

 PDF
PDF Citation
Citation Print
Print


Introduction
The placenta is an essential organ that is formed transiently during mammalian pregnancy. It enables the transport of nutrients, gases, and wastes between the mother and the developing fetus (1). In order to maintain a healthy pregnancy, the placenta must properly develop into the appropriate trophoblast lineages (2). Abnormal placental development is associated with a reduction in placental function and pregnancy-associated disorders, such as fetal growth restriction (FGR), placental insufficiency and/or preeclampsia (3).
Both human and rodent studies have investigated the underlying pathways that regulate placental differentiation and development. TGF-β signaling has been implicated as a possible regulator of placental development and dysfunction, as all three TGF-β isoforms have been shown to be elevated in human preeclamptic pregnancies (4–6). TGF-β has been shown to regulate a large number of critical cellular actions such as cell growth, differentiation, lineage determination, extracellular matrix restructuring, and apoptotic cell death (7).
TGF-β signals through type I and type II receptors that heterodimerize and phosphorylate Smad proteins (8). Loss of the primary type I receptor for TGF-β, Activin-receptor Like Kinase 5 (ALK5 or TβRI), is embryonically lethal and the loss of type II receptor, TGF-βRII, is perinatally lethal (9, 10). TGF-β can signal through phosphorylation of either of two receptor-regulated Smads (R-Smads), Smad2 or Smad3, which can then dimerize with Smad4 allowing for nuclear translocation and binding to response elements that activate downstream signaling pathways (11). Knockout studies have shown that both Smad2 and Smad4 are embryonically lethal, whereas, ablation of Smad3 results in viable offspring, indicating some non-overlapping function of the regulatory Smads (12). In addition to R-Smads, inhibitory Smads (I-Smads) can also affect the signal transduction pathway induced by TGF-β. Smad7 is upregulated by TGF-β and has the ability to inhibit signaling of the TGF-β family through a negative feedback loop (13, 14). Studies in mice have found that a Smad7 deficiency can also result in strain-dependent defects in mice ranging from decreased adult size to perinatal death, indicating that the entire TGF-β signal transduction pathway is essential to proper development and embryonic survival (13).
As the rodent placenta has numerous structures and functions that are comparatively similar to the human placenta, mouse models have been useful in establishing the importance of TGF-β and Smad proteins in placental development (12, 15). Placental labyrinthine cells are closest to the fetus and are important for the transport of nutrients and wastes between the mother and developing fetus (16, 17). SM10 cells are a mouse labyrinthine progenitor cell line, which have been previously shown to differentiate after treatment with TGF-β and can be used as a model for the transport layer of the placenta (18–21). The pathway activated by TGF-β to induce SM10 progenitor differentiation, however, has yet to be fully investigated. Our previous studies have shown that TGF-β induced down regulation of the stem cell regulator, Id2, is necessary for SM10 trophoblast progenitor differentiation (20). In this study, we show that all three TGF-β isoforms have the ability to terminally differentiate SM10 cells, whereas other members of the TGF-β superfamily, i.e. Activin A and Nodal, do not. Additionally, we have identified that TGF-β signaling in SM10 placental labyrinthine cells is primarily induced via Smad2 phosphorylation and transactivation of the Activin response element, in contrast to the more classical activation of 3TP and SBE promoter response elements, to alter gene expression necessary to induce differentiation.
Go to : 
Materials and Methods
Materials
RPMI1640/L-glutamine, DMEM/High glucose, 2β-mercaptoethanol (35602), antibiotic-antimycotic (SV30079.01) and Supersignal West Pico Chemiluminescence Substrate (34080) were purchased from Thermo Scientific. Sodium pyruvate (25-000-CI) was purchased from Mediatech Inc. Fetal Bovine Serum (FBS) (S01520) was purchased from Biowest. Metafectene transfection reagent (T020) was purchased from Biontex Laboratories (Germany). 3[H]-2-deoxyglucose was purchased from MP Biochemicals. TGF-β1 (240-B), TGF-β3 (243-B3), Nodal (1315-ND-025CF) and Activin A (338-AC) were obtained from R&D Systems. TGF-β2 was kindly provided by Dr. Steven Ledbetter (Genzyme). SB 431542 (#1614) was purchased from Tocris. The 3TP-lux reporter has been previously described (22, 23). Anti-Smad2 rabbit polyclonal (51-1300), rabbit polyclonal anti-Smad3 (51-1500), were purchased from Zymed. Rabbit polyclonal anti-phosphoSmad3 was a kind gift of Dr. Ed Leof, Mayo Clinic (24). Mouse monoclonal anti-Smad4 (B8)(sc-7966) and rabbit polyclonal anti-phosphoSmad2 (06-829) were purchased from Santa Cruz Biotechnology and Upstate Biotechnology, respectively. The secondary antibodies anti-rabbit IgG (W401B) and anti-mouse IgG (W402B) were purchased from Promega. The pcDNA3 vector was obtained from Invitrogen.
