

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Chemotherapy-induced peripheral neuropathy (CIPN) is one of the most severe and long-lasting side effects caused by chemotherapeutic agents [1]. Symptoms of CIPN, such as abnormal touch sensation, altered responses to thermal stimuli, and pain, which are often dose dependent and progressively worse during and after chemotherapy [2], typically develop in the “glove and stocking” areas of the distal extremities [3]. Although medications have been employed to treat the symptoms associated with CIPN, few medications have been consistently effective or tolerable [4-6]. Also, despite intense efforts, no definite analgesics have been developed to treat CIPN.
Neurotensin (NTS) is an endogenous 13-amino acid peptide, originally isolated from bovine hypothalamus [7]. This neuropeptide and its receptors are widely distributed in the central nervous system (CNS), including sites important for pain transmission and modulation [8]. The multiple effects of NTS derive from its interaction with three different receptors (NTSR1, NSTR2, NSTR3). Microinjection of NTS, or NTS analogs, to specific brain areas exerts various biological effects through interactions with neurons secreting dopamine, serotonin, or norepinephrine [9-11]. Furthermore, NTS1 agonists have shown antinociceptive effects in various pain models. For example, intrathecal injection of NTSR1 agonist dose-dependently attenuates formalin-induced pain behavior in rats [12], and systemic administration of NTSR1 analogs mediates the visceral analgesic effects of NTS in the writhing model [13]. Moreover, intrathecal delivery of NTSR1 agonists reversed heat hyperalgesia and tactile allodynia induced by chronic neuropathic pain models, such as sciatic nerve constriction [14].
Serotonin (5-hydroxytryptamine, 5-HT) is involved in either facilitatory or inhibitory nociceptive transmission via the activation of different 5-HT subtype receptors [15], which is dependent on the physiological or pathophysiological status of the experimental animal. Recent studies have shown that 5-HT1A has a protective effect against CIPN-related pain [16], and brain NTSR1 activation induced serotonergic neuronal activity and 5-HT release [11,17,18]. Also, the antinociceptive effect of NTSR1 agonist administered to the rostral ventromedial medulla (RVM) on noxious thermal stimulus was inhibited by intrathecally administered non-selective serotonergic antagonist [9,10]. These results suggest that the antinociceptive effects of NTSR1 agonist are mediated by interactions between NTSR1 and neurons that release 5-HT. Thus, the aim of the present study was to assess the role of NTSR1 in antinociception and to determine its association with the serotonergic pathway in a CIPN animal model at the spinal level.
Go to : 
MATERIALS AND METHODS
1. Animals and the chemotherapy-induced neuropathic pain model
Male Sprague–Dawley rats (weight 150-180 g) were purchased for this study. The animals were acclimated to the laboratory for 3-5 days before use. They were allowed free access to a standard rat diet and tap water in their home cage environment. The room was maintained at 24℃ with a 12-hr/12-hr light/dark cycle. This study proposal was reviewed and approved by the Institutional Animal Care and Use Committee of Chonnam National University (IACUC-H-2016-47). The rats were injected intraperitoneally with 2 mg/kg cisplatin for 4 days to induce the CIPN model, as described previously [19].
2. Implantation of an intrathecal catheter for the injection route
A length of 8.5 cm polyethylene-10 tube was inserted into the subarachnoid space under sevoflurane anesthesia through a small incision in the atlantooccipital membrane 5 days after the first cisplatin injection, as previously described [20]. The tip of the catheter was advanced to the lumbar enlargement. A recovery period of 9 days after implantation was provided before proceeding to the behavioral study. Rats with neurological damage were excluded.
3. Drugs
Cisplatin (Tocris Cookson Ltd., Bristol, Avon, UK) was freshly dissolved in 0.4% dimethyl sulfoxide (DMSO) to a final concentration of 2 mg/mL. PD 149163 (Sigma-Aldrich, St. Louis, MO), a selective NTSR1 agonist, was dissolved in saline. SR 48692 (Tocris), a selective non-peptide NTSR1 antagonist, and dihydroergocristine (Tocris), a 5-HT receptor antagonist, were dissolved in 100% and 70% DMSO, respectively. The drugs were administered intrathecally in a 10-μL solution using a microinjection syringe pump. The control group (vehicle) was given DMSO of equal volume and concentration. Intraperitoneal injections were performed with a 26G needle attached to a syringe.
