

 PDF
PDF Citation
Citation Print
Print


Introduction
Hepatitis C virus (HCV) infection is a major leading cause of chronic liver disease (CLD) worldwide (1). Egypt (with the genotype IV as the most common) has the highest prevalence of HCV in the world, estimated nationally at 14.7% (2). The consequences of chronic HCV infection represent compelling health problems and is the most frequent cause of viral-related cirrhosis and liver cancer and the principal indication for liver transplantation worldwide (3). HCV causes persistent infection in ~75–80% of patients. In these individuals, the function of HCV-specific immune cells is impaired by ligation of inhibitory receptors, the repertoire of which has expanded considerably in the past few years (4). B7-like molecules and their cognate receptors constitute important co-stimulatory pathways that control and fine-tune immune responses. In recent years, an array of new members of the B7 family has been identified, including B7-H1 (PD-L1) and B7-DC (PD-L2) (5). Both are ligands for programmed death-1 (PD-1), which is expressed on activated T and B cells (6–8). PD-L1 expression has been described to be induced in a variety of organs (5, 9). An overwhelming number of studies supported the role of PD-L1 as a negative regulator of T cell responses and suggest that PD-L1 promotes peripheral tolerance (10). Recently, hepatic accumulation and impaired apoptosis in CD8+ T cells have been observed in experimental autoimmune hepatitis in PD-L1-deficient mice leading to accelerated damage of hepatocytes (11). Golden-Mason et al. (12) and Radziewicz et al. (13) showed that PD-1/PD-L1 pathway was critical in persistence of HCV infection and represented a potential novel target for reversible immune dysfunction.
Acute and chronic liver injuries with different forms of cellular damage induce recruitment of stem cells from the bone marrow (BM) and their involvement in liver regeneration (14). They represent the third progeny, after hepatocytes and hepatic progenitor cells that contribute to liver repair. PD-L1 (B7-H1) was reported recently in non-parenchymal liver cells (15, 16). mesenchymal stem cells (17) and cultured bone marrow–derived mast cells (18). Here we analyzed the expression of PD-L1 on BM-derived HSCs and provide data suggesting novel bidirectional interaction with immune cells and their possible correlation with T-cell apoptosis in chronic HCV infected patients.
Go to : 
Materials and Methods
Study participants
50 subjects were recruited and divided into two groups: chronic HCV-infected patients (n=30), and healthy controls (n=20). All patients were prospectively recruited from the outpatient clinic of the Liver Hospital, Mansoura, Egypt, during the period from December 2016 to March 2017. All HCV-infected patients were negative for other chronic liver diseases including viral hepatitis A and B, which were defined by seronegativity with enzyme immunoassays. Patients with a history of habitual alcohol consumption or hepatocellular carcinoma, previous interferon treatment, decompensated liver disease (ascites, jaundice, variceal hemorrhage, or encephalopathy) and liver transplantation were excluded from the study. HCV infection was confirmed by detectable plasma HCV viral load measured by PCR assay using (COBAS Ampliprep/COBAS TaqMan; Roche Diagnostics AQ5, USA) for HCV-RNA. Healthy donors were negative for HCV, HBV and HIV-1 infection. Our study was approved by the local ethics committee and all patients provided written informed consent before their enrollment in the study.
Clinical biochemical tests
Liver associated enzymes including ALT, AST and other variables such as albumin, bilirubin, platelets and WBCs count were measured by clinical standardized methods. Full baseline characteristics and virological data of enrolled subjects are summarized in Table 1.
Table 1
Clinical characteristics of the study population
| Parameters | CHC (n=30) | Control (n=20) | p value |
|---|---|---|---|
| Male/Female | 18/12 | 13/7 | |
| Age (years) | 44.8±13.7 | 45.15±12.1 | 0.935 |
| ALT (IU/L) | 50.1±17.6 | 18.3±5.1 | <0.0001* |
| Total bilirubin (mg/dl) | 2.8±0.98 | 0.6±0.22 | <0.0001* |
| Albumin (g/dl) | 3.36±0.7 | 4.1±0.59 | <0.0001* |
| WBC count (×103/μl) | 7.9±1.7 | 6.8±2.1 | 0.445 |
| Platelet count (×109/L) | 241.3±59.2 | 275.9±56.2 | 0.191 |
| HCV genotyping (4a/4c) | 16/14 | ND | |
| HCV RNA (log10IU/ml) | 5.7±1.3 | ND | |
| Prior medication for HCV | 0 | 0 |
![]()
Flow cytometric analysis of circulating CD34+ HSCs
Blood samples were obtained in the outpatient department for the study groups and also for the healthy control subjects. Ten milliliters of blood was drawn from the antecubital vein. Mononuclear cells (MNCs) were then isolated by density-gradient centrifugation of Ficoll 400 (Ficoll® Paque Plus, GE17-1440-02, Sigma-Aldrich, Germany) as previously described (19). Then CD34+ cells were isolated by using CD34+ MiniMacs high-gradient magnetic separation column (Miltenyi Biotec, Bergisch Gladbach, Germany) following manufacturer’s instruction. The isolated CD34+ cells were then analyzed by conventional dual color immunofluorescence using PE-conjugated anti-CD34, vioblue-conjugated anti-CD45 antibodies (Miltenyi Biotec, Germany). Surface expression of PD-L1 on isolated CD34+ HSCs was analyzed by using APC-conjugated anti-PD-L1 (clone, 130021) antibodies (FAB1561A, R&D System, USA). PE, vioblue or APC-conjugated anti-mouse IgG was used as isotype control. After washing with PBS, the cells were fixed with 1% paraformaldehyde and then analyzed by flow cytometry (MACSQuant Analyser, 2440, Miltenyi Biotec, Germany). Data were analyzed using MACSQuantify software (version 2.4). Live cells were identified by forward and side scatter. Gating was based upon isotype controls. Peripheral blood mononuclear cells (PBMCs) and HSCs were identified as CD45 and CD34 positive, respectively. The assays for HSCs and PD-L1 expression in each sample were performed in duplicate and the mean levels were reported. The results were expressed as a percentage and mean fluorescence intensity (MFI) per sample. Intra-assay variability based on repeated measurement of the same blood sample was low with a mean coefficient of variance of 4.3% in CHC patients and 4.15% in healthy control subjects.
