilateral and contralateral spinal cord pNR1 of the vehicle group showed significant increase when compared with the psilateral and contralateral spinal cord pNR1 of the sham group, respectively. When compared to the vehicle group, the ipsilateral spinal cord pNR1 of the LA group and HA group showed significant decrease (F3,12 = 699.8, P < 0.05, Fig. 3A), as well as the contralateral spinal cord pNR1 (F3,12 = 1045.7, P < 0.05, Fig. 3B). The ipsilateral pNR1 of the HA group decreased significantly compared to the LA group (F3,12 = 699.8, P < 0.05, Fig. 3A), as well as the contralateral pNR1 compared to the LA group (F3,12 = 1,045.7, P < 0.05, Fig. 3B).
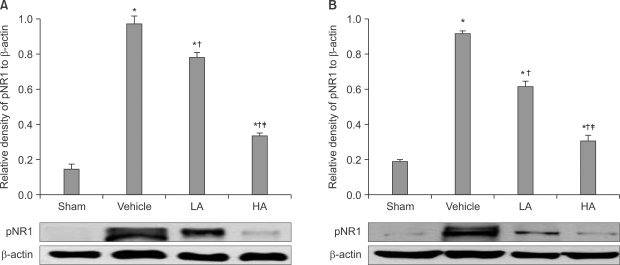 | Fig. 3The average gel density ratio of pNR1 and β-actin in ipsilateral spinal cord (A) and contralateral spinal cord (B) and an example of Western blot gel (bottom). The data are presented as average ± SEM. Sham: non invasive sham control, Vehicle: normal saline i.p., LA: allopurinol 4 mg/kg i.p., HA: allopurinol 40 mg/kg i.p., *P < 0.05 compared with sham group, †P < 0.05 compared with vehicle group, ‡P < 0.05 compared with LA group.
|
The ipsilateral spinal cord pNR1 of the SOD + L-NAME group showed significant decrease with that of the SOD group and L-NAME group, while the ipsilateral spinal cord pNR1 of SOD group showed significant increase compared to the L-NAME group (F2,9 = 635.7, P < 0.05, Fig. 4A). The contralateral pNR1 of the SOD + L-NAME group and SOD group showed significant decrease with that of the L-NAME group (F2,9 = 1,209.1, P < 0.05, Fig. 4B), but no significant difference in contralateral pNR1 was found between the SOD + L-NAME group and SOD group.
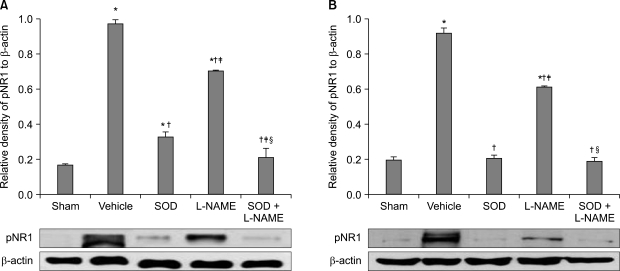 | Fig. 4The average gel density ratio of pNR1 and β-actin in ipsilateral spinal cord (A) and contralateral spinal cord (B) and an example of Western blot gel (bottom). The data are presented as average ± SEM. Sham: non invasive sham control, Vehicle: normal saline i.p., SOD: superoxide dismutase (SOD) 4,000 U/kg i.p., L-NAME: N-nitro-L-arginine methyl ester 10 mg/kg i.p., SOD + L-NAME: SOD 4,000 U/kg plus L-NAME 10 mg/kg i.p., *P < 0.05 compared with sham group, †P < 0.05 compared with vehicle group, ‡P < 0.05 compared with SOD group, §P < 0.05 compared with L-NAME group.
|
Go to :
DISCUSSION
The main findings of this study are as followings: First, in the CPIP model where mechanical allodynia was developed by rat hindpaw IR injury, allopurinol significantly decreased the ipsilateral and contralateral spinal cord pNR1. This suggests that the superoxide mediated by post-IR injury XO may cause ipsilateral and contralateral allodynia by inducing NMDA receptor phosphorylation in spinal cord cells. Second, SOD and L-NAME, which block generation of superoxide and NO, also decreased ipsilateral and contralateral spinal cord pNR1. This shows that increased superoxide and NO are important ROS in causing ipsilateral and contralateral allodynia, as they enhance NMDA receptor phosphorylation in the spinal cord to induce central sensitization.
