

 PDF
PDF Citation
Citation Print
Print


Introduction
hUCB-derived CD34+ progenitor/stem cells transplantation studies in animal models have indicated that these cells are able to circulate in the blood and home into sites of typical injured tissues to recover normal tissue function (1). Mesenchymal stem cells (MSCs) from human umbilical cords ameliorate mouse liver injury or disease in vivo (2). Moreover, infused MSCs participate in liver regeneration and functional improvement through reducing liver fibrosis induced by S. mansoni in adult mice (3).
hUCB is another source of stem and progenitor cells for transplant, being of fetal origin with considerable proliferative potential, and was recognized as an alternative source for therapeutic applications, clinical transplantation and regenerative medicine (4–7).
CD34+ cells from the hUCB are somatic non-differentiated progenitor/SCs (8). It can differentiate into multiple of specialized cell types but are less plastic in their differentiation capacity (9). That means that they can only differentiate to two or more different specialized somatic cell types along a developmental path, making them multipotent (10).
Schistosomes (blood flukes) are parasites, mainly belonging to three species of the trematode Schistosoma: S. mansoni, S. japonicum and S. hematobium. Humans have suffered and still suffer greatly from schistosomiasis. S. mansoni occurs in Africa, South America, Caribbean and areas of the Middle East (11). The hyperendemic infection with S. mansoni was exceptionally so common in the Nile delta, Egypt, a situation persisting into this day (12, 13).
In S. mansoni chronic infection the liver is damaged by deposition of eggs. The granulomas serve to contain the eggs deposited in the liver due to a peculiar host response against eggs. The granulomas resolve as the eggs die and leave severe fibrosis. S. mansoni granulomas in infected individuals are of different composition reflecting their different age (14).
Xu and co-workers (15) have reported that MSCs were able to ameliorate S. japonicum-induced liver fibrosis in vivo and this effect was enhanced by combining MSCs with conventional drug praziquantel (PZQ) therapy to treat S. japonicum-infected mice.
In this study, we aimed to investigate the potential effects of CD34+ cells isolated from hUCB on laboratory adult mice infected with the parasitic flatworm S. mansoni-induced liver injury de novo and the subsequent consequences in vivo.
Go to : 
Materials and Methods
Parasites and laboratory animals
A total of 60 female and male Swiss albino mice (Mus muscullus, CD1 strain), 6∼8 weeks old and weighting 24±1 g, were purchased from the Schistosoma Biology Supply Center (SBSC) at Theodor Bilharz Research Institute (PO Box 30 Imbaba, Giza, Egypt). Each mouse was infected through the abdominal skin via subcutaneous injection with about 100 pre-harvested cercariae of the human parasitic flatworm S. mansoni (Egyptian strain) to induce liver fibrosis. All procedures involving animals were performed in accordance with the guidelines of the Institutional Animal Care and Use Committee of Theodor Bilharz Research Institute (Giza, Egypt).
Mice were transported to the animal care facility of the Department of Zoology at the Faculty of Science, Suez Canal University. The Mice were housed in standard individual mouse cages (10 animals per two cages) for two weeks before treatment and cages placed in acclimatized rooms with a controlled 12 hours light/12 hours dark cycles. Animals had reared in free access to standard pellet animal diet and tap water, constant room temperature and humidity, bedded on wood shavings and checked upon twice daily for health status. All animal experiments were approved by the local committee for laboratory animals research care and use set by the Department of Zoology (in Faculty of Science, Suez Canal University).
Mice were equally divided into seven groups (groups 1–7). Group 7 was used as a normal uninfected negative control group and kept in a similar manner, and the same guidelines as mentioned above were followed. Mice of group 7 were also purchased from the Experimental Animal Center at the Theodor Bilharz Research Institute (Giza, Egypt). Mice in groups 1–6 were injected with either 0.3 mL of phosphate-buffered saline (PBS; intravenous), or hUCB-derived CD34+ cells (1×105 cells suspended in PBS; intravenous). PBS was purchased from Gibco (Germany). Mice in group 6 served as infected positive controls without any type of treatment and were assigned for the sake of comparison; mice in groups 1-5 received hUCB CD34+ cells (1×105 cells/mouse suspended in 0.3 mL PBS; intravenous IV).
In this study, we avoided the use of immune-deficient mice for several reasons, including; difficulties interpreting the results owing to the interference linked to the immunological consequences during schistosomiasis, a slow immune reconstitution after CD34+ cells transplantation and a naivety of the cells reducing the risk incidence of immune rejections (5), the ability to induce teratomas or death when injected into immune deficient mice in vivo (16) and the need for specific assays to develop laboratory adult mice infected with S. mansoni creating liver fibrosis where intact immune system is required.
