

 PDF
PDF Citation
Citation Print
Print


Introduction
Human mesenchymal stem cells (MSCs) are able to self-renew or differentiate to other lineages (1–3), and these cells have been isolated from different tissues such as brain, liver, bone marrow, adipose tissue, foetal tissues, umbilical cord (UC), and placenta (4–7). Various types of human mesenchymal stem cells (MSCs) reside in the hypoic microenvironment, which seems to be conductive to stem cell longevity and the physiological niches (8, 9). Hypoxia has a strong effect on several aspects of cell biology such as metabolism, angiogenesis, innate immunity and stemness induction, and hypoxic is also essential for the self-renewal and the maintenance of multipotency of human MSCs and hematopoietic stem cells (HSCs) (9–11). In recent years, many studies support that the hypoxic culture of human MSCs inhibits cellular senescence, maintains MSCs properties, augments the differentiation capacity, and enhances their tissue regenerative potential, indicating that hypoxia increases the lifespan and the differentiation potential of MSCs (10–13).
MSCs-based cell therapy is a potential therapeutic approach for the treatment of various diseases, including stroke and myocardial infarction (14–17), traumatic brain injuries (18), diabetes mellitus (19), inflammatory bowel disease (20), and acute kidney (21, 22) and liver injuries (23–25). However, the large amount of engrafted MSCs decreased dramatically after transplantation due to immune rejection and toxic microenvironments. Thus, adopting appropriate priming strategies provide an effective way of promoting survival and avoiding immune rejection (14). In addition, recent studies have also shown that hypoxic priming induces the expression of pro-survival markers (26), and growth factors involved in cell proliferation, anti-apoptosis and angiogenesis (27) in MSCs. However, it is not yet clear whether the benefit of hypoxic priming is the expansion, cellular longevity, or multi-potent differentiation capacity of human MSCs. In this study, we found that hypoxic priming extends cell cycle lifespan but reduce genetic damage susceptibility, and thus maintain the multipotency of MSCs during differentiation lineage.
Materials and Methods
Cell culture
Human umbilical cord blood derived mesenchymal stem cells (hUCB-MSCs; PromoCell) were grown in Dulbecco’s Modified Eagle’s Medium (DMEM; Hyclone) containing 10% fetal bovine serum (FBS; GIBCO) and 1% Penicillin/Streptomycin antibiotics at 37°C in a 5% CO2 incubator with 21% O2 (normoxia) or 1% O2 (hypoxia).
Carboxyfluorescein succinimidyl ester (CFSE) assay
For assessment of the potential of cell proliferation, MSCs were trypsinized and washed once with phosphate buffered saline (PBS). CFSE (Invitrogen, 10 mM in PBS) was added to the cells and incubated at 37°C in the dark for 15 min. An equal volume of serum containing growth medium was added for quench the CFSE reaction. Cells were again incubated at 37°C in the dark for 5 min. CFSE-labeled MSCs were washed twice with growth medium and seeded in a culture plate.
Senescence associated β-galactosidase (β-gal) assay
Cells were fixed with 4% formaldehyde for 10 min and incubated overnight at 37°C with 1 mg/ml X-gal (5-bromo-4-chloro-3-indolyl-β-D-galactopyranoside), 5 mM K3Fe(CN)6, 5 mM K4Fe(CN)6, 150 mM NaCl, and 2 mM MgCl2 in 40 mM citric acid/sodium phosphate pH 6.0.
Immunofluorescence
hUCB-MSCs were grown on coverslips (1×104 cells) for 2 days. Cells were fixed with 4% formaldehyde in PBS for 10 min and rinsed three times with a wash solution of (0.1% Triton X-100 in PBS. Cells were blocked with 3% skim milk/wash solution for 30 min and incubated with primary antibody (1:200) and secondary antibody (1:200) for 1 hr.
Metaphase chromosome spreading assay
Cells were incubated with colcemid (final concentration 100 ng/ml) for cell cycle arrest around metaphase. Mitotic cells were then collected by gentle pipetting, washed with PBS twice, counted, and suspended in a hypotonic solution (0.8% sodium citrate). After 15 min incubation at room temperature, swelled cells were centrifuged (1,000 rpm, 4 min, 4°C). Most of the supernatant was aspirated and the pellet was gently in the remaining approximately 500 μl of supernatant. Carnoy’s fixative solution (75% methanol, 25% acetic acid) was slowly added and incubated at room temperature for 10 min. The fixation was repeated 3~4 more times. The suspended cells were dropped onto cold wet slides and allowed to dry at room temperature. The slides were mounted and examined by fluorescence microscopy (Carl Zeiss).