Cell culture and trophoblast differentiation
Mouse SM10 cells were maintained in RPMI1640/L-glutamine medium supplemented with 1mM sodium pyruvate, 50 μM 2β-mercaptoethanol, 1% antibiotic-antimycotic, and 10% FBS (18–21). SM10 cells were treated with TGF-β1 (5 ng/ml), TGF-β2 (5 ng/ml), TGF-β3 (2 ng/ml), Activin A (5 ng/ml), Nodal (250 μg/ml), or vehicle (4mM HCl, 0.5% mg/ml BSA) for 72 hrs to induce differentiation. COS-7 cells were obtained from ATCC (CRL-1651) and were cultured in DMEM/High glucose, 10% heat inactivated fetal bovine serum, and 1% antibiotic-antimycotic. All cell types were passaged at 80~90% confluence.
Receptor inhibition
Inhibition of Alk receptors in SM10 cells was conducted by plating 3×105 SM10 cells in p60s. Cells were then treated with or without the ALK-5 (TGF-β receptor) specific inhibitor SB431542 (10 μM) for 1 hour, followed by treatment without or with TGF-β2 (5 ng/ml) for 48 hours.
Growth inhibition
SM10 cells were plated at 1×105 cells/ml in 24 well plates for 24 h prior to treatment. Cells were then treated with TGF-β2 or Activin A for 72 hours and subsequently incubated with 1 μCi/ml 3H-Thymidine (ICN) per well for 4 h. The cells were washed twice with 10 per cent tri-chloroacetic acid (TCA) for 10 min and lysed by adding 300 μl of 0.2 N NaOH containing 40 μg/ml of salmon sperm DNA. Cell lysates (100 μl) were counted in Scintiverse scintillation fluid (8).
Western blotting
Western blotting was performed as previously described (20, 25) using rabbit polyclonal anti-Smad 2, rabbit polyclonal anti-Smad 3, mouse monoclonal anti-Smad4 (B8), rabbit polyclonal anti-phosphoSmad2, rabbit polyclonal anti-phosphoSmad3, or mouse monoclonal anti pan-actin (C4) antibodies at 1/500 overnight at 4°C overnight in blocking buffer (5% [w/v] non-fat milk, pH 7.4 in 1× PBS containing 0.05% Tween-20 [1× PBST]). Following overnight incubation with primary antibody, the membrane was washed with 1xPBST, probed with HRP-conjugated anti-rabbit IgG or anti-mouse secondary antibody at 1/10,000 for 1 hour at RT, and visualized using Enhanced Chemi-Lumeniscence reagent according to manufacturer’s instructions.
Luciferase reporter transactivation
SM10 cells were plated at 2×105 cells/well and were transfected the next day with 0.2 μg pRLSV40 and either 1 μg 3TP-lux; 1 μg Glut1-lux; or 1 μg ARE-lux and 0.8 μg hFAST1 for 24 hours. 1 μg pCDNA3 or 1 μg Smad2 DNA were also transfected in designated assays using Metafectene transfection reagent, as previously described (26). The cells were placed in fresh media and vehicle or 5 ng/ml Activin A, or 5 ng/ml TGF-β was added for 72 hrs, after which the cells were rinsed in 1XPBS, and analyzed using the Dual Luciferase Reporter system according to manufacturer’s directions (27). All ARE-Luc values were normalized to pRLSV40 values.