ROS are involved in an intracellular signal transduction pathway, as the normal derivatives of oxygen metabolism. They are balanced by the antioxidant capacity in the body, but when the balance is lost, ROS rapidly increases and causes oxidative stress. Particularly, during ischemia, hypoxanthine is accumulated by ATP depletion, and superoxide and hydrogen peroxide are generated in the process of hypoxanthine oxidation by XO during reperfusion [19]. Superoxide acts as a proinflammatory by increasing endothelial permeability, generating chemotactic factors, such as leukotriene B4, and inducing neutrophil [20,21]. Reacting with NO, superoxide forms peroxynitrite, which has strong cytotoxicity and acts as a pro-inflammatory substance. Peroxynitrite is involved in post-ischemia cellular injury by continuously generating superoxide, inactivating intrinsic MnSOD by means of nitration [22]. Clarifying the pain-related role of ROS involved in such IR injury, Kim et al. [23] revealed that plasma superoxide mediated by XO was blocked with a high dose of allopurinol before ischemia in the CPIP model; furthermore, Kwak et al. [3] reported that allodynia was reduced as a superoxide and NO was selectively blocked in the CPIP model. Khalil and Khodr [24] reported that the activity of XO, which catalyzes superoxide generation, significantly increased in the sciatic nerve of the CCI model rats and the thermal threshold was decreased. These results suggest that ROS is related to the increase of nociceptor sensitivity, as well as pain transmission pathway and mechanism, and may serve as the proofs that show the action of ROS in peripheral tissue on chronic pain.
The development and maintenance of chronic pain is known to be related with central sensitization. Superoxide and NO have drawn attention as the ROS mainly involved in central sensitization, and the role of superoxide in relation with mitochondrial MnSOD has been focused. Park et al. [7] reported that mitochondrial ROS increased in the spinal dorsal horn of the spinal nerve ligation model. Wang et al. [25] reported that intraplantar injection of carrageenan in rats caused hyperalgesia and increased MnSOD nitration in the spinal cord, and M40403, the SOD analog, decreased hyperalgesia and MnSOD nitration. Schwartz et al. [26] measured mitochondrial superoxide of the spinal dorsal horn in the inflammatory pain model rats (capsaicin induced secondary hyperalgesia) and reported that the increased superoxide was important in the sensory processing of hyperalgesia in the spinal cord and maintained by MnSOD nitration. Alternatively, Xu et al. [27] reported that NO, as a retrograde messenger, released glutamate through the presynaptic cGMP-PKG pathway and increased the neuronal NO synthase (nNOS) activity; then NO itself increased NO generation to neuropathic pain along with continued afferent stimuli. After peripheral nerve injury, release of excitatory amino acids, inclunging glutamate, in the dorsal horn stimulates NMDA receptor; then superoxide and NO are produced by activation of enzymatic cascades through NMDA receptor. MnSOD nitration by peroxinitrite, reactants of superoxide and NO, maintain high level of superoxide, then spinal cord hypersensitivity and central sensitization mediated by NMDA are increased [8,28].
Physiologically, spinal cord central sensitization is defined as the increased reactivity of the spinal dorsal horn to the peripheral nociceptor stimuli [29], and induction of the spinal cord central sensitization is well known to be related with NMDA receptor activity. The NMDA receptor is a complex, and five NMDA receptor subtypes have previously been categorized. Among them, NMDA receptor subtype 1 is the subtype that constitutes functional receptor channel complex and plays the original function of the NMDA receptor. The NMDA receptor is activated by phosphorylation of NMDA receptor subtype 1 [30,31]. In spinothalamic tract of the inflammatory pain model, pNR1 is increased by the intradermal injection of capsaicin [9]. In the inflammatory pain model, injection of NMDA induced by formalin increases pain behavior, and the NMDA receptor antagonist decreases hyperalgesia [10]. Gao et al. [8,11] reported that pNR1 immunoreactive neurons increased in the spinal dorsal horn of rats with nerve injury, and the decrease of pNR1 immunoreactive neurons in the spinal dorsal horn is consistent with the decrease of hyperalgesia after the injection of PBN, which was the antioxidant in the neuropathic pain model and inflammatory pain model. They thus concluded that ROS is involved in the central sensitization through NMDA receptor activation.