4. Assessment of mechanical allodynia
Mechanical allodynia was tested after the rats recovered from intrathecal surgery. The rats were placed on a wire net cage individually to evaluate paw withdrawal thresholds. The rats were acclimated to the environment for 30 minutes before testing. Mechanical allodynia was measured with von Frey filaments (Stoelting, Wood Dale, IL) to the hind paw using the up-and-down method. A series of eight von Frey filaments (0.4, 0.6, 1, 2, 4, 6, 8, 15 g) was gently applied perpendicular to the plantar surface of the hind paw for 5 seconds after the filament was bent. Brisk withdrawal or paw flinching was regarded as a positive response. The absence of a positive response to the pressure of the 15 g von Frey filament was regarded as the cut-off value. Only rats with definite allodynia were tested (mechanical withdrawal threshold < 5 g after CIPN was induced). Test compounds were randomly administered. The same volume of vehicle (saline or DMSO) was administered to the control group animals. All rats were evaluated only once, and the investigator was blinded to the treatment status of all animals in the experimental tests.
5. General behavior
Additional rats received the highest doses of the drugs to evaluate the behavioral changes after administration of each drug 1 hour after intrathecal or intraperitoneal delivery. Righting and place-stepping reflexes were observed to check motor functions, as described previously [21]. Pinna and corneal reflexes were judged as present or absent. Other abnormal behaviors, such as serpentine movement or tremors, were also evaluated.
6. Assessment of serotonin concentration in the spinal cord
The concentration of serotonin in the spinal cord was measured as described previously [22]. Tissue was obtained from the dorsal area of the lumbar (L4–5) spinal cord 14 days after the first injection of cisplatin. PD 149163 was administered intraperitoneally or intrathecally 2 hours before the harvest of spinal cord tissue. The serotonin concentration was detected using high-performance liquid chromatography and liquid chromatography-tandem mass spectrometry systems supplied by Shimadzu (LC 20A System and API 4000 Q Trap; Tokyo, Japan). A 0.4-mL aliquot of 0.5 M formic acid was added to homogenate the tissue.
7. Statistical analysis
Data are expressed as mean ± standard error. The dose-response data are presented as the percentage of maximum possible effect (%MPE): %MPE = [(post-drug threshold − post-injury baseline threshold)/(cutoff threshold − post-injury baseline threshold)] × 100. The dose-response data and serotonin concentrations were analyzed by one-way analysis of variance with Tukey’s post hoc test or the paired t-test. The statistical analysis was performed using IBM SPSS Statistics 22.0 software (IBM Corp., Armonk, NY). A P value < 0.05 was considered significant.
Go to :
RESULTS
A total of 171 rats were used in this experiment. Place-stepping and righting reflexes were observed at the highest dose used in this study, and normal, pinna and corneal reflexes were normally displayed as well. No overt abnormal behavioral changes were observed.
We initially investigated whether NTSR1 regulated the spinal nociceptive signal in the CIPN model. As shown in Fig. 1A and B, intrathecal administration of the selective NTSR1 agonist PD 149163 dose-dependently and markedly improved mechanical allodynia induced by cisplatin. In addition, the selective NTSR1 antagonist SR 48692, administered intrathecally, inhibited the antinociceptive effects induced by PD 149163 (Fig. 1C, D). These results indicate that NTSR1 plays a critical role in regulating CIPN-related pain at the spinal cord level. Next, we administered PD 149163 into the peritoneal space, a common route of systemic drug administration. As shown in Fig. 2A and B, intraperitoneal administration of PD 149163 increased the paw withdrawal threshold in a dose-dependent manner. However, intrathecally administered SR 48692 did not inhibit the antinociceptive effects induced by intraperitoneally administered PD 149163 (Fig. 2C, D).