Statistical analysis
All experiments were conducted a minimum of three times independently, with similar results. Quantitative data is represented by average±Standard Error of the Mean (SEM). Data was analyzed via GraphPad Prism 7.0. Statistical analysis for the Glut1-lux assay (Fig. 1E) and growth inhibition (Fig. 2E) was conducted by using one-way ANOVA followed by Dunnett’s multiple comparisons test. No statistical analysis was required for Fig. 3. Statistical analysis for 3TP-lux assays were conducted by Student t-test (Fig. 4A). Statistical analysis for ARE-lux assays were conducted by two-way ANOVA with Tukey’s post hoc test for Smad7 overexpression (Fig. 4F) and Sidak’s multiple comparisons test for Smad 2 overexpression (Fig. 4E). P values for 95% confidence interval were calculated. p<0.05 was considered significant and is denoted by * or θ.
 | Fig. 1TGF-β isoforms induce trophoblast progenitor differentiation and Glut-1 transactivation. SM10 cells were treated for 72 hrs with (A) vehicle control, (B) 5 ng/ml TGF-β1, (C) 5 ng/ml TGF-β2, or (D) 2 ng/ml TGF-β3 to induce differentiation. (E) Glut-1 luciferase reporter activity in SM10s cells treated with vehicle control, TGF-β1, TGF-β2, or TGF-β3. Cells were transiently transfected with Glut1-lux and pRLSV40 using Metafectene. Twenty-four hours post-transfection, cells were treated with vehicle control, TGF-β1 (5 ng/ml), TGF-β2 (5 ng/ml), or TGF-β3 (2 ng/ml) for 24 or 72 hours. Luciferase activity was analyzed using the dual luciferase assay system. The Glut1-lux transactivation values were normalized to the constitutively active reporter, pRLSV40. Error bars represent standard error of the mean. (θ) indicates a significant increase in Glut1 luciferase transactivation compared to vehicle control at 24 hours and (*) indicates a significant increase from control at 72 hours. p<0.05.
|