Statistical analysis
All data were presented as means±standard errors of means (SEMs). Statistical analyses were performed using the Student t test.
Fluorescence-activated cell sorting (FACS)
Human UCB-MSCs were evaluated using surface marker detection at passage 5 (P5) to confirm the effect of oxygen concentration (hypoxia and normoxia) on MSC characterization. hUCB-MSCs at 80% confluence were harvested and suspended in FACS buffer (1×107 cells/ml). Antibody was then added to each samples: Anti-CD90 fluorescein isothiocyanate (FITC), Anti-CD44 phycoerythrin (PE), Anti-CD105-PerCP-Cy5.5, Anti-CD73-allophycocyanin (APC), MSC negative antibodies set (Anti-CD34/CD11b/CD19/CD45/HLA-DR-PE), positive isotype cocktail (mIgG-FITC, mIgG-PerCP-Cy5.5, mIgG-APC), negative isotype cocktail (mIgG1-PE, mIgG2a-PE) (562245, BD Biosciences) followed by incubation at 4°C for 30 min. For cell cycle analysis using propidium iodide, cells were fixed with cold 70% ethanol overnight at −20°C. They were then washed once with PBS and incubated in PBS containing 50 μg/ml propidium iodide and 1 mg/ml RNase A for 30 min at room temperature. After staining, cells were washing with PBS and measured by flow cytometry. Antibodies listed above are purchased from BD Biosciences.