 | Fig. 1Antinociceptive effect of intrathecal PD 149163 in chemotherapy-induced peripheral neuropathic rats. (A, B) The effect of intrathecal PD 149163 (PD: 0, 3, 10, and 30 ng) on the hind paw withdrawal threshold (PWT) to von Frey filaments. (C, D) The effect of intrathecal SR 48692 (SR: 0 or 100 µg) on the antiallodynic effect caused by PD 149163 (30 ng). SR 48692 was delivered 10 minutes before PD 149163 administration. Each line or bar represents mean ± standard error of 6-7 rats. MPE: maximum possible effect, i.t.: intrathecal administration. *P < 0.05 compared to vehicle. #P < 0.05 compared to PD 149163 (i.t.)-treated group.
|
 | Fig. 2Antinociceptive effect of intraperitoneal PD 149163 in chemotherapy-induced peripheral neuropathic rats. (A, B) The effect of intraperitoneal PD 149163 (PD: 0, 0.03, 0.1, and 0.3 mg/kg) for the hind paw withdrawal threshold (PWT) to von Frey filaments. (C, D) The effect of intrathecal SR 48692 (SR: 0 or 100 µg) on the antiallodynic effect caused by intraperitoneal PD 149163 (0.3 mg/kg). SR 48692 was delivered 10 minutes before PD 149163 administration. Each line or bar represents mean ± standard error from 6 rats. MPE: maximum possible effect, i.t.: intrathecal administration, i.p.: intraperitoneal administration. *P < 0.05 compared to vehicle.
|
Previous studies have shown that the antinociceptive effects induced by PD 149163 injected directly into the brain are inhibited by intrathecal administration of a non-selective serotonergic antagonist [9]. Therefore, we examined whether the effect of systemic administration of the selective brain-penetrating NTSR1 agonist PD 149163 in the CIPN model could be blocked by intrathecal administration of dihydroergocristine, a non-selective serotonergic antagonist. As shown in Fig. 3A and B, the antinociceptive effects of intraperitoneally administered PD 149163 were not inhibited by intrathecal administration of dihydroergocristine. However, when PD 149163 was administered intrathecally, the beneficial effect of PD 149163 on mechanical allodynia was diminished by intrathecally administered dihydroergocristine (Fig. 3C, D). Serotonin concentration at the spinal cord level decreased in CIPN-induced rats compared to the controls, but intrathecal or intraperitoneal administration of PD 149163 did not affect the observed decrease (Fig. 4).
 | Fig. 3Association between the serotonin (5-hydroxytryptamine) receptor and the antinociceptive effect of intraperitoneal or intrathecal administration of PD 149163. (A, B) The effect of intrathecal dihydroergocristine (DHE: 0 or 30 µg) on the antinociceptive effect of intraperitoneal PD 149163 (PD: 0.3 mg/kg). (C, D) The effect of intrathecal dihydroergocristine (30 µg) on the antinociceptive effect of intrathecal PD 149163 (30 ng). Dihydroergocristine was delivered 10 minutes before PD 149163 administration. Each line or bar represents mean ± standard error of 6-7 rats. PWT: paw withdrawal threshold, MPE: maximum possible effect, i.t.: intrathecal administration, i.p.: intraperitoneal administration. *P < 0.05 compared to vehicle. #P < 0.05 compared to PD 149163 (i.t.)-treated group.