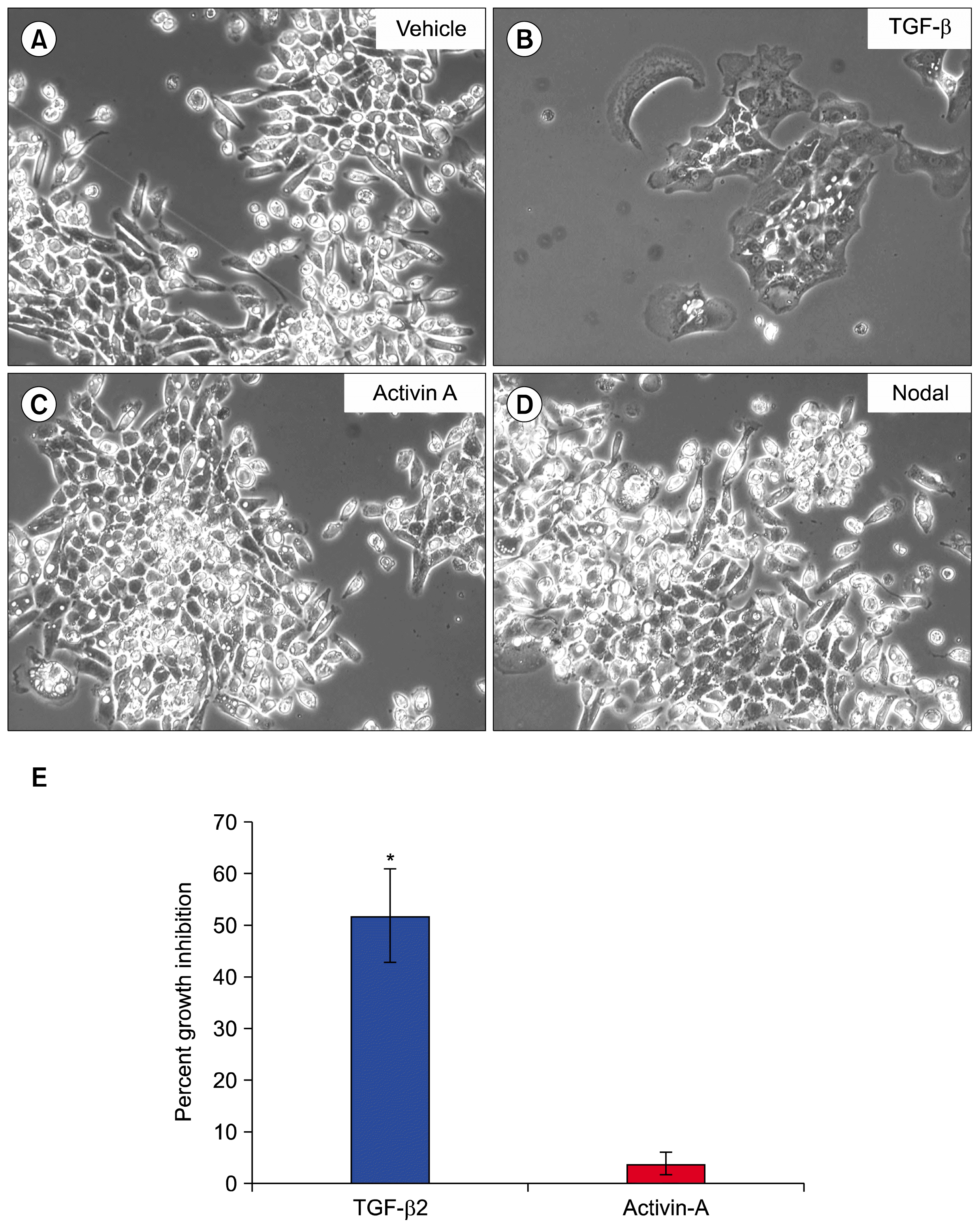 | Fig. 2TGF-β induces SM10 cell differentiation and growth inhibition. SM10 cells were treated with (A) vehicle control, (B) TGF-β2 (5 ng/ml), (C) Activin A (5 ng/ml), or (D) Nodal (250 μg/ml) for 72 hrs to induce differentiation. (E) After 72 hours of treatment, cells were incubated with 1 μCi/ml 3H-Thymidine per well for 4 hours. Cells were then lysed and counted in scintillation fluid. Percent growth inhibition was determined as the amount of 3H-Thymidine incorporation compared to control. Percent growth inhibition is normalized to vehicle treated cells. Error bars represent standard error of the mean. *p<0.001.
|
 | Fig. 3TGF-β induced differentiation can be mediated through the Alk-5 receptor. SM10 cells were treated without (A, B) or with (C, D) the Alk inhibitor SB431542 (10 μM) for 1 hour, followed by treatment without (A, C) or with (B, D) TGF-β 2 (5 ng/ml) for 48 hours.
|
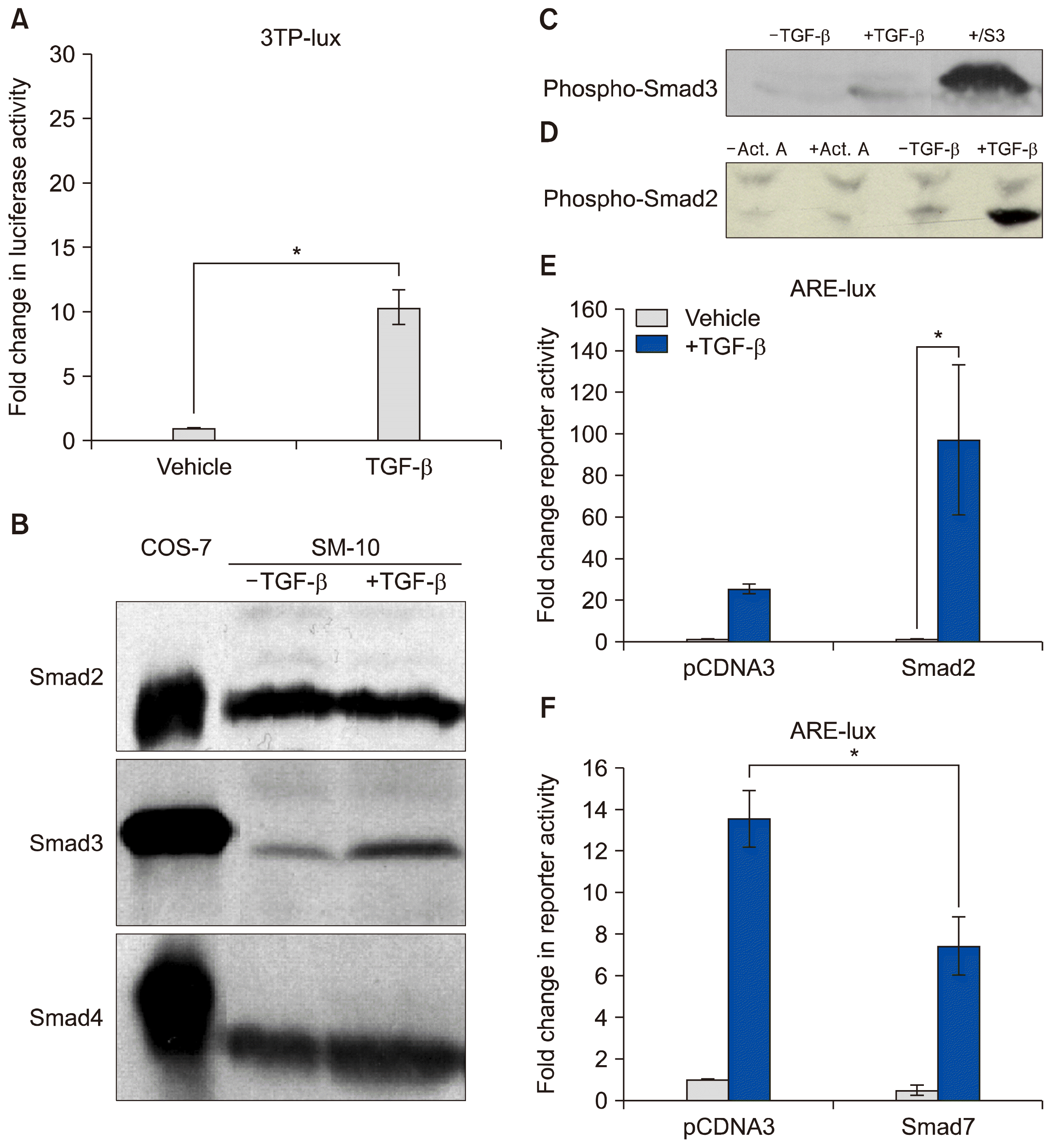 | Fig. 4TGF-β triggers Smad2 phosphorylation and transactivation. (A) 3TP luciferase reporter activity in SM10s cells treated with vehicle control or TGF-β2. Cells were transiently transfected with 3TP-lux and pRLSV40 using Metafectene. Twenty-four hours post-transfection, cells were treated with vehicle or TGF-β2 (5 ng/ml) for 72 hours. Luciferase activity was analyzed using the dual luciferase assay system. The 3TP-lux transactivation values were normalized to the constitutively active reporter (pRLSV40). (B) Western blotting analysis of Smad2, Smad3, and Smad4 in TGF-β2 or vehicle-treated SM10 cells. COS-7 cells were used as a positive control. (C) Western blotting analysis of Phospho-Smad3 in TGF-β2 or vehicle-treated SM10 cells. COS-7 cells transiently transfected with the plasmid, pXIX-Smad3 and treated with TGF-β were used as a positive control. (D) Western blotting analysis of Phospho-Smad2 in the TGF-β2, Activin A, or vehicle-treated SM10 cells. (E) ARE luciferase reporter activity in SM10 