Peripheral blood T- cell apoptosis assay
Peripheral blood T cell subset measurements were performed during each follow-up review. Percentages of T cell subsets were determined on 100 μl ethylenediamine tetra-acetic acid (EDTA) blood sample, using a dual-color direct immunofluorescence method (Miltenyi Biotec, Germany). CD3+ T cells were autogated and analyzed for CD8 expression using CD8/CD3 cell dual staining method. PE-conjugated anti CD3 and APC-conjugated anti CD8 were used for analysis. PE or APC-conjugated anti-mouse IgG was used as isotype control. Erythrocytes were lysed with lysis buffer (0.155 M NH4 Cl, 0.012 M NaHCO3, 0.1 mM EDTA, pH 7.2) for 5 min at room temperature. After washing with PBS, cells were stained simultaneously with FITC-conjugated Annexin V and propidium iodide (PI) (both from Miltenyi Biotec, Germany) according to the manufacturer’s instructions. During flow analysis, a live acquisition gate was placed on lymphocytes based on FSC/SSC profile. Side and forward angle light scattering were used to electronically gate the cells of choice and to exclude debris. Ten thousand events within the gate region were collected for each sample. Rare contaminating events were excluded on the basis of FSC/SSC. The percentages of CD3+ and CD8+ T cells that were annexin V and/or PI positive were measured by MACSQuant Analyzer using MACSQuantify software (version 2.4).
Effect of CD34+ HSCs on T-cell apoptosis
CD34+ HSCs and MNCs were prepared as described above. T cells (1×106) were cultured alone or co-cultured with CD34+ HSCs (1×105) in ratio of 1:10 or stimulated with PHA (50 mg/ml final concentration) for 3 days, harvested and quantified, stained with Annexin V kit (Miltenyi Biotec, Germany) and analyzed by flow cytometry (MACSQuant Analyzer, 2440).
Reverse transcriptase-polymerase chain reaction for PD-L1 expression on HSCs
The expression of PD-L1 on isolated CD34+ HSCs was assessed by reverse transcription- polymerase chain reaction (RT-PCR) amplification using gene-specific primers (PD-L1, F:5′-CACGGTTCCCAAGGACCTAT-3′and-reverse: 5′-TGGAGGATGTGCCAGAGGTA′). B-actin was amplified to demonstrate the integrity and the use of equal amounts of RNA and used as internal control (F: 5′-agcacagagcctcgcctttg-3′, R: 5′-tgacgatgccgtgctcgatg-3′, 267 bp). Total cellular RNA was extracted from purified CD34+ cells by using Trizol reagent (Invitrogen, CA, USA). RNA samples were treated with 10 μl DNase I (Roche Diagnostics, USA) per ml at 37°C for 30 min to decrease the contamination of genomic DNA, then heated at 70°C for 5 min to inactivate DNase I. Reverse transcription was performed using the (LightCycler® 480 System, Roche Diagnostics AQ5, USA) following manufacturer’s instructions. The expression of different transcripts was assessed by PCR amplification following standard protocols. The PCR products were analyzed on a 1% agarose gel stained with ethidium bromide.
Statistical analysis
Results are expressed as means±SD (range) or percent. Comparison between groups was made using the Student’s unpaired t-test. Pearson correlation analysis of PD-L1 expression on circulating CD34+ HSCs and T-cell apoptosis was performed. Differences were considered statistically significant at p<0.05. All calculations were performed using statistical computer package GraphPad Prism 7.03 for Windows.
Go to :
Results
Characteristics of study populations
Two groups of participants were recruited in this study: chronic HCV-infected patients and healthy controls. The demographic and laboratory characteristics of all participants were summarized in Table 1.