In this study, the result that pNR1 was decreased by blocking the superoxide mediated by XO, as well as the superoxide and NO (the precursors of peroxynitrite), showed that ROS is involved in the central sensitization mechanism related with the NMDA receptor in this IR injury model, as in the previous reports based on the inflammatory pain model and neuropathic pain model [8-11], and central sensitization that contributes to continued neuropathic pain, such as mechanical allodynia, can be reduced by blocking superoxide and NO. However, NO is the type of molecule that can have dual effects depending on its concentration, NOS isoform, and redox balance, etc [32,33]. As contrasting research results show that NO is the molecule acting on the development of neuropathic pain, No has been reported to be involved in analgesia. With respect to this, there are reports that the L-arginine-NO pathway was involved in peripheral and central analgesia [34], NO reduced hyperalgesia by carrageenan, and NO donor, such as nitroglycerine, reduced hyperalgesia [34,35]. In addition, the L-arginine-NO-cGMP pathway was reported to be involved with analgesia of morphine, clonidine, codeine, ketamine, and gabapentin, etc [34-41]. Furthermore, Xanthos et al. [42] reported that the NOS inhibitor and NO donor injected on the second day after reperfusion in the CPIP model relieved vasoconstriction and ischemia, and thus, reduced the pain. Therefore, in the post-IR injury animal pain model of this study, blocking NO, which is the precursor of peroxynitrite, can contribute to pain development by reducing peripheral and central sensitization and/or cancel the vasoconstriction and ischemia-relieving effect of NO through an alpha receptor. In particular, blocking a superoxide was more effective than blocking NO in reducing the expression of mechanical allodynia and central sensitization, which is assumed to be the possible action of NO which has such dual effects.
Of the CRPS-I patients, 10% showed mirror image pain, which is diffused to other limbs that are not injured [43], and allodynia and hyperalgesia were also found in a contralateral part in the CPIP model [2,3,23]. Mirror image pain is found not only in the CPIP model, but also in the neuropathic pain model and inflammatory pain model, and such theories as central mechanism and humoral mechanism have been postulated. One suggested opinion is that activation of a glial cell in the spinal cord and the discharge of cytokine may cause the distant or mirror image pain [44,45], and Gordh and Sharma reported that structural change was observed in the contralateral spinal cord two weeks after the nerve injury and continued for eight weeks [46]. Since sensory response in the peripheral afferent nerve is not directly projected to the contralateral spinal dorsal horn [47], the contralateral sensitization in the spinal cord is assumed to be caused by commissural interneurons [48,49]. In the recent study of Kwak et al. [50], the mirror image pain in the CPIP model was reportedly due to central sensitization related to the activation of the NMDA receptor; and through this study, the mechanical allodynia by such contralateral central sensitization was confirmed to be caused by ROS.
The two possibilities that must be considered are that the ROS scavenger, which was intraperitoneally injected before and after the IR injury, when mechanical allodynia was generated, may have reduced the afferent stimuli that cause central sensitization and thus central sensitization in spinal cord, or reduced sensitization of the spinal cord through direct blocking of superoxide and NO in the spinal dorsal horn cell. However, the exact part of the action could not be determined. Thus, various approaches, including intrathecal drug injection, should be used in the study to clarify the exact action parts of the pain mechanism where ROS acts in the post-IR injury animal pain model.
In conclusion, superoxide by xanthine oxidase, and superoxide and NO, which are the precursors of peroxynitrite, are assumed to be involved in central sensitization related to NMDA receptor phosphorylation in the CPIP model and contribute in causing and maintaining mechanical allodynia.
These results suggest the possibility of using ROS scavengers, such as SOD and L-NAME blocking superoxide and NO, which are the precursors of allopurinol and peroxynitrite, as effective therapeutics for patients of chronic pain related to ischemic injury.
Go to :
 XML Download
XML Download