cells transiently transfected with pCDNA3 control or pRK5-Smad2 and treated with vehicle control or TGF-β2. Cells were transiently transfected with ARE-lux, pLv-CMV-hFAST-1, and pRLSV40 using Metafectene. Twenty-four hours post-transfection, cells were treated with vehicle or TGF-β2 (5 ng/ml) for 72 hours. Luciferase activity was analyzed using the dual luciferase assay system. The ARE-lux transactivation values were normalized to the constitutively active reporter (pRLSV40). (F) ARE luciferase reporter activity in SM10 cells transiently transfected with pCDNA3 control or pRK5-Smad7 and treated with vehicle control or TGF-β2. Cells were transiently transfected with ARE-lux, pLv-CMV-hFAST-1, and pRLSV40 using Metafectene. Twenty-four hours post-transfection, cells were treated with vehicle or TGF-β2 (5 ng/ml) for 72 hours. Luciferase activity was analyzed using the dual luciferase assay system. The ARE-lux transactivation values were normalized to the constitutively active reporter (pRLSV40). Error bars represent standard error of the mean. *p<0.05.
|
Go to :
Results
TGF-β isoforms differentiate SM10 progenitor cells
Previous studies have shown that transforming growth factor beta (TGF-β), and in particular TGF-β2, has the ability to cause SM10 placental progenitor cells to differentiate into cells with labyrinthine trophoblast characteristics (19–21). In order to further understand the signal transduction mechanisms involved in placental labyrinthine cell differentiation, SM10 cells were treated with different TGF-β isoforms (β1, β2, or β3) and cell morphology was examined by light microscopy. Vehicle-treated SM10 cells remained undifferentiated (Fig. 1A). TGF-β1, TGF-β2, and TGF-β3 isoforms all induced similar morphologic trophoblast differentiation, characterized by cell aggregation and colony formation, characteristic of the labyrinthine lineage, as we have previously reported (Fig. 1B~D) (19–21).