|
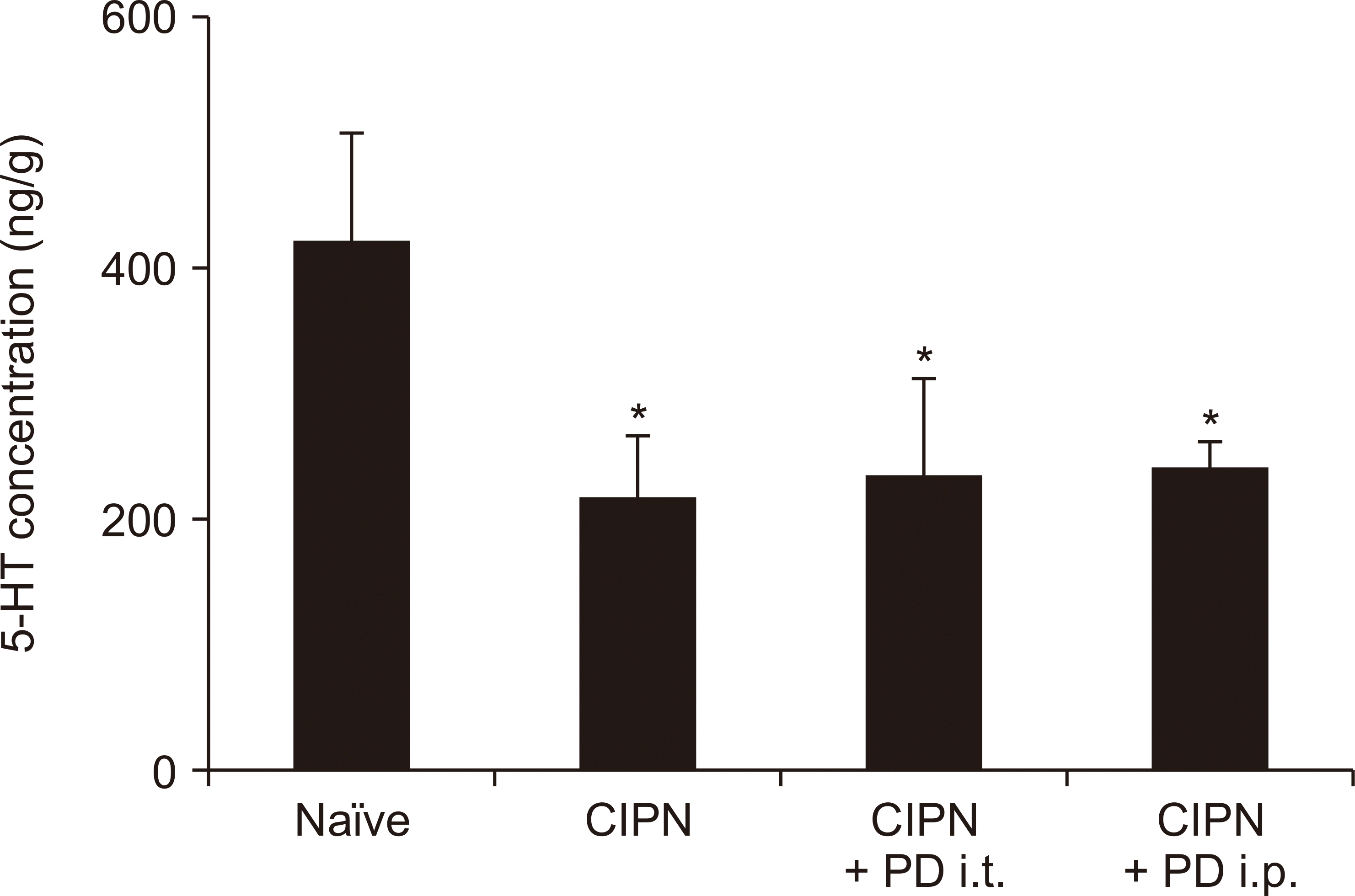 | Fig. 4The levels of serotonin (5-hydroxytryptamine, 5-HT) in the spinal cord of chemotherapy-induced peripheral neuropathic (CIPN) rats. The rats were divided into four groups: Naïve group, CIPN group, CIPN group treated with intrathecal PD 149163 (PD), CIPN group treated with intraperitoneal PD 149163. Each bar represents mean ± standard error of 6 rats. i.t.: intrathecal administration, i.p.: intraperitoneal administration. *P < 0.05 compared to naïve rats.
|
Go to :
DISCUSSION
The present study demonstrated potent anti-allodynic effects of intrathecal and intraperitoneal NTSR1 agonist in CIPN, and provided evidence that the NTSR1 receptor plays significant roles in spinal- and supraspinal-level pain processing. The anti-allodynic effect of intrathecal PD 149163 was reduced by intrathecal pretreatment with the NTSR1 antagonist SR 48692, which parallels the analgesic effect shown in previous studies using the formalin test and sciatic nerve constriction model [12,14]. In addition, these previous studies showed that NTSR1 is distributed in the dorsal root ganglia and the superficial layers of the dorsal horn at the spinal cord [12]. We also examined the effect of the intrathecal NTSR2 agonist levocabastine in the CIPN model, but it did not affect mechanical allodynia intensity (data not shown). Moreover, a selective agonist of NSTR3 has not been developed, so it could not be added to this experiment. Therefore, these results indicate that NTSR1, not NTSR2, is involved in the regulation of the chemotherapy-evoked nociceptive transmission in the spinal cord. However, previous studies showed that intrathecal NTSR2 agonists, including JMV431, levocabastine, or β-lactotensin, produced analgesic effects in sciatic nerve injury- and formalin-induced pain models [23,24]. Although such discrepancy on the effect of NTSR2 agonist could not be exactly understood, it may be due to the kinds of drug and nociception. Thus, further research is necessary in future.