CD34+ HSCs in chronic HCV-infected patients
In an attempt to quantitatively analyze the circulating CD34+ cells in the peripheral blood of patients, the aliquots of blood samples of 30 CHC patients and 20 healthy controls were assessed by two-color flow cytometry. Cells were first gated according to forward/side scatter analysis (Fig. 1A). Then cells were further gated by CD45 positive staining (Fig. 1B). A representative of double-positive cells (CD34+/ CD45+) from one CHC patient is shown in Fig. 1C. As shown in Fig. 2, a baseline of the percentage of CD34+/CD45+ cells in the PBMCs ranged from 0.01 to 1.3% (0.5±0.42%) in healthy controls. In comparison with healthy controls, a significant increase in the percentage of circulating CD34+ cells, which ranged from 0.9 to 14.9% (5.4±3.7, p<0.0001), were observed in CHC patients. The average numbers of circulating leucocytes were (5.9±2×103/μl) in healthy controls and (6.82±1.8×103/μl) in CHC patients. No significant difference was found between two groups (p value>0.05) suggesting that elevated CD34+ cells in patients are not due to the elevation of circulating leucocytes.
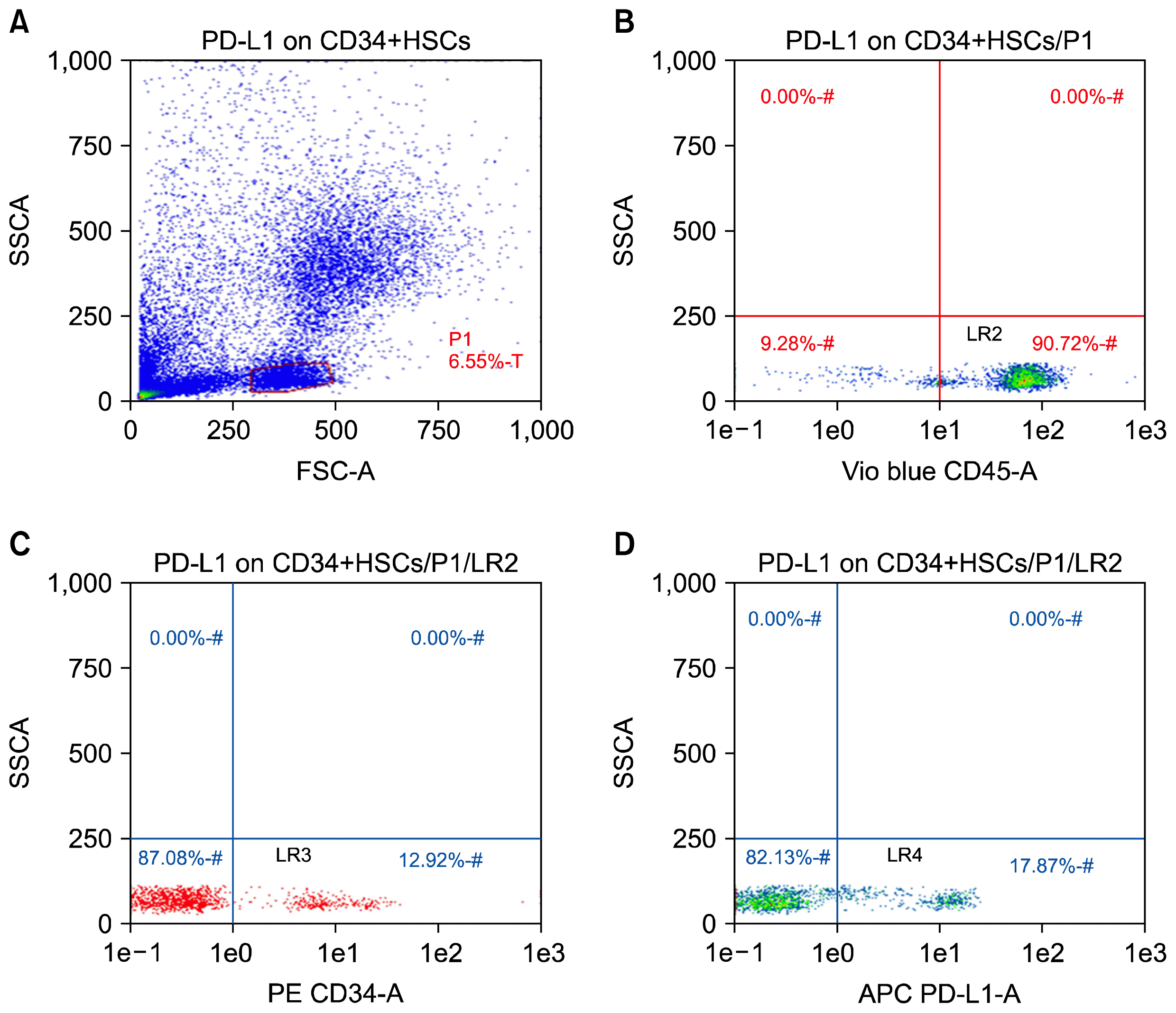 | Fig. 1PD-L1 expression on CD34+ HSCs in peripheral blood of chronic HCV-infected patients by flow cytometry. The peripheral blood CD34+ cells of patients and healthy controls were isolated by using MiniMacs high-gradient magnetic separation column and subsequently analyzed by flow cytometry using anti-CD45 and anti-CD34 antibodies. (A) A representative plot of enriched leukocytes shown by forward versus side scatter. The P1 gate represents the isolated PBMCs region. (B) The events in gate P1 are then displayed on a CD45 vs. SSC dot plot and a second gate (LR2) is produced to include the cluster of CD45+ events. (C) This plot is obtained by plotting the events that are fulfilling the criteria of gates P1 and LR2 (sequential gating). Cells forming a cluster with characteristic positivity to CD34 antibody are displayed in this plot creating the region LR3. (D) This plot is created by displaying cells that are positive for both CD45 and CD34 antibodies (LR3). Cells in lower right quadrant LR-4 are those with positive expression of PD-L1.