Isolation of the hUCB CD34+ cells
The hUCB samples were collected immediately after normal full-term maternal donor deliveries, pre-approved by the local Ethics Committee at Suez Canal University Hospital, and used for the isolation of mononuclear CD34+ cells. The collected placental (cord) blood was delivered in sterile collection tubes (50 mL) containing CPDA-1 as an anti-coagulant (5 mL). Blood was diluted 1:1 with Dulbecco’s phosphate-buffered saline (PBS). A density-gradient centrifugation at 800 g for 20 min at 20°C using Ficoll-paque (1.077 g/mL; Lonza Bioscience) was performed to collect the buffy coat layer according to the manufacturer’s protocol. After Ficoll-paque gradient separation, collected hUCB MNCs were washed twice in Dulbecco’s PBS. Isolation of the positively selected CD34 cells was then carried out using immunomagnetic separation technique by immune beads of Dynal CD34+ progenitor cell selection system (MACS®) following the procedure protocol according to the manufacturer’s recommendations. The medium used for derivation and separation of CD34+ cells consists of lymphocyte separation medium according to manufacturer’s descriptions. The total number of CD34-postive cells in suspension was assessed with the electronic-based cell counter device, according to the manufacturer’s instructions (Gen S, Tectron).
hUCB CD34+ cells transplantation
For transplantation, the percentage of the positively selected CD34 cells were calculated and adjusted to achieve the desired number of cells/dosage. We sought to achieve the desired doses of 1×105 cells/mouse. A volume of 0.3 mL of PBS solution was added to the viable CD34+ cells pellet for final volume 1×105 cells. CD34+ cells were transplanted into mice of all different groups by IV injection into the lateral tail vein in a dose of 1×105 cells/mouse using a Hamilton syringe. Fifty mice received CD34+ cells injections. Six mice showed no transplantation survival and three died before started the CD34+ cells treatment.
Specimens sampling
After transplantation with the same doses of 1×105 cells/mouse, specimens sampling were done to treated and control mice groups on 24 hour, 48 hour, 1 week, 2 weeks and 3∼4 weeks with regular measurement of ova count per 0.1 gram of feces for each group after specific time interval until 4 weeks. Deposition of eggs as well as the number of liver granulomas was examined. Blood sample was withdrawn from the retro-orbital venous plexus and was kept in room temperature to be clotted. Thereafter it was centrifuged for 15 min. Serum was collected and stored at −20°C for measurement of serum alanine (amino) transferase (ALT) level and serum albumin (ALB) secretion by the standard photometric method for each group specific time interval until 4 weeks for comparison of concentrations in the S. mansoni-infected post-treated mice groups in those with, and without, S. mansoni control animals.
Specimens sampling were taken from normal uninfected and S. mansoni infected control mice groups at all different time points during the course of the experiments: immediately, on 24 hour, 48 hour, 1 week, 2 weeks until 4 weeks. For simplicity of interpretation of results compared with that of S. mansoni-infected post-treated experimental mice groups, data were tabulated on average and statistically analyzed to evaluate the difference between the groups under study as regards the various parameters.
Preparation of histological specimens
On death or killed by cervical dislocation, portions from liver samples of each mouse were taken aseptically and immediately fixed in 4% paraformaldehyde. The liver tissues from all different groups were embedded in paraffin and sliced into 4 μm sections, dewaxed and rehydrated, then stained with hematoxylin and eosin (H & E) staining for histological examination. The histopathological data on liver stained sections were photographed and analyzed to confirm the biochemical results. All slides were kindly reviewed by pathologists at the Department of Pathology (Faculty of Medicine, Suez Canal University) using a double headed microscope. Only granulomas that appeared circular in form were considered.
Stool examination and eggs per 0.1 gram (EPG)
We counted the number of eggs per 0.1 gram (EPG) stool and a cumulative arithmetic mean for each group was calculated with the assumption that feces were collected for 24-hour time periods into a small open beaker which were then fixed in 2 mL of 10% neutral-buffered formalin, dispersed in a food processor for 2 minutes at a half speed, and the resulting suspension was passed through nylon cloth. Eggs were counted in a haemocytometer using 100× magnification under a light microscope with an equivalent of 0.1 gram of feces/mL until the fecal samples was calculated, i.e., if 10 μl was examined, 0.5% of eggs/2 mL was used for the calculation.