Results and Discussion
Hypoxia extends cell cycle lifespan and multi-potency of humans MSCs
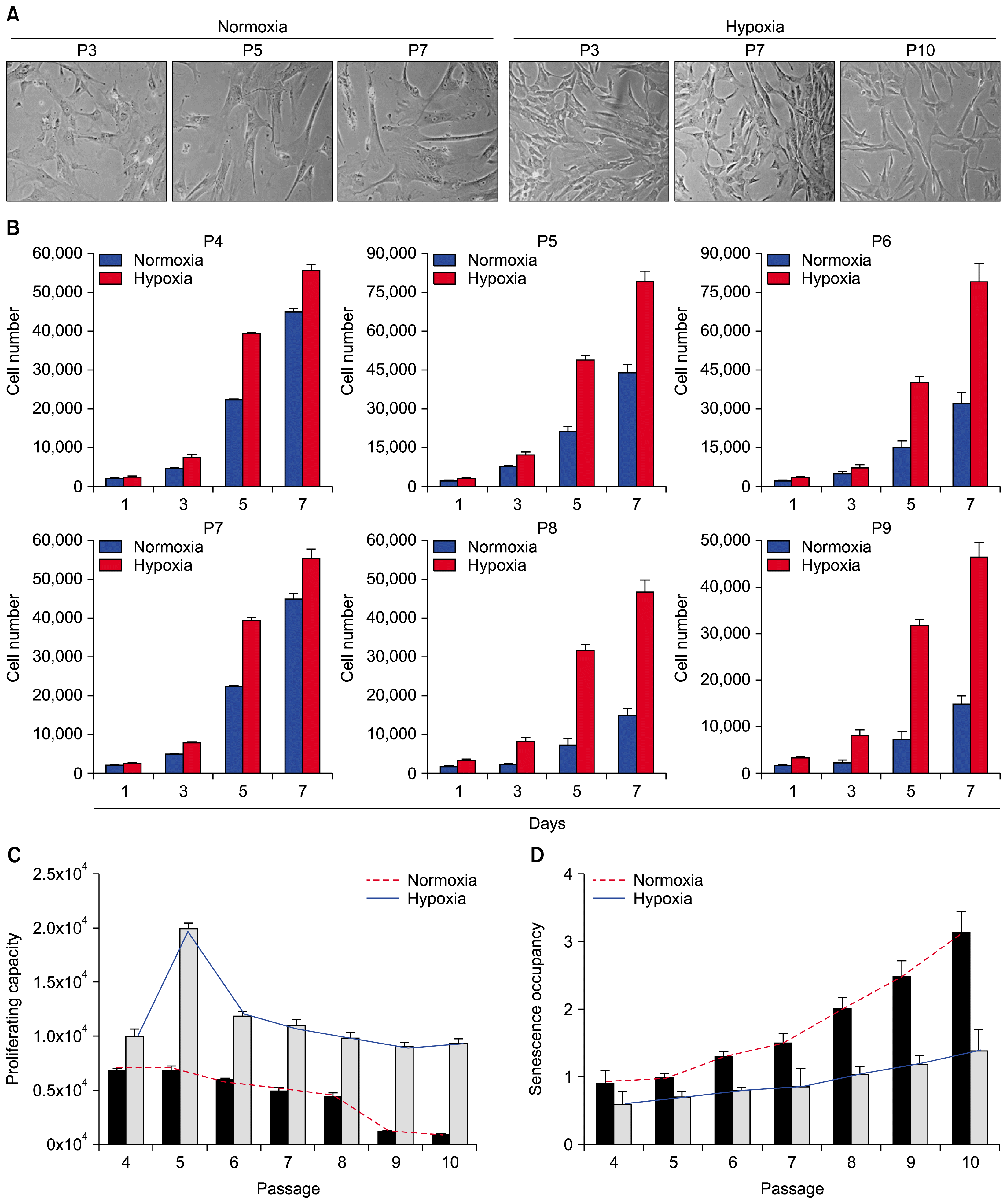
To delineate the key mechanistic benefit of hypoxic priming, human MSCs were isolated from human umbilical cord blood (UCB) by adhesion to tissue culture-coated plates in complete culture medium as previously described (14). Culture passaging generated a homogenous population of UCB-MSCs (hereafter MSCs) that were used to measure proliferation rate, multi-potency, and senescence (Fig. 1A~1C). Human MSCs were cultured under normoxic (21% oxygen) or hypoxic (1% oxygen) conditions. When the culture became nearly confluent, the cells were trypsinized and subcultivated. As expected, hypoxia significantly accelerated the proliferation rate and extended the life span of human MSCs compared to normoxic cultures (Fig. 1B and 1C). Similar results were obtained with MSCs from two different donors (data not shown).
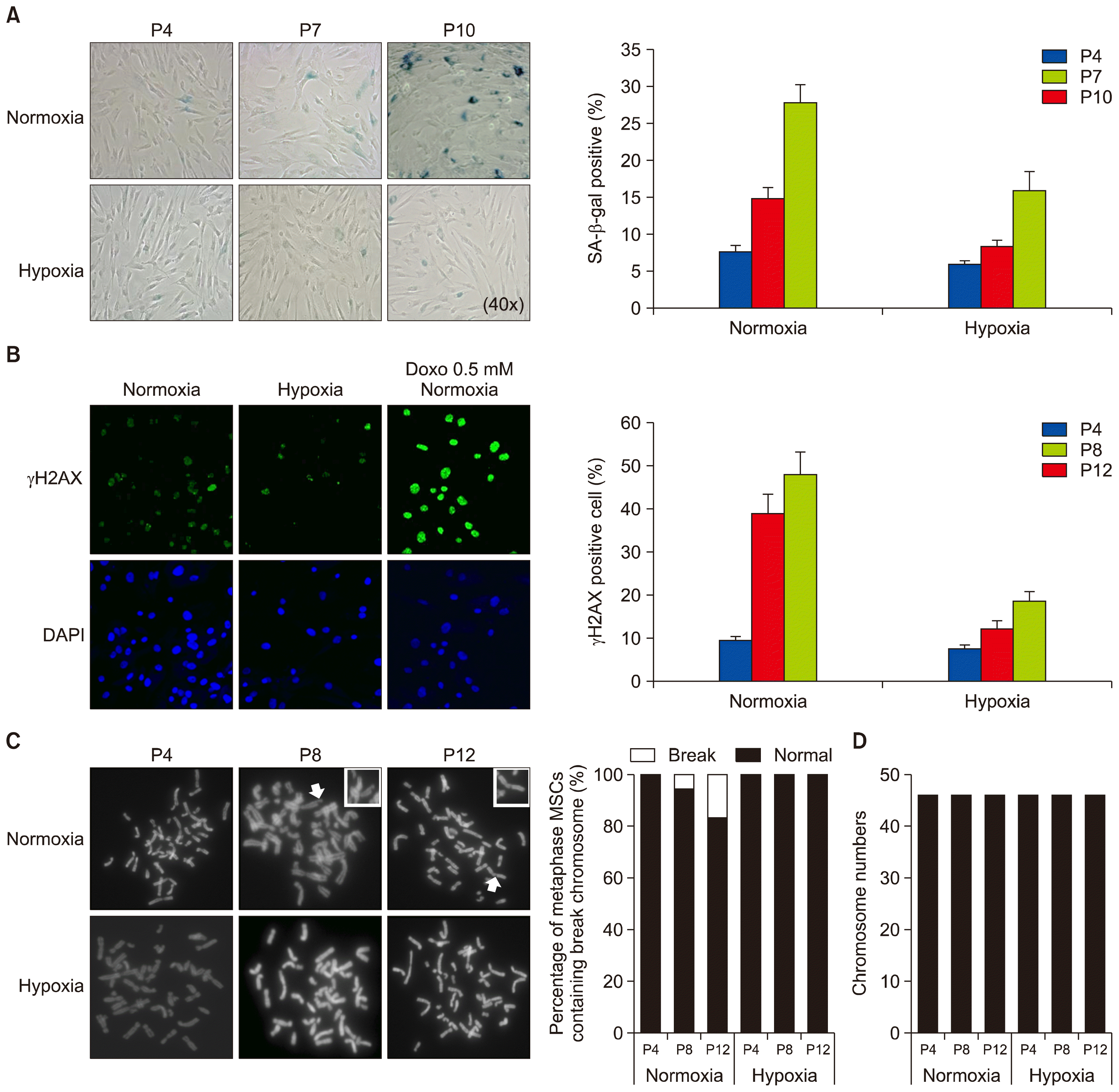
Under the hypoxic condition, late passaged MSCs continuously proliferated and maintained a similar morphology as early passaged cells (Fig. 1B and 1C). However, most of late passaged MSCs (passage 8 and later) cultured in the normoxic condition almost completely lost their proliferation capacity and adopted a flat, enlarged shape and ceased proliferation at subconfluent densities, thus manifesting characteristics of senescence (Fig. 2A), indicating that hypoxia extended the cell proliferation life span of MSCs. In support of this notion, cytochemical senescent phenotype analysis involving staining of the senescence-associated β-galactosidase (SA β-Gal) showed that cells in normoxic cultures of late passaged MSCs mostly displayed SA β-Gal activity, while hypoxic cultures showed significantly less amounts of SA β-Gal staining (Figs. 1D and 2A). In addition, the percentage of γH2AX-positive cells (as a marker for DNA double strand breaks) was markedly decreased compared to normoxic culture (Fig. 2B). Further metaphase chromosome spreading assays also showed significantly less population of MSCs with chromosome damage (Fig. 2B and 2C). Together, these results indicate that hypoxia extends cell cycle lifespan but reduces chromosome damage susceptibility.