|
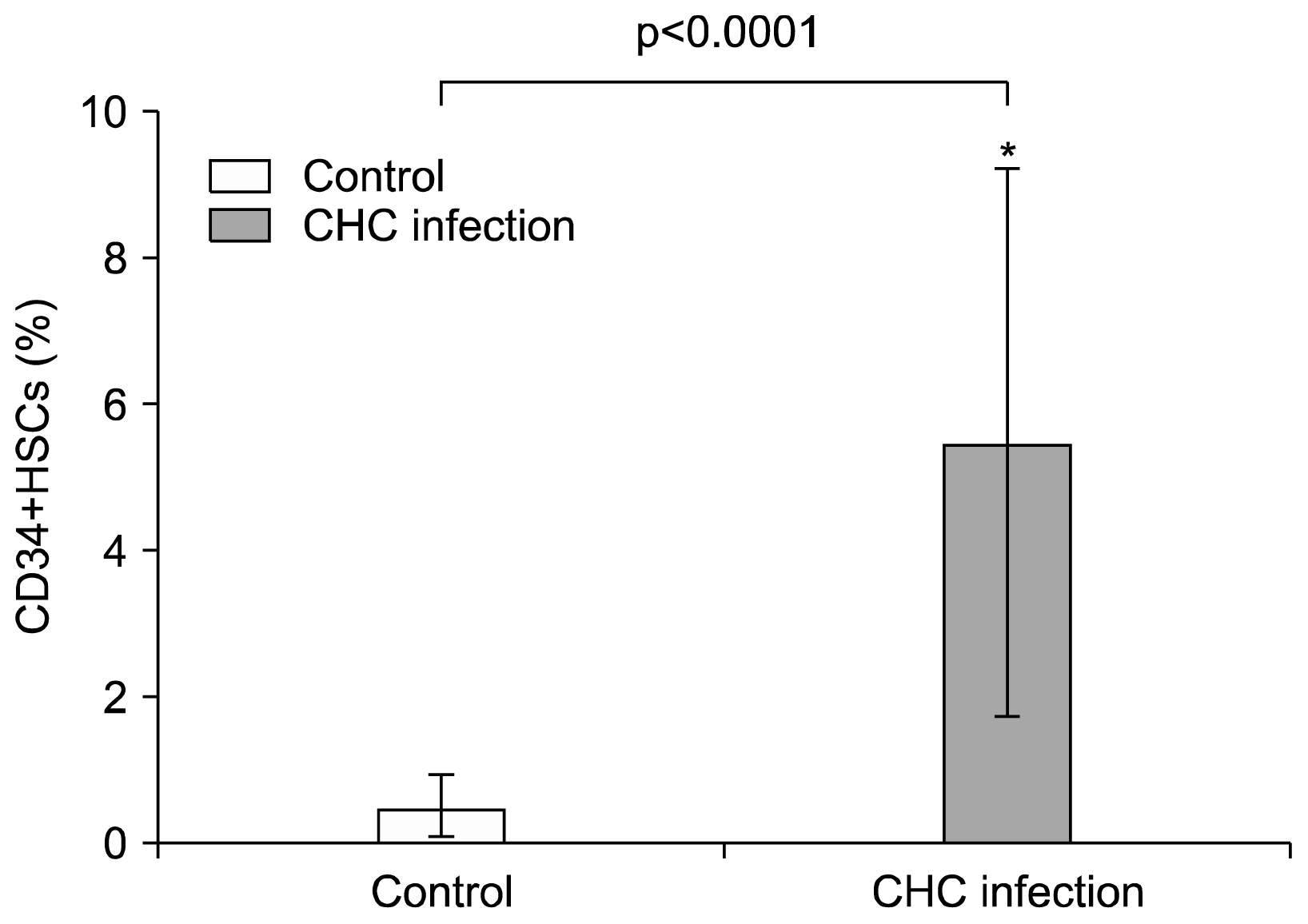 | Fig. 2Elevation of CD34+ HSCs in the peripheral blood of CHC patients. A Percentage of CD34+ cells in PBMCs fraction. Data represent the percentages of circulating CD34+ cells in the PBMCs fractions of 30 CHC patients and 20 healthy controls. Values are expressed as mean±SD. *indicates significant deviation from the control as indicated by p value<0.05.