In order to further investigate the ability of each TGF-β isoform to transcriptionally induce downstream targets in SM10 cells, luciferase assays were performed at 24 and 72 hours post TGF-β treatment to assess the transactivation of glucose transporter 1, Glut-1. Previous studies have shown that TGF-β has the ability to stimulate Glut-1 signaling (28, 29). In the current study, all three TGF-β isoforms also exhibited similar rates of Glut-1 transcriptional activation after 24 hrs of treatment; however, TGF-β2 exhibited the greatest activation at 72 hrs and therefore was used to differentiate SM10 cells for the remaining experiments (Fig. 1E).
TGF-β, but not Activin A or Nodal, promote SM10 progenitor differentiation
Since TGF-β isoforms were capable of inducing SM10 cell progenitor differentiation, we investigated whether other predominant members of the TGF-β superfamily, Activin A and Nodal, could promote differentiation. Vehicle treated cells did not differentiate, whereas cells treated with TGF-β were differentiated as expected, as indicated by characteristic cell aggregation (Fig. 2A, B). Treatment with Activin A or Nodal was did not differentiate SM10 progenitors (Fig. 2C, D). As SM10 cells differentiate, their rate of proliferation decreases. Therefore, to assess the ability of Activin A and TGF-β to inhibit cell proliferation, growth inhibition studies were conducted using 3H-thymidine. TGF-β treatment induced 52% growth inhibition of SM10 cells, while Activin A had no effect on proliferative activity of SM10 cell progenitors (Fig. 2E).
TGF-β induced differentiation is dependent upon TGFBR1 signaling
Further investigation of the signaling pathways governing TGF-β dependent differentiation of SM10 progenitors were conducted by treating cells with SB431542 in the presence or absence TGF-β. SB431542 inhibits Activin Receptor-Like Kinases (ALKs): ACVR1B (ALK-4), TGFBR1 (ALK-5), and ACVR1C (ALK-7) (30). TGF-β is known to signal through ALK-5, whereas Activin A signals through ALK-4, and Nodal via ALK-7 (31, 32). SM10 cells treated with vehicle or SB431542 alone did not differentiate (Fig. 3A, B), whereas cells treated with TGF-β differentiated as expected (Fig. 3C). SM10 cells treated with both SB431542 and TGF-β, however, were unable to fully differentiate as indicated by the lack of colony formation (Fig. 3D). These data suggest that TGF-β -induced SM10 differentiation can be mediated via ALK-5, as TGF-β has not been reported to signal through either ALK-4 or ALK-7 (32).
TGF-β induces phosphorylation of Smad2 but not Smad3 in SM10 cells
In order to examine the signaling pathways transcriptionally activated by TGF-β induced differentiation in SM10 progenitor cells, luciferase assays with 3TP-lux reporter were conducted. 3TP-lux is a luciferase construct containing a luciferase reporter gene. It has three copies of the TPA response element (3TP) from the human collagenase gene and one TGF-β response element from the human plasminogen activator inhibitor 1 (PAI-1) gene, which can be transcriptionally activated by TGF-β (23). TGF-β treatment of SM10 cells induced 3TP luciferase activity by approximately 10-fold (Fig. 4A).
Previous studies have shown that TGF-β can signal through either Smad2 or Smad3 phosphorylation to induce cellular signaling. Both Smad2 and Smad3 are regulatory (R-Smads) that, once phosphorylated, can bind to Smad4, create a heterodimer, and translocate to the nucleus to activate gene transcription (33). In order to determine which R-Smads are activated by TGF-β-induced differentiation, SM10 cells were analyzed by Western blot for the presence of Smad2, Smad3, and Smad4 protein after TGF-β or vehicle treatment (Fig. 4B). COS-7 cells were used as a positive control (Fig. 4B). Smad2, Smad3, and Smad4 were all present in undifferentiated SM10 progenitor cells. Additionally, TGF-β treatment did not increase the levels of Smad2 and Smad4; however, TGF-β treatment resulted in slightly increased levels of Smad3 protein.