NTS has little analgesic effect when administered peripherally because it is readily degraded by peptidases in the periphery, and does not easily pass the blood-brain barrier [25,26]. However, PD 149163 is a stable and brain penetrating NTSR1 agonist [27]. As shown in our experimental data, intraperitoneal administration of PD 149163 effectively abolished mechanical allodynia. However, intrathecal SR 48692 had no influence on the effect of intraperitoneal PD 149163. These results suggest that intraperitoneal PD 149163 mainly acts on brain NTSR1 and its supraspinal analgesic action was sufficient to suppress neuropathic pain in the CIPN model. NTS is widely distributed in the CNS, and acts as a neurotransmitter in the brain. For example, systemic administration of NTSR1 agonists regulate various CNS disorders, including schizophrenia [28]. In addition, direct administration of NTS into specific brain regions (periaqueductal gray region and RVM) exerted strong analgesic effects [29,30].
Previous studies have demonstrated NTSR1 regulation of serotonergic neuronal activity and 5-HT release [11,17]. NTS increases 5-HT release in brain frontal cortex slices, which was inhibited by NTSR1 antagonists [17]. Local injection of NTS in the prefrontal cortex dose-dependently increased 5-HT release in freely moving rats, which was also inhibited by SR 48692 [11]. It is possible that supraspinal NTSR1 is involved in descending serotonergic pain control. Indeed, another study found that direct injection of NTS or PD 149163 in the RVM produced an antinociceptive effect in the thermal tail flick test. Intrathecal administration of a non-selective serotonergic antagonist blocked this effect, which suggests that increased 5-HT release from spinally projecting serotonergic neurons in the RVM is responsible for the antinociceptive effect of RVM NTSR1 agonists [9].
However, in our study using the CIPN model, the anti-allodynic effect of intraperitoneal PD 149163 was not affected by pretreatment with intrathecal dihydroergocristine, a non-selective serotonergic antagonist. This could stem from different pain modalities: acute thermal nociception in naïve animals versus mechanical allodynia in the neuropathic CIPN model. Compared with naïve animals, the CIPN animals showed a significant decrease in spinal cord 5-HT content. Thus, activation of NTSR1 in the brain did not increase 5-HT in CIPN rats to the level of naïve animals. In addition, intraperitoneal PD 149163 had no effect on spinal cord serotonin levels in our CIPN model. Together, these results provide evidence for pathologic changes in the serotonergic descending inhibitory pathway in CIPN. Another possibility is that the intraperitoneal NTSR1 agonist may activate the noradrenergic descending inhibitory pathway. Support for this possibility stems from recent studies demonstrating that local injection of NTSR1 agonist to the brain partially inhibited the thermal antinociceptive effect via the intrathecal α2-adrenoceptor antagonist [10].
In contrast to the anti-allodynic effect of the intraperitoneal NTSR1 agonist, which was independent of spinal level serotonergic action, intrathecal administration of the NTSR1 agonist diminished the effect of the intrathecal nonspecific 5-HT antagonist. It is possible that the intrathecal 5-HT antagonist further suppressed the spinal serotonergic modulating system responsible for reduction of spinal 5-HT content. These data suggest that the basal activity of the serotonergic receptor is required for the NTSR1 agonist to exhibit a maximal antinociceptive effect in the spinal cord. However, NTSR1 agonist may not directly affect the synthesis of serotonin.
Collectively, our study demonstrates that PD 149163 has a strong anti-allodynic effect in a rodent model of CIPN at the supraspinal as well as the spinal level and provides new therapeutic targets for effective CIPN analgesics. In particular, the effect of PD 149163 may be indirectly related to the serotonergic pathway.
Go to :
 XML Download
XML Download