|
PD-L1 expression level was elevated on CD34+ HSCs of CHC infected patients
Initially, we analyzed PD-L1 mRNA expression in isolated circulating CD34+ HSCs. In accordance with previous studies analyzing hematopoietic cells (17, 20) we detected PD-L1 mRNA expression in isolated CD34 cells (Fig. 3C). In addition to this, the PD-L1 expression levels on CD34+ HSCs in all 30 chronic HCV-infected patients and healthy controls were analyzed by FCM. Representative FCM plots of circulating PD-L1-positive CD34+ HSCs in one chronic HCV-infected patient are presented in Fig. 1D. Both the percentage of PD-L1-positive CD34 cells and the mean fluorescence intensity (MFI) of PD-L1 expression were used to evaluate the PD-L1 expression level. Our results showed that the percentage of PD-L1-positive CD34+ HSCs in chronic HCV- infected patients (17.5±5.4) was more than that in normal controls (8.3±3.2, p<0.0001) (Fig. 3A). Also, the MFI of PD-L1 expression in chronic HCV- infected patients (34.4±11.2) was significantly increased as compared with that in normal controls (11.3±4.8, p<0.0001) (Fig. 3B).
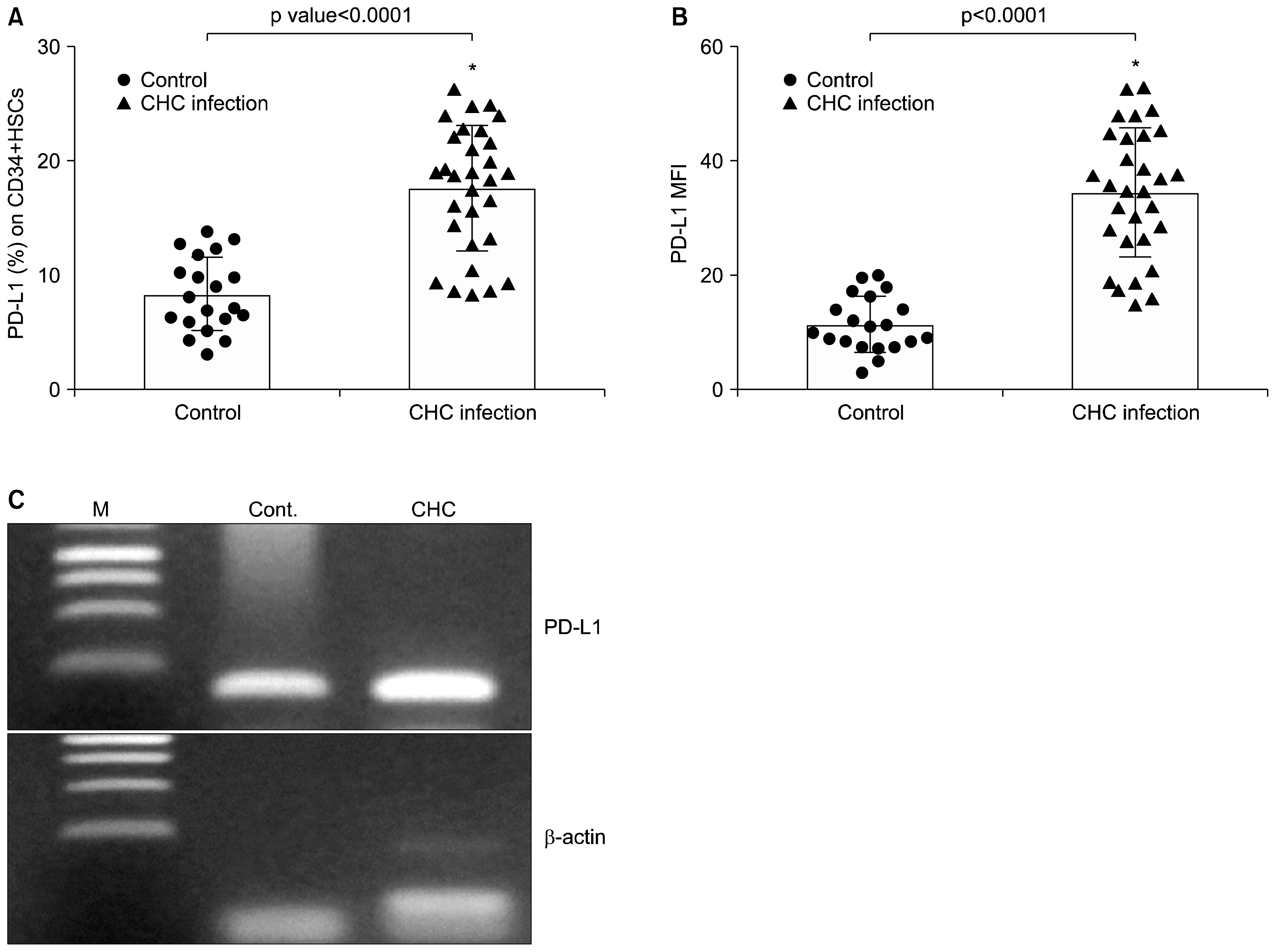 | Fig. 3PD-L1 expression on CD34+ HSCs in peripheral blood of CHC patients (▲) and healthy controls (●). Data were presented as percentage of positive cells (A) and mean fluorescence intensity (B). Graphs showed vertical scatter plots and mean columns. *indicates significant deviation from the control as indicated by p value<0.05. (C) Detection of PD-L1 transcript in purified CD34+ HSCs of CHC patients by RT-PCR. cDNA was synthesized from total RNA of purified CD34+ cells and β-actin was used as an internal control to demonstrate equal loading and integrity of RNA. M, marker, lane 1 healthy control (cont.) and lane 2 CHC patients.