While increased protein levels can often be helpful in understanding signaling transduction pathways, the activation of TGF-β receptor signaling is propagated via phosphorylation of the R-Smads to ultimately allow for binding of Smad4 and transcriptional activation. To examine phosphorylation levels of Smad2 and Smad3; SM10 cells were treated with TGF-β for 72 hours and analyzed by Western blot using phospho-specific antibodies (34). Phosphorylated Smad3 (phospho-Smad3) was weakly detected and present at very low levels in SM10 cells regardless of TGF-β treatment (Fig. 4C). In contrast, phospho-Smad2 was substantially increased upon TGF-β administration (Fig. 4D). These results clearly indicate that Smad2, but not Smad3, activation is induced by TGF-β in SM10 cells during the process of differentiation. Activin A was not able to induce Smad2 phosphorylation in SM10 cells (Fig. 4D), consistent with its inability to induce SM10 progenitor differentiation.
TGF-β induces transactivation of Activin Responsive Element via Smad2
To investigate TGF-β signaling via phospho-Smad2, we examined 3TP and ARE (Activin Response Element) luciferase reporter assays, which have been shown to be activated by TGF-β (22, 35). Smad binding element reporter activity (SBE-lux) was not assessed due to the known preferential specificity for phospho-Smad3 and the lack of Smad3 activation in SM10 cells (Fig. 4C) (36). ARE activity induced by TGF- β or Activin A was assessed using luciferase reporter assays using an ARE-lux construct in the presence of forkhead activin signal transducer-1 (FAST-1), as it is required for induction of the activin response element (35). We determined if ARE transactivation could be stimulated in SM10 cells by overexpressing Smad2 by transient transfection. SM10 cells transfected with control DNA (pCDNA3) and treated TGF-β had an increased level of luciferase activity compared to SM10 cells treated with vehicle (Fig. 4E). Additionally, SM10 cells transiently transfected with a Smad2 and subsequently treated with TGF-β had a significant increase in ARE-lux activation compared to SM10 cells overexpressing Smad2 and treated with vehicle (Fig. 4E). Therefore, TGF-β treatment induces luciferase activity of the Activin response element in SM10 cells, and triggers the expression of genes involved in the regulation of cell growth and differentiation.
Smad7 reduces activation of the activin responsive element
To further investigate TGF-β mediated transactivation of the Activin response element, SM10 cells were transiently transfected with ARE-lux, FAST1, and Smad7 and treated with TGF-β. Smad7 is an inhibitor of TGF-β signaling that works by preventing R-Smad activation (37). Smad7 is upregulated by TGF-β, and therefore, can act as part of a negative feedback loop (37). SM10 cells treated with TGF-β increased ARE-lux transactivation in cells transfected with control DNA (pCDNA3) compared to vehicle treated cells (Fig. 4F). Overexpression of Smad7 via transient transfection in SM10 cells significantly inhibited TGF-β induced transactivation of ARE-lux (Fig. 4F).
Go to :
Discussion
Our previous studies have demonstrated the importance of stem cell regulator, Id2, in the regulation of trophoblast progenitor differentiation and have shown that Id2 downregulation is necessary for TGF-β induced differentiation in SM10 cells (20). As proper trophoblast differentiation of labyrinthine cells is critical for establishment of feto-maternal placental nutrient transport and the regulation of cell metabolism, we investigated the signaling mechanisms involved in TGF-β induced differentiation of SM10 cells. Our data indicate that all three TGF-β isoforms were able to induce trophoblast progenitor cell differentiation; while other members of the TGF-β superfamily, Nodal and Activin A, were not. Previous work in trophoblast stem (TS) cells showed that Actvin A prolonged proliferation and promoted differentiation along the labyrinth lineage; whereas TGF-β promoted labyrinth differentiation, but did not prolong proliferation (38). The inability of Activin A to differentiate SM10 cells may be, in part, due to the more committed nature of these progenitor cells, compared to TS cells, which are capable of differentiating into all trophoblast lineages. In addition, our data indicate that TGF-β induced differentiation can be mediated through the ALK5 receptor with subsequent phosphorylation of Smad2. Our data further indicate that the TGF-β/phospho-Smad2 signal is mediated via the Activin Response Element (ARE) in order to regulate downstream targets involved in cell growth and trophoblast differentiation in response to TGF-β in labyrinthine trophoblast cells. ARE activation in the presence of TGF-β can be inhibited by upregulation of Smad7, which is known to inhibit the activation of Smad2 and Smad3. In summary, our studies identify an important regulatory signal transduction pathway in SM10 cells that serves as a model of labyrinthine trophoblast progenitor differentiation (Fig. 5).
