

 PDF
PDF Citation
Citation Print
Print


Introduction
Embryonic development and later rejuvenation of adult tissue is regulated by a population of stem cells that, by undergoing self-renewal maintain their own pool and give rise to differentiated progenitors that replace cells used up during the life cycle (1). Therefore, SCs are guardians of tissue/organ integrity and regulate the life span of an adult organism.
The most important SC population, from a regenerative point of view, is pluripotent stem cells (PSCs) (2). Such cells, according to their definition, must fulfill some in vitro as well as in vivo criteria, such as i) they give rise to cells from all three germ layers, i.e. the ectoderm, mesoderm, and endoderm; ii) complete blastocyst development; and iii) form teratomas after inoculation into experimental animals (3). During embryo development, pluripotent stem cells are established from the most primitive stem cells, i.e. fertilized zygote. These cells are called totipotent – and possess the ability to give rise to both embryo and placenta. Totipotent zygotes further develop into PSCs in the inner cell mass (ICM) of blastocysts and may give rise to all three germ layers of a developing embryo, however, they lose the ability to differentiate into placenta. The PSCs in this stage could be expanded ex vivo as embryonic stem cell (ESCs) lines (4, 5).
The pluripotency is inherited from fertilized zygote, cells from inner-cell mass in blastocysts, to epiblasts from primitive ectoderm after implantation (6). After gastrulation, almost all cells lose their pluripotency and are committed into specialized lineage. However, primordial germ cells (PGCs) derived from proximal epiblast cells, germ-line stem cells maintain their pluripotency by expressing pluripotency-specific genes, such as Oct4 and Nanog (7). The PGCs also lose their pluripotency and initiate their commitment to gametes, i.e. oocytes and sperm cells, after migration into the genital ridge. Therefore, the PSCs could be specific population detected during only the narrow developmental period of early embryogenesis. Indeed, embryonic stem cells (ESCs), epiblast-derived stem cells (EpiSCs), and embryonic germ cells (EGCs) are established from these early embryonic PSCs using specialized in-vitro cell cultures (3).
As previously mentioned, PSCs are detected only during very early embryonic development and they disappear in adulthood as they differentiate into terminally differentiated monopotent somatic or germ-line cells (2). However, several attempts have been made in the past few years to purify a population of pluripotent stem cells (PSCs) from adult tissue. Potential PSCs in adult tissue were described as i) multipotent adult progenitor cells (MAPCs) (8), ii) multipotent adult stem-cells (MASCs) (9, 10), iii) unrestricted somatic stem-cells (USSCs) (11), iv) marrow-isolated adult multilineage-inducible (MIAMI) cells (12), and v) multilineage-differentiating, stress-enduring stem (Muse) cells (13). It is conceivable that all of these cells may be closely related and some stem cell described by different investigators may be overlapped.
Our group recently isolated a population of pluripotent, very small, embryonic-like stem cells (VSELs) from adult, murine bone marrow (BM) (14), fetal livers (15), several adult-murine organs (16), and also from human cord blood (17). VSELs express many morphological, e.g. relatively large nuclei containing euchromatin, and molecular, e.g. expression of SSEA-1, Oct4, and Nanog, markers characteristic for ESCs (14). We hypothesize that VSELs are deposited during early gastrulation in developing tissues/organs, survive into adulthood, and have an important role as a back-up population of PSCs in the turnover of tissue-committed stem cells (TCSCs). In this review article, we will discuss in detail the molecular nature of the Oct4+ VSELs.
Go to : 
Very small, embryonic-like stem cells (VSELs) obtained from adult tissue
VSELs were isolated using the multiparameter fluorescence-activated cell sorter (FACS) as a population of Sca-1+Lin−CD45− in several other adult murine organs, e.g. brain, liver, skeletal muscles, heart, and kidney, and as a population of CD133+CXCR4+Lin−CD45− in human cord blood and peripheral blood (14, 16, 17). These very small sized (∼3–6 μm) cells express pluripotent markers such as Oct4, Nanog, and SSEA-1 (mouse) or SSEA-4 (human). Morphologically, VSELs similarly to ESCs possess large nuclei containing unorganized chromatin (euchromatin) and exhibit a significantly higher nuclear/cytoplasm (N/C) ratio and a lower cytoplasmic area compared to HSCs (16).
When freshly isolated VSELs are cultured under a C2C12 myoblast feeder layer, they form spheres that correspond to embryoid bodies (EBs) from ESCs (18). The VSEL-derived spheres (VSEL-DSs) contain primitive SCs that, after replating into specific, differentiation-inducing media differentiate into cells from all three germ layers. Furthermore, we reported that VSELs are mobilized into peripheral blood during several models of organ injury and circulate there in an attempt to enrich and regenerate damaged tissue, e.g. heart infarct and stroke (19–24). This physiological mechanism undoubtedly has a more significant role in the regeneration of some small tissue and organ injuries.
Go to :
Molecular signature of VSELs residing in BM
To investigate the relationship between VSELs and the embryonic type of PSCs, e.g. ESCs, EpiSCs, PGCs, and EGs, we used several molecular strategies to evaluate the molecular signature of VSELs. Highly purified Sca-1+Lin−CD45− VSELs from murine BM were evaluated for expression of i) pluripotent genes, ii) epiblast/germ line markers, and iii) expression of developmentally crucial imprinted genes (Fig. 1).
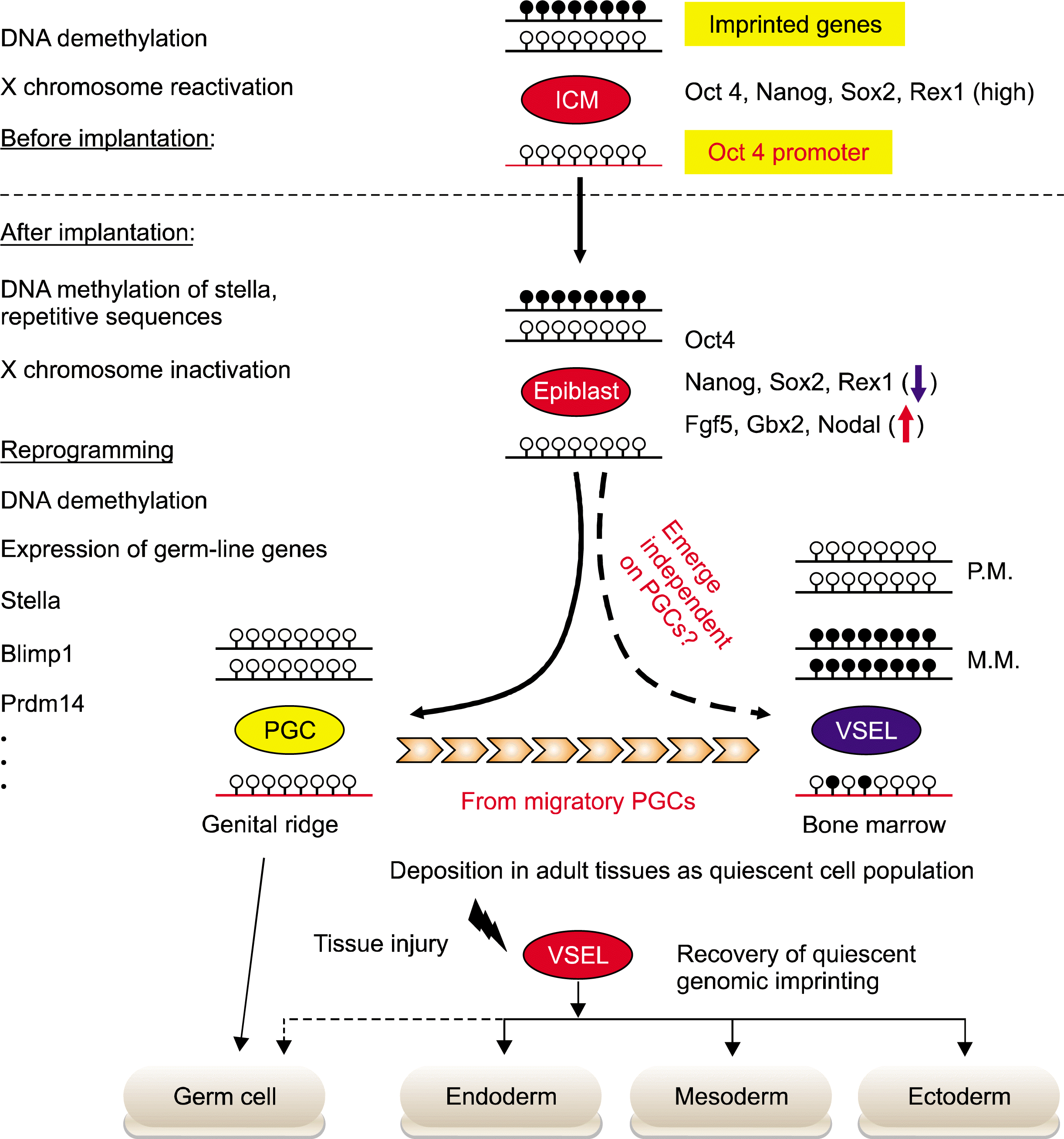 | Fig. 1.Epigenetic modification of VSELs during embryogenesis and tissue regeneration. Epigenetic modifications control the differentiation potential of stem cells during embryogenesis and tissue regeneration. During the implantation of embryo, ICM-derived epiblast stem cells methylated again i) X chromosome, ii) promoters for the genes characteristic for PSCs in ICM (Rex-1 and Stella), and iii) repetitive sequences. However, at the beginning of gastrulation the proximal, epiblast-specified PGCs can reset their epigenetic profile to one that characterizes ICM-derived PSCs. Subsequently, during PGCs migration to genital ridges, the global DNA demethylation leads to erase the genomic imprints. As it is hypothesized that VSELs originate from the epiblast-derived PGCs population, they show the PGCs-like epigenetic profiles, including the partial DNA demethylation in the regulatory DNA elements of several pluripotency, germ-line genes, and genomic imprints. The epigenetic profiles of developing VSELs are retained after their deposition into adult tissue. This parental-specific reprogramming of genomic imprinting of VSELs deposited in adult -tissue, e.g. BM, functions as i) a “lock-in mechanism” to prevent their unleashed proliferation and ii) a mechanism to restrict their sensitivity to Ins/Igf signaling. After exposure to tissue injury, quiescent VSELs de-repress “locked-in” genomic imprints along with progressive methylation of DNA in the Oct4 promoter. As a result of these epigenetic changes, VSELs become involved in the tissue regeneration process by differentiation into cells of all three germ layers, i.e. meso-, ecto-, and endoderm.
|
VSELs express PSCs genes
Unlike differentiated somatic cells, PSCs commonly express pluripotency core transcription factors (TFs), such as Oct4, Nanog, and Sox2. These TFs form the pluripotent core circuitry by reinforcing the expression of genes, and are involved in maintaining the PSC undifferentiated status but repress the differentiation inducing transcription (25). The significance of these core factors is experimentally demonstrated by the inducible pluripotent stem cells (iPSCs) protocol in which fully differentiated somatic cells can be reprogrammed into ESC-like stem cells by transduction of the so-called Yamanaka factors (Oct4, Sox20, Klf4, and cMyc) (26, 27). We noted that murine VSELs express both the Oct4 transcript and protein (28). However, a few recent reports cast doubts regarding the true expression of this important PSCs marker in cells isolated from adult tissue, especially, as postulating the expression of several Oct4 pseudogenes can produce false-positive RT-PCR results (29, 30). Therefore, we evaluated in murine the epigenetic status for the Oct4 promoter, which are the most convincing in vitro criteria for the evaluation of putative stem cells. When the DNA methylation status of the Oct4 promoter was examined by bisulfate sequencing using the highly purified Sca-1+Lin−CD45− VSELs, the Oct4 promoter in VSELs, similar to that in cells isolated from ESCs-derived EBs, was hypomethylated (28% and 13.2%, respectively) (28). With the Carrier chromatinimmunoprecipitation (ChIP) assay using human hematopoietic cell-line THP-1 as the carrier (31), we found that the Oct4 promoter chromatin in murine VSEL is enriched with H3Ac, an open chromatin histone code, but was less associated with H3K9me2, a heterochromatin histone code (28). For determining the epigenetic status for the Nanog promoter, it was methylated (∼50%) at the DNA level, however, quantitative ChIP data confirmed that the H3Ac/H3K9me2 ratio supports the active status of the Nanog promoter in these cells (28). Based on these results, VSEL exhibits the open chromatin structure in the promoter of the Oct4 and the Nanog in order to support the true expression of these genes. Of note, we also reported that VSELs express also several other markers of PSCs, such as SSEA-1 antigen as well as Sox2 and Klf4 transcription factors.
Expression of germ-line markers
After gastrulation, most embryonic cells lose their expression of pluripotency-specific TFs except for some of the SC population from proximal epiblasts, which are precursors for PGCs development. The PGCs derived from epiblast reset epigenetic marks to ICM-like status, resulting in re-activation of pluripotency and germ-line-related genes (6). The PGCs are epiblast-derived, alkaline phosphatase (AP)-positive, migratory-stem cells. They emerge in extraembryonic mesorderm at the base of the allantois, an appendage arising from the posterior, primitive streak around E7.25 (32). At this time, approximately 40 cells are detected. The specification of PGCs is initiated by the expression of germ-line master regulators, such as Fragilis, Blimp1, and Stella, in response to signals from extra-embryonic tissues (33). At E8.5, PGCs enter back into the primitive streak region of the embryo and begin migration through hindgut endoderm and mesentry in aorta-gonads-mesonephros (AGM), and reach the genital ridge around E11.5 to E12.5. During this period, there is active proliferation, leading to ∼25,000 PGCs at 13 dpc. After E13.0 (post-migration), PGCs i) cease proliferation, ii) change in cell morphology and cell-adhesion molecules, iii) lose cell migratory capacity, and iv) initiate gametogenesis. During migration, PGCs initiate epigenetic reprogramming, such as erasure of genomic imprinting, DNA demethylation, and X chromosome reactivation and complete them after colonizing at the genital ridge. This epigenetic reprogramming could be responsible for the proper expression timing of various germ-line genes, such as Blimp1, Stella, Mvh, Dazl, and Sycp3 (7). Therefore, early emerging, migratory, and post-migratory PGCs show distinguished characteristics regarding the epigenetic status and gene expression profile.
Importantly, VSELs highly express the genes involved in the germ-line specification of the epiblast, e.g. Stella, Prdm14, Fragilis, Blimp1, Nanos3, and Dnd1. Subsequently, the expression of Stella, Blimp1, and Mvh protein was confirmed by immunostaining (34). The Stella promoter is partially demethylated regarding DNA methylation. In addition, this promoter was highly enriched with transcriptionally active histone modifications (H3Ac and H3K4me3), but was less enriched for transcriptionally repressive histone markers (H3K9me2 and H3K27me3) (34). Taken together, VSELs express specific genes and display a Stella promoter chromatin structure that is characteristics for germ-line specification. Interestingly, late migratory PGCs markers including Dppa2, Dppa4, and Mvh are highly expressed in VSELs, however, these cells do not express Sycp3, Dazl, and LINE1 genes that are highly expressed in post-migratory PGCs (34). Thus, our results in toto support the concept that VSELs deposited into murine BM show some similarities in gene expression and epigenetic signatures to epiblast-derived migratory PGCs (∼E10.5–E11.5).
Presence of bivalent domain in murine VSELs
Owing to their limited number in adult tissue, it has been difficult to gain a better insight regarding the molecular nature of VSELs in detail. We recently established a cDNA library from 20 cells of FACS-sorted VSELs, hematopoietic stem cells (HSCs) or a cultured embryonic stem-cell line, ESC-D3 (35). By performing single-cell-based, genome-wide gene expression analysis (36), we found that VSELs show similar, but characteristic transcriptome compared with ESCs and HSCs (35). One gene of particular interest which is highly expressed in VSELs is Ezh2, a polycomb group protein. Ezh2 has an essential role for maintaining the bivalent domain (BD) epigenetic marker. In undifferentiated PSC, the promoter regions of some developmentally crucial transcription factors are epigenetically marked both the transcriptionally active H3K4me3 codes and the repressive histone ones such as trimethylated on lysine27 of histone3 (H3K27me3) and this epigenetic modification is called bivalent domain. The bivalent domain is detected mainly in the promoter region for the homeodomain containing development master TFs, such as Dlx, Irx, Lhx, Pou, Pax, and Six family proteins (37–39). Due to the overwhelming effect of transcription-repressive activity endowed from H3K27me3, the transcription of bivalent domain target genes is transiently re-pressed in order to prevent the premature expression of the differentiation inducing TFs. However, in response to the development stimuli, the bivalent domain of these promoters is switched into a monovalent domain in order to determine their gene expression. By conducting differentiation and RNA interference experiments, we demonstrate that Ezh2 in VSELs has a similar role for preventing premature activation of lineage-committing transcription factors (35). Taken together, therefore, both positive (expression of Oct4-Nanog-Sox2 pluripotency circuitry) and negative (repression of differentiation inducing TFs by bivalent domain) regulatory mechanisms are indispensable in order to maintain the pluripotency of VSELs.
Go to :
Epigenetic changes of imprinted genes that regulate VSELs pluripotency
Unlike ESCs, we noticed that murine Oct4+ VSELs do not proliferate in vitro if cultured alone and do not grow teratomas in vivo. On the other hand, cells isolated from VSEL-DSs regain