|
Expression of PD-L1 in circulating CD34+ HSCs paralleled with T-cell apoptosis
Correlation between PD-L1 expression levels on CD34+ HSCs and degree of CD-8 T cell apoptosis was summarized in Fig. 4. Numbers of apoptotic CD8+ T cells were significantly increased in CHC patients (9.9±4) when compared with healthy controls (1.2±0.4, p<0.0001) identified by cells positive for Annexin V-FITC antibody (Fig. 5). PD-L1 expression on CD34+ HSCs (percentage and MFI) was strongly correlated with CD8+ T-cell apoptosis (Fig. 4A and B, r=0.8979 and 0.8448 respectively, p<0.0001) in patients chronically infected with hepatitis C virus. T-cells apoptosis using Annexin V kit was detected when cells were co-cultured HSCs, the ratio of T-cell apoptosis was 1.3±0.36 in absence of CD34+ HSCs, but 10.35±4.9 in presence of CD34+HSCs (T- cell apoptosis rate was statistically different, p<0.0001).
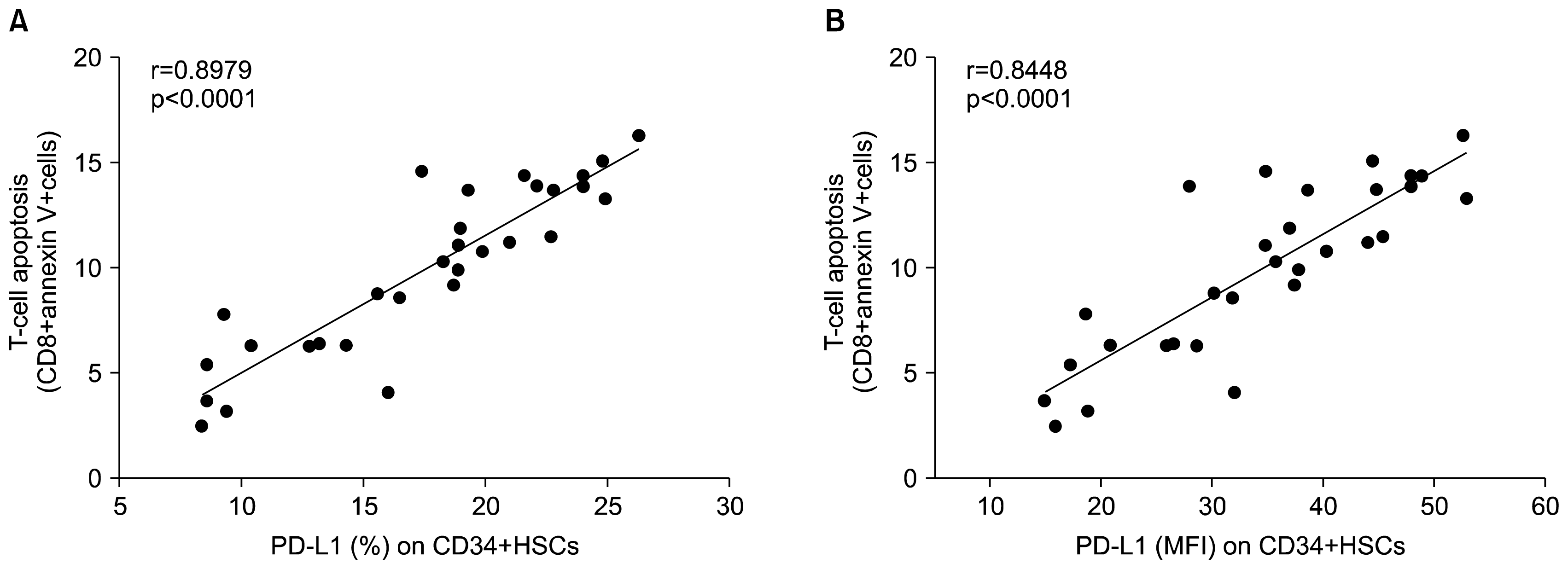 | Fig. 4Correlation analysis of expression of PD-L1 on CD34+HSCs and T-cell apoptosis (CD8+ Annexin V+ cells) in chronic HCV infected patients. (A) Percentage (%) of PD-L1 expression (B) Mean fluorescence intensity (MFI). r=0.8979 and 0.8448 respectively. p<0.0001.