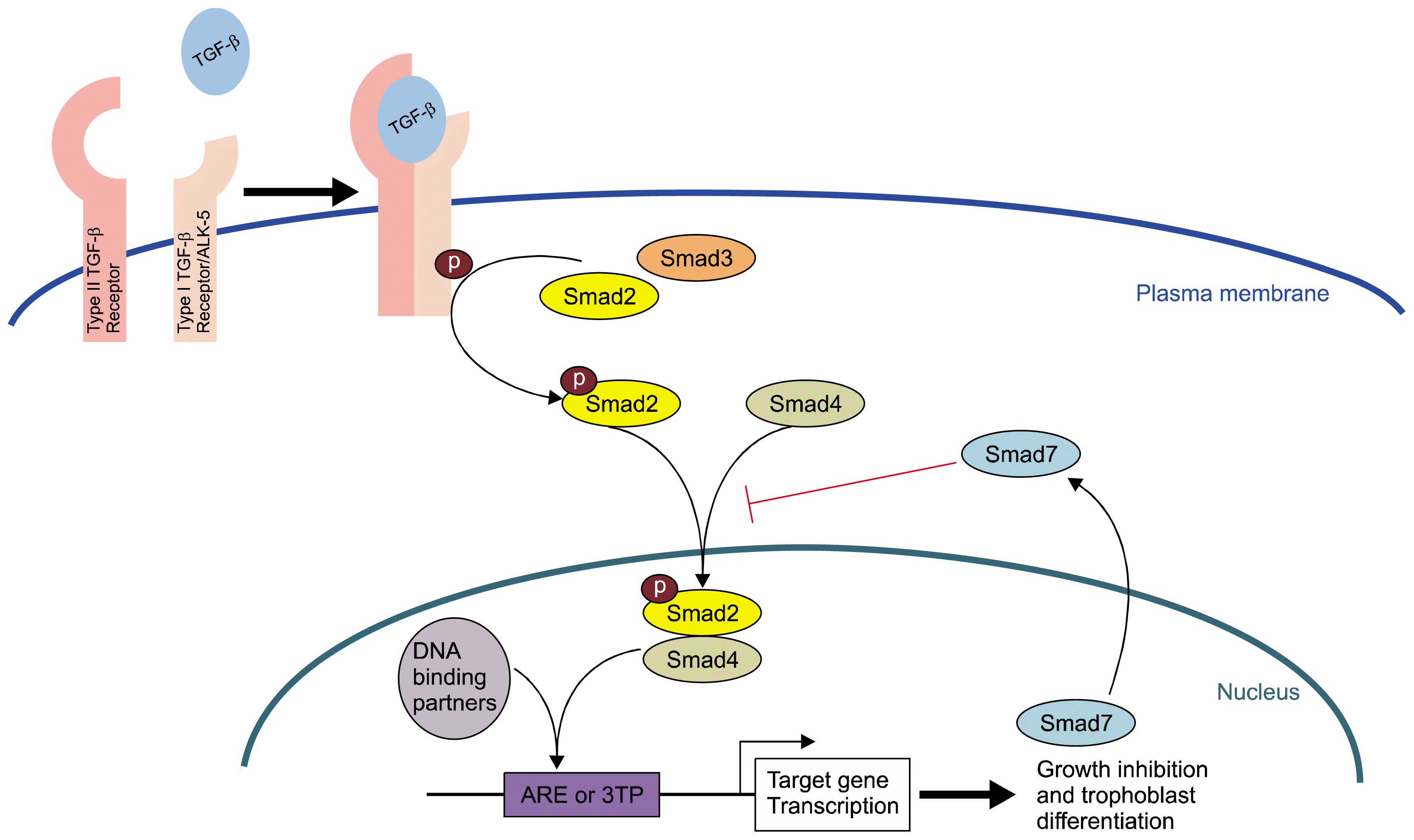 | Fig. 5TGF-β induces Smad2 Phosphorylation, ARE Induction, and Trophoblast Differentiation Diagrammatic representation of the TGF-β signaling pathway in SM10 growth inhibition and trophoblast differentiation. The ligand, TGF-β, binds to cognate receptors and triggers activation via the TGF-β Type I receptor (ALK-5) to induce the phosphorylation of Smad2. Phospho-Smad2, via classically presumptive Smad 4 binding, triggers the transactivation of ARE response elements in target genes that inhibit cell growth and promote labyrinthine trophoblast differentiation. Smad 7 acts as a classical negative feedback inhibitor for the TGF-β signaling pathway.
|
Go to :
 XML Download
XML Download