Statistical analysis
All obtained data from the experimental groups were subjected to statistical analysis by performing one-way analysis of variance (ANOVA) for comparison of normal and S. mansoni-infected control groups versus infected post-treated mice groups. The post-treated experimental groups were divided into five categorical groups: early (24 hour, 48 hour), mid (1∼2 weeks), and late (3∼4 weeks), and ANOVA was used to compare the median level in each of these groups. The data collected for the differences between animal groups were compared by paired student’s t-test when appropriate. Differences between groups were considered to be statistically significant at a p-value of <0.05. The statistically significant values suggested that the variation explained between groups was not due to chance. The results data are presented as means±standard deviation (SD) of mean in all cases. Figures were made using GraphPad PRISM 3.0 statistical software package, for categorical data.
Go to :
Results
CD34+ preparation (cells/hUCB)
The yield of the selected CD34+ in the fetal-hUCB-derived cells approximately ranged from 4 to 5.5×106 cells/hUCB and calculated based on the total count of collected hUCB mononuclear cells (MNCs). The CD34+ cells were then adjusted to a number of 1×105 viable cells in 0.3 mL of PBS solution for final volume using an electronic cell counter device (Gen S, Tectron). The CD34+ cells data collection information was also recorded for age (years) of each maternal-donor deliveries and gender (male or female) of the newly delivered infant. The dates of CD34+ cells preparation and treatment into experimental groups were also recorded which might also affect this study rational approaches of this work (data not shown). Moreover, the time of CD34+ cells treatment in treated experimental groups must be within the first day from the date of CD34+ cells preparation.
Serum liver function parameters
In this work we selected two important routine laboratory tests of assessment of liver function parameters, serum concentrations of albumin (g/dL) and also the ALT (U/L) parameter. We noted that the mice infected with S. mansoni followed by treatment of CD34+ cells have a strong association with the improvement of the serum albumin concentrations (g/dL) level at the 2 weeks of treated animal groups (mean=4.1±0.2 SD) and showed no significant differences to normal control mice (mean=4.1±0.6 SD). Furthermore, similar results were reported in earlier treated studied groups after 24 hour (mean=4.4±0.7 SD). However, a decreased level in serum albumin concentrations was observed in 48 hour (mean=2.9±0.2 SD), up to one week (mean=3±0.4 SD) and on long-term of three weeks of treatment (mean=3±0.2 SD). The infected pre-treated mice showed lower serum concentrations of albumin (mean=3.5±0.3 SD) compared to normal control animals (mean=4.1±0.6 SD), see Table 1 and Fig. 1A.
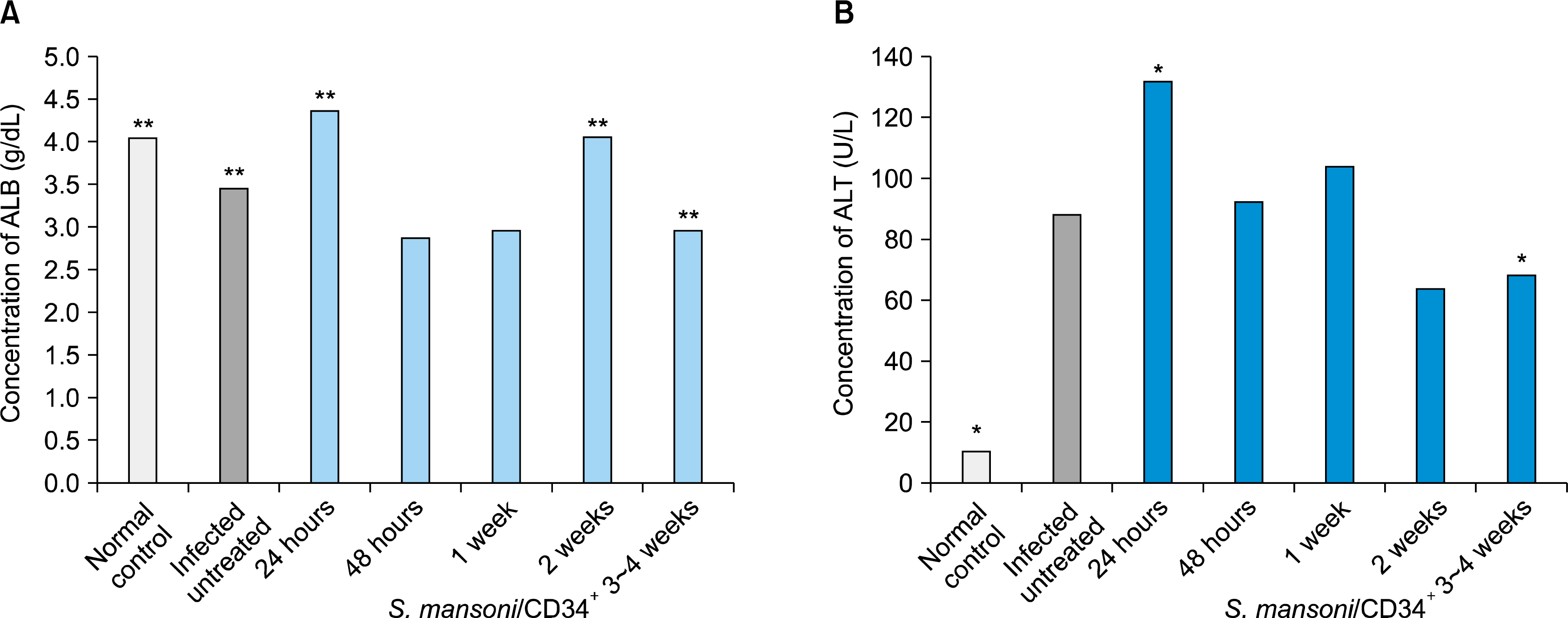 | Fig. 1.Changes in serum concentrations of albumin (A), and also the ALT values (B) in both normal and S. mansoni-infected control groups versus infected mice groups followed by treatment of CD34+ cells for 24 hour, 48 hour, 1 week, 2 weeks and 3∼4 weeks. *p-value <0.05 and **p-value <0.05.
|
Table 1.
Serum albumin (g/dL) and ALT (U/L) levels in the normal (−ve controls), S. mansoni-infected (+ve controls) and post-treated (1×105 hUCB-selected CD34+ cells/mouse) experimental mice groups
| Parameter of Interest | −ve Controls (n=6) | +ve Controls (n=8) | Studied Groups | ||||
|---|---|---|---|---|---|---|---|
|
|
|||||||
| 24 hour (n=6) | 48 hour (n=6) | 1 week (n=6) | 2 weeks (n=6) | 3∼4 weeks (n=9) | |||
| Albumin (g/dL) | 4.1**±0.6 | 3.5**±0.3 | 4.4**±0.7 | 2.9±0.2 | 3±0.4 | 4.1**±0.2 | 3**±0.2 |
| ALT (U/L) | 11*±3.5 | 88.4±35.9 | 132*±63.4 | 92.5±64.2 | 104.3±47.4 | 64±21.5 | 68.7*±10.5 |
![]()
Assessment of changes of the liver enzyme serum ALT (U/L) post-treatment was performed on 24 and 48 hours (measured as mean serum level of ALT±SD=132±63.4 and 92.5±64.2 U/L) as well as changes on week 1, 2 and up to 3∼4 weeks (mean±SD=104.3±47.4, 64±21.5 and 68.7±10.5 U/L), respectively. We also noted a significant clear evidence that the elevated serum ALT (U/L) levels among the S. mansoni-infected pretreated mice control (mean=88.4±35.9 SD) was significantly reduced at the 2 weeks of the treated study groups (mean=64±21.5 SD) and retained such decreases up to three to four weeks (mean=68.7±10.5 SD) after CD34+ cells treatment. All mice of a normal non-infected control with a mean 11 ±3.5 SD of serum concentrations of ALT (U/L) levels were also reported (Table 1 and Fig. 1B).
Fecal egg counts and eggs per 0.1 gram (EPG)
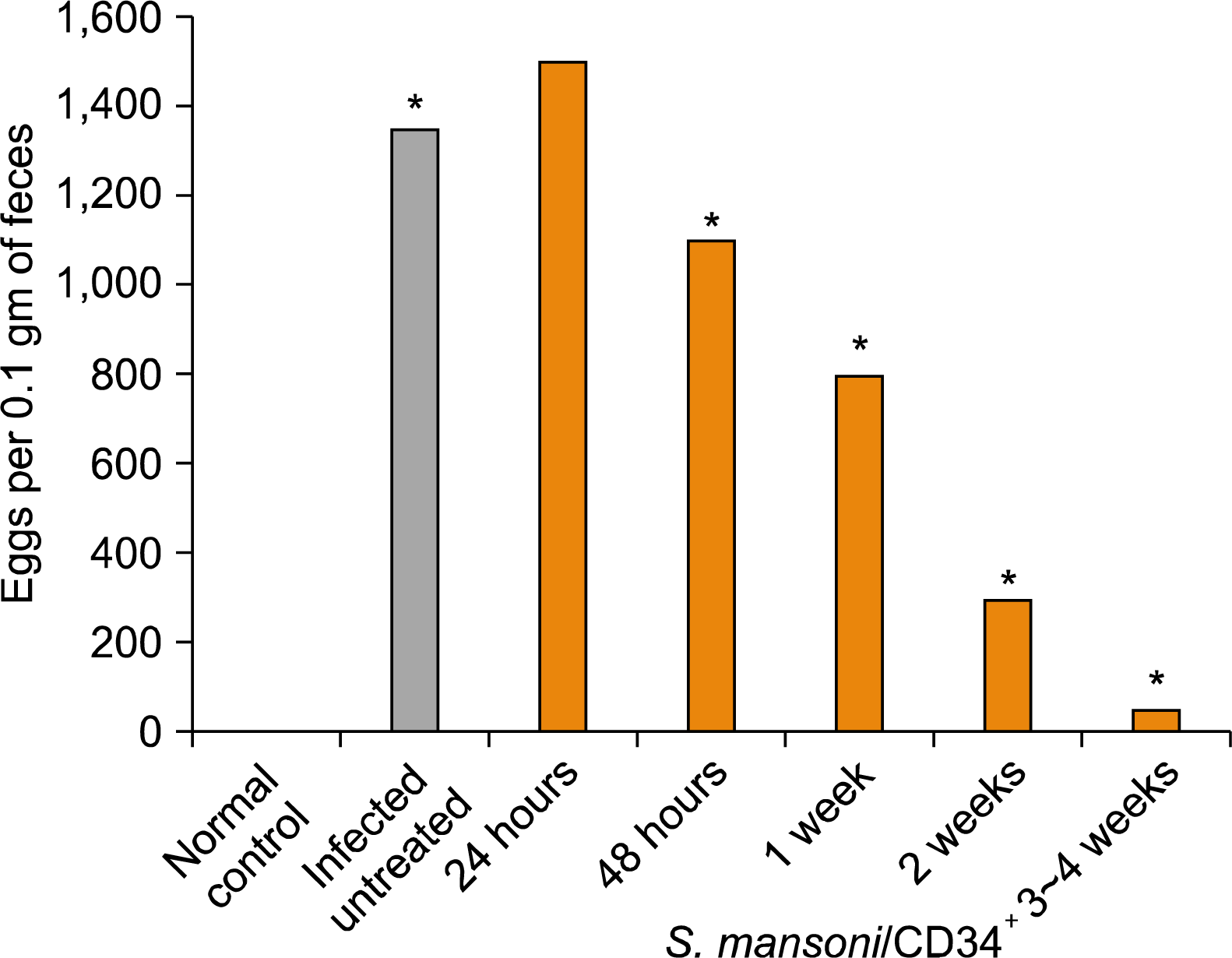
In general, the high level of fecal egg counts in S. mansoni-infected control (mean=1350±71 SD EPG) is considered to be a good reflection of infection intensity. The number of egg counts/0.1 gram stool per day in post-treatment studied groups was performed on day 1 and 2 as well as on week 1, 2 and 3∼4 weeks (mean±SD=1500±71, 1100±141, 800±71, 300±35, 50±5 EPG), respectively, see Table 2.
Table 2.
Average fecal egg counts per 0.1 gram (EPG) in pre-treated S. mansoni-infected mice (+ve controls) and post-treated (1×105 hUCB-selected CD34+ cells/mouse) experimental groups
| Parameter of Interest | −ve Controls (n=6) | +ve Controls (n=8) | Studied Groups | ||||
|---|---|---|---|---|---|---|---|
|
|
|||||||
| 24 hour (n=6) | 48 hour (n=6) | 1 week (n=6) | 2 weeks (n=6) | 3∼4 weeks (n=9) | |||
| Av. EPG (eggs/0.1 g) | 0.0 | 1350*±71 | 1500±71 | 1100*±141 | 800*±71 | 300*±35 | 50*±5 |
![]()
It was found that EPG in infected untreated mice (mean=1350±71 SD EPG) was significantly reduced in the mean number at the 1 and 2 weeks of the treated experimental groups (mean±SD=800±71, 300±35 EPG) respectively, and retained such a lowest decline mean egg counts up to three weeks post-treatment (mean±SD=50± 5 EPG) by the end of the experiment. In the other treated experimental group of 48 hour, a reduction of arithmetic mean number of fecal EPG was also observed (mean= 1100±141 SD EPG). In contrast, the mean number of egg counts per studied group in 24 hour (mean=1500±71 SD EPG) after treatment was higher than that in the infected non-treated mice (mean=1350±71 SD EPG) and the differences between them were not significant (p>0.05). However, the EPG per studied experimental group after treatment was significantly decreased as the time of CD34+ cells treatment increased. At the end of fourth week post-treatment, the number of fecal eggs per 0.1 gram was significantly decreased (mean=50±5 SD EPG) as compared with that of the infected non-treated controls (p<0.05). Results of the egg counts per studied group are shown in Table 2 and Fig. 2.
Histopathological examination of the liver
We made histopathological slices of liver samples harvested from uninfected non-treated controls, S. mansoni-infected non-treated controls and CD34+ cells treated studied mice groups for shorter and longer time periods. S. mansoni infection is characterized by deposition of parasite eggs in the hepatic portal tracts of the host with granuloma formation as a result of cell-mediated immunity (CMI) reactions in response to soluble egg antigen (SEA) (17). This may progress to irreversible host liver fibrosis and consequently severe portal hypertension specific for liver affection in schistosomiasis due to the size of the schistosomal ova (50∼150 μm) (18).
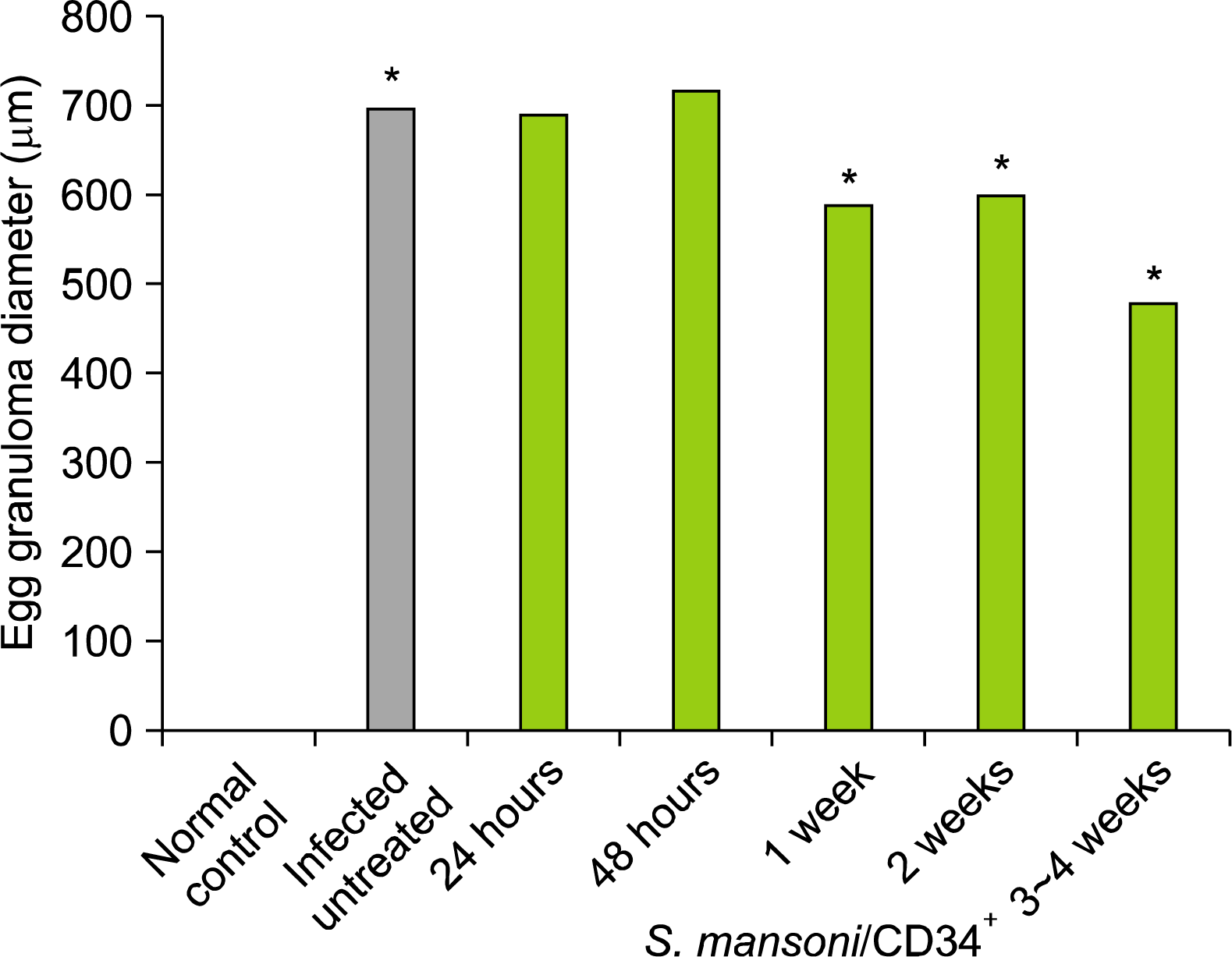
Granulomas in the liver, when present, is regarded as diagnostic feature of schistosomiasis and therefore given greater weight than other features. Moreover, the size of the anti-egg granuloma in the liver may vary with schistosomiasis infection intensity (19). The granulomas diameters (μm) in the liver of the 20 largest granulomas that appeared only circular in form per each studied experimental group were measured with an ocular micro-meter and calculated assuming a spherical shape as described by (19, 20), see Table 3.
Table 3.
Mean egg granuloma diameter (μm) in the liver of the S. mansoni-infected (+ve controls) versus infected post-treated experimental mice groups on 24 hour, 48 hour, 1 week, 2 weeks and 3∼4 weeks (n=20)
| Parameter of Interest | −ve Controls (n=6) | +ve Controls (n=8) | Studied Groups | ||||
|---|---|---|---|---|---|---|---|
|
|
|||||||
| 24 hour (n=6) | 48 hour (n=6) | 1 week (n=6) | 2 weeks (n=6) | 3∼4 weeks (n=9) | |||
| granuloma diameter (μm) | 0.0 | 700*±46 | 690±46 | 720±141 | 590*±39.3 | 600*±40 | 480*±32 |
![]()
Histopathological data after examination of the liver samples harvested from untreated S. mansoni-infected mice revealed the occurrence of parasite egg chronic numerous granulomas and host liver fibrosis (Fig. 4 panels a, b and c). Examination of the liver granulomas of CD34+ cells treated mice on day 1 and 2 as well as on week 1, 2 and 3∼4 weeks were lower in the number (data not shown) and smaller in the diameter than that in the S. mansoni-infected, non-treated mice. As shown in Table 3 and Fig. 3, the mean egg granuloma diameter was decreased in size two-fold between three and four weeks and did not change in size thereafter post treatment (480±32 SD μm), and was significantly smaller than that in the infected non-treated group (700±46 SD μm, a p-value of <0.05). These results demonstrate that hUCB-selected CD34+ cells can alleviate liver injury caused by the parasitic flatworm S. mansoni. The hepatic lobular architecture restored its normal organization and most hepatocytes showed normal appearance, even in the vicinity of the granuloma. However, some sinusoids were still dilated and infiltrated with lymphocytes, and some Kupffer-cells were still hypertrophied (Fig. 4 panel d).
 | Fig. 4.Photomicrographs of (H&E)-stained liver tissue sections from mice infected with S. mansoni. Panels a, b and c: Untreated control group of CD34+ cells. Panels a and b showing characteristic fibrous granulomas with trapped central eggs and sinusoidal dilation (original magnification 100× and 200× respectively). Panel c showing a large granuloma causing distortion of the hepatic lobule and accumulation of inflammatory cells around the disrupted central vein (original magnification 400×). Panel d showing a section of the liver tissue from mice infected with S. mansoni followed by treatment of CD34+ cells with a reduced egg granuloma diameter and the relief of ameliorated hepatocytes after three to four weeks of treatment (power of magnification 400×).
|
Go to :
Discussion
hUCB-derived CD34 positive stem and progenitor cells reduce serum ALT levels of S. mansoni-induced liver fibrosis and can restore the serum albumin liver secretion. The mouse model is susceptible to S. mansoni infections (21), and hence subsequent progression of liver pathology in the form of fibrosis (18) with a major end-stage liver failure. Concentrations of serum albumin secretion can be used to assess the synthetic specific function of the liver (22). Concentrations of serum ALT can be used to assess ongoing hepatocyte injury and fibrosis (23). The serum levels of ALT usually rise in cases of schistosomiasis, while low levels of liver-associated serum albumin secretions also indicate liver function decomposition stage in schistosomiasis-infections (24). In the present study, we found that elevated serum ALT (mean=88.4±35.9 SD U/L) above the upper limit of normal (mean=11±3.5 SD U/L) indicate a good predictor for development of liver fibrosis in the S. mansoni-infected mice control. We have then evaluated the efficacy of hUCB-derived CD34 positive cells on those with S. mansoni-infected control animals, and demonstrated that levels of liver-associated serum enzyme ALT was deceased gradually at the 2 weeks of the treated study groups (mean=64±21.5 SD) and retained such decreases up to three to four weeks (mean= 68.7±10.5 SD) after CD34+ cells treatment. There was a statistically significant between sampling time post-treatment and a decreased serum ALT, independent of the dose of treatment that mice were taking. We compared concentrations of serum albumin secretion (measured as mean serum levels of ALB g/dL) in the CD34+ cells treated mice groups in those with, and without, S. mansoni infection. The results of this study showed that the treatment with hUCB-selected CD34+ cells improved the levels of serum albumin concentrations (g/dL) and hence the residual liver functions in a time-dependent manner. We noted that the treated animal groups with CD34+ cells have a strong association with the improvement of the liver serum albumin concentrations at the 2 weeks of treatment (mean=4.1±0.2 SD g/dL) and showed no significant differences to normal control mice (mean=4.1±0.6 SD g/dL).
hUCB-selected CD34+ cells reduced the mean egg granuloma diameter and the number of granulomas in the livers of the S. mansoni-infected control animals. S. mansoni is characterized by formation of parasite egg granulomas and host liver fibrosis (25, 26) - by pathologists experienced in the interpretation of such liver histopathological features in infected humans and experimental animals (27). As expected typical S. mansoni egg granuloma formation was seen in livers of all infected mice group. When mice infected with S. mansoni were treated intravenously with CD34+ cells, mice developed much less lodged granuloma formation in the livers of mice of different experimental groups than control S. mansoni-infected non-treated group. The results of the histopathological data indicated a reduced egg granuloma number as well as smaller egg granuloma diameter between three and four weeks after treatment (on the average by 480±32 SD μm) as determined by analyses of histological liver sections of infected, CD34+ cells treated mice vs. infected, non-treated mice (on the average by 700±46 SD μm, a p-value of <0.05), suggesting that treatment with hUCB-selected CD34+ cells allow the host to alleviate liver damage caused by the worm eggs which is responsible for the granuloma formation, and hence a role to prevent hepatic granuloma formation in schistosomiasis. This would provide a basis for subsequent experimental investigations and clinical trials to pinpoint the interesting role of hUCB-derived CD34+ cells against complications of schistosomiasis pathology as a result of granuloma formation in the liver.
Quantitative estimation of the S. mansoni eggs counted in fecal samples of infected mice is reduced in a time dependent manner after hUCB-derived CD34+ cells treatment. The detection of changes of S. mansoni eggs in the feces had begun to decrease as the time of CD34+ cells treatment increased. At the end of fourth week post-treatment, the number of fecal eggs was minimal in infected post-treated mice (mean=50±5 SD EPG) as compared with that of the infected non-treated controls (mean=1350±71 SD EPG). We hypothesized that the substantial reduction of the parasite fecal eggs in infected, treated mice owing to mice positively responded to the CD34+ cells experimental conditions used which is not present in the infected non-treated mice. A correlation between diminished egg granulomas in the liver specimens and changes in the number of the eggs in the feces of S. mansoni-infected post-treated experimental mice groups on 24 hour, 48 hour, 1 week, 2 weeks and 3∼4 weeks was also apparent in a time dependent manner. Correspondingly, the egg production is positively influenced by CD34+ cells treatment. However, it is not clear if the parasite (not egg) stop to produce newly deposited eggs than at earlier times during infection or the inefficient passage of eggs into the host feces due to accumulating tissue-trapped eggs, which eventually die within the tissues of infected mice. This issue needs further exploration.
Effects of intravenous administration of hUCB-derived CD34+ cells after damaged liver-tissues by infection with the human parasitic flatworm S. mansoni not only alleviated liver injury and fibrosis in vivo but also prolonged the survival time of infected mice. Regarding liver regeneration, CD34+ cells can differentiate into hepatocytes in vitro (28) and in vivo (29); and transplantation of CD34+ cells can also suppress fibrotic changes in rat livers with established fibrosis or cirrhosis (30, 31). In exploring the efficacy of hUCB-selected CD34+ cells in S. mansoni-infected mice, it was found that the intravenous administration of isolated CD34+ cells have modestly reduced histological liver damage; reduced the increase in liver-associated serum levels of ALT; significantly improved the decrease in liver serum levels of albumin and reduce egg granuloma diameter in the livers of S. mansoni-infected mice especially at later time points post-treatment.
Our findings suggest that hUCB-derived CD34+ cells treatment might represent a novel potential therapeutic source for treatment of schistosomiasis in mice and be able to ameliorate S. mansoni-induced liver injury and fibrosis in vivo and this effect was enhanced for long time periods after three to four weeks of treatment. However, the efficacy and underlying mechanisms remain to be further characterized.
Go to :
 XML Download
XML Download