|
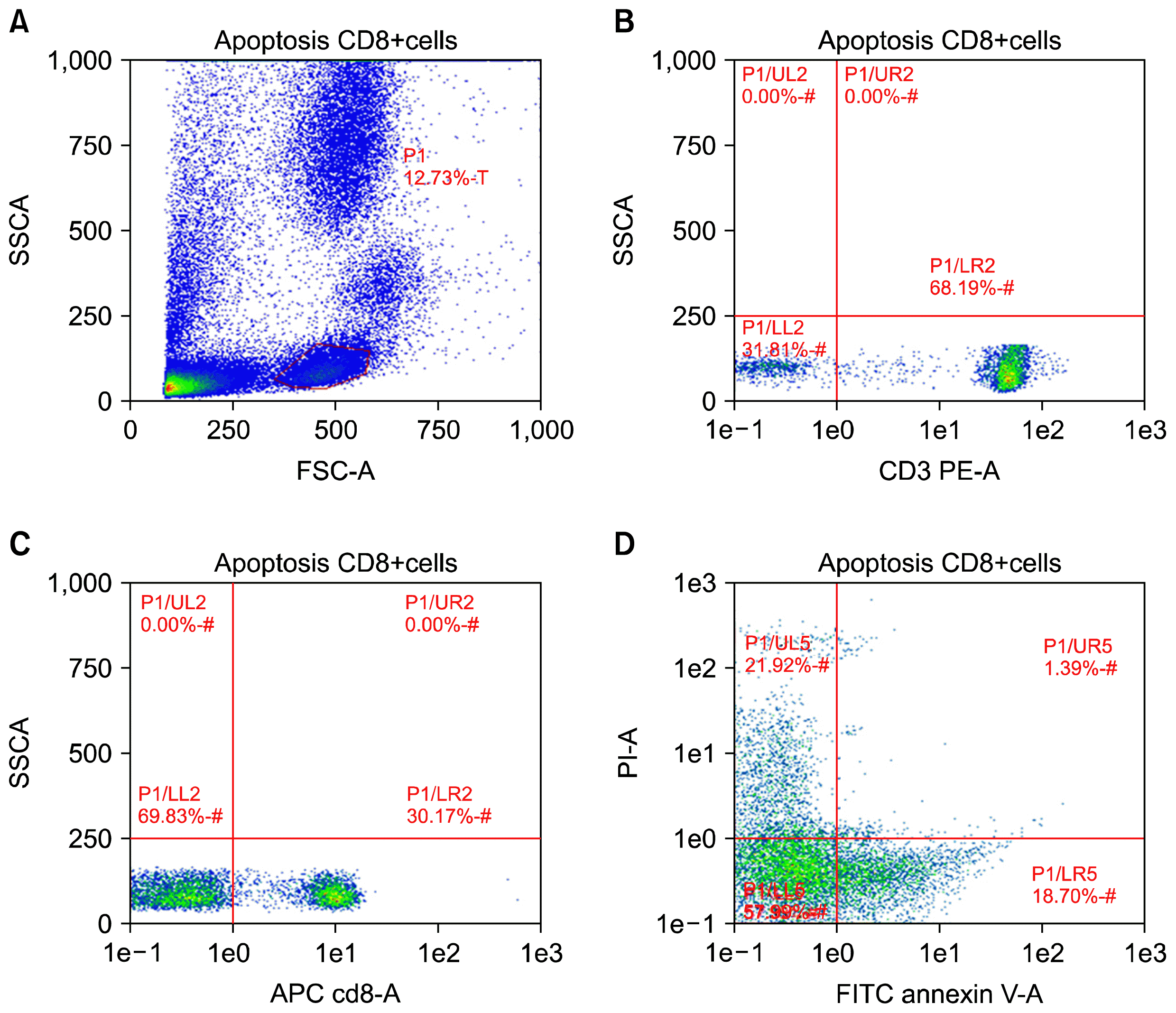 | Fig. 5Expression of PD-L1 on CD34+ HSCs mediated CD-8 T-cell apoptosis in peripheral blood of chronic HCV-infected patients. Apoptosis in T-cells was examined by flow cytometric analysis using AnnexinV/PI staining. Numbers of apoptotic cells (positive for Annexin V stain) are indicated in plot D. One of three independent experiments is depicted.
|
Go to :
Discussion
During the last decades, host adaptive immune responses and especially the effects of antiviral CD8+ T cells have been shown to play an essential role in the elimination or control of the most important human viral infections. Thus, a virus-specific CD8+ T-cell response can clear the virus by cytolytic and non-cytolytic effector mechanisms. Importantly, however, these viral infections can lead to viral persistence. Compelling progress has been achieved in understanding mechanisms of T-cell effector function and failure. These findings are important for understanding of immunological factors of virus–host interactions, as well as antiviral vaccine development strategies against HCV and viral immune-therapies that help to cure chronic HCV infections (21). However, there are still several open questions that need to be addressed in order to completely explore T-cell failure in viral hepatitis. Indeed, it worth noting that while the effector functions of HCV-specific CD8+ T cells are mainly elucidated, other aspects of the immune response of CD8+ T cells are less understood. For example, the precise mechanisms of liver homing of virus-specific T cells, effect of general proliferation signals mediated by HCV infection on virus-specific T cells, impact of stem cell-mediated liver repair and cell to cell contact on T- cells are still not completely discovered. Thus understanding these factors may help to catch on the underlying mechanisms of T cells failure and thus chronic disease progression.
We demonstrated that circulating CD34+ HSCs constitutively express low levels of PD-L1, while its expression is strongly enhanced by activated T cells and viral infection. Moreover, PD-L1 expression on HSCs induces apoptosis in T cells. The detection of constitutive PD-L1 expression in human HSCs is novel and surprising, since a recent study analyzing PD-L1 expression detected antibody staining exclusively in culture expanded murine hematopoietic stem cells (22). Currently, only a few studies have analyzed PD-L1 expression in tissues from different species. In some of those, different PD-L1 tissue expression patterns were observed comparing tissues of murine and human origin (23–27). Therefore, human HSCs were analyzed confirming PD-L1 expression species independently in circulating HSCs.
Accumulating evidence has revealed that the BM-derived human HSCs can migrate to the liver and contribute to liver regeneration. Results from gender mismatch liver transplantation studies proposed that BM-derived HSCs of host origin can differentiate into hepatocytes and cholangiocytes in transplanted livers, while hepatocyte chimerism was present in the patients affected by recurrent hepatitis (28, 29). Wynn, 2007 stated that inflammation is a critical factor in the initiation and maintenance of liver fibrogenesis and cirrhosis from hepatitis viral infection. Inflammation reactions and damaged cells in liver release inflammatory mediators, such as TGF-beta1 and TNF-alpha, which can recruit reacting cells in circulating blood to the liver (30). Thus, it seems that elevated CD34+ levels in peripheral blood of CHC patients are related to persistent liver inflammation and imbalance between injury and endogenous repair capabilities.
Our results demonstrate PD-L1 mRNA expression in HSCs isolated from patients with chronic HCV infection, in addition to its expression in quiescent cells (isolated from healthy controls) but at lower levels (by both positive percent expression and mean fluorescent intensity). In line with these findings, Zheng et al. (22) determined the expression of CD274 (PD-L1) on freshly isolated and culture expanded human cord blood HSCs. While only ~10% of freshly isolated human CD34+ cells express CD274 on their surface, the CD274+ population increased to more than 50% after culture activation. Basic expression but even more inducible induction of PD-L1 on HSCs may have several implications. The observed profound up-regulation of PD-L1 expression on circulating HSCs may serve as a part of a protective negative feedback mechanism in T cell-mediated inflammatory events. Interestingly, a study in PD-L1-deficient mice suggested that PD-L1 is a key protein regulating the accumulation and deletion of intrahepatic CD8+ T cells (31). We found increased levels of PD-L1 upon viral infection (Fig. 3A and B). In line with our findings, previous studies found an up-regulation of PD-L1 expression in airway epithelial cells in response to double-stranded RNA (32) and in lymphocytes in response to HIV infection (27). Interestingly, there was a strong correlation between degree of PD-L1 expression and T-cell apoptosis detected by Annexin V staining in both peripheral blood and ex-vivo when isolated CD34+ HSCs were co-cultured with T lymphocytes (r=0.8979, p<0.0001).
Our findings may shed new light on bidirectional interaction between CD34+ bone marrow (BM) derived HSCs and lymphocytes. HSCs isolated from peripheral blood of healthy controls express low level of PD-L1. Exposure to general inflammatory and proliferation signals mediated by chronic HCV infection results in up-regulation of PD-L1 on HSCs. That, in turn, induces apoptosis of activated lymphocytes. Thus, circulating HSCs beside its role in liver regenerative process and restoration of liver mass upon chronic infection may contribute to immune tolerance induction by deletion of activated T cells through induction of apoptosis. This may provide new insight on the possibility of isolation and culture expansion of HSCs and may benefit allogeneic transplantation for patients. The study has limitations; first, only PD-L1 was detected in our work, other co-stimulatory molecules expressed on stem cells that can modulate the immune response were not elucidated. Second, despite our interesting findings, the sample size of our series is relatively small. All conclusions based on our results are, therefore, tentative and need further support from well-controlled clinical studies of a larger scale with a wide range of study subjects.
In summary, our study demonstrated that PD-L1 is readily expressed upon HSCs. Its expression is up-regulated with chronic HCV infection. There is strong correlation between PD-L1 expression on HSCs and degree of T-cell lymphocyte apoptosis. Expression of such an immune inhibitory signal on HSCs may provide new insight into the process of allogeneic transplantation of HSCs. Cellular mechanism by which PD-L1 co-stimulates T cell response and in other circumstances these responses are inhibited remain elusive and need to be explored further.
Go to :
 XML Download
XML Download