Uncoupling between cell cycle lifespan and multi-potency of hypoxic primed humans MSCs
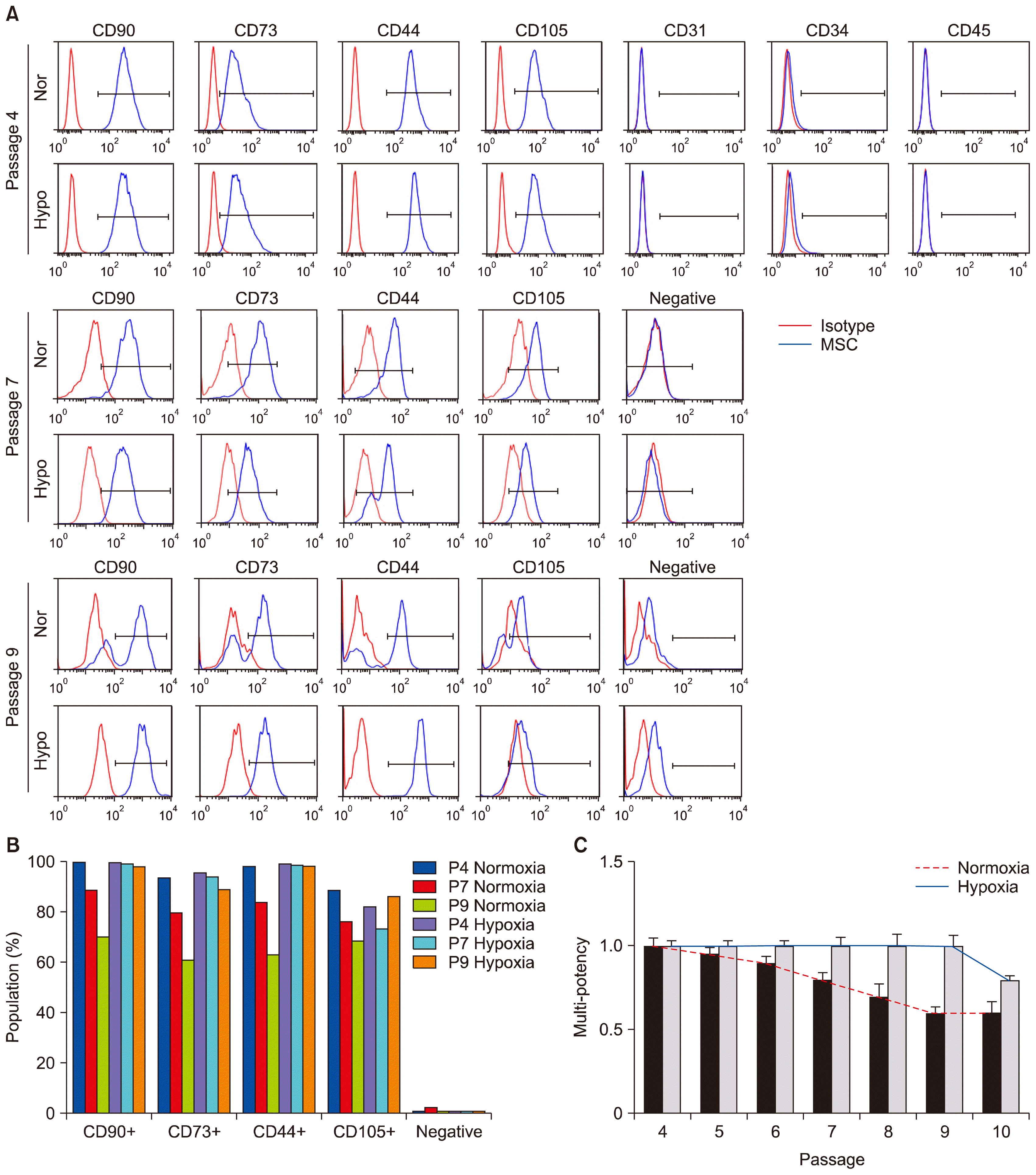
Multi-potency of MSCs by normoxic and hypoxic priming was compared by the examination of the surface expression of multi-potent MSC antigenic markers. Positive markers included CD44, CD73, CD90, and CD105. Negative markers included CD34, CD45, CD11b, and/or CD19 (Fig. 3A and 3B). Unexpectedly, early and middle passage MSCs (passages 4 and 7, respectively) grown under normoxic or hypoxic condition showed very similar levels of surface CD marker profiles; cells were consistently positive for CD44, CD73, CD90, and CD105, and negative for CD34, CD45, CD11b, and/or CD19 (Fig. 3B and 3C), indicating that hypoxic priming does not augment the differentiation capacity of MSCs at early and intermediate passages. However, although hypoxic conditioning drastically extended the cell cycle lifespan and delayed senescence in late passaged MSCs, comparison of the multi-potency by measuring the same number of cells revealed that normoxic primed late passaged MSCs still retained the significant capacity of multi-potency compared to hypoxic conditioning. These results raise the important notion that hypoxic priming is an efficient tool for expanding the actively proliferating pool of MSCs, but not for augmentation of their multi-potency. The multi-potency of MSCs may be able to be determined by measuring their proliferating potential and/or strength of cell cycle lifespan, but not by the expressions of pre-existing MSC markers.
 XML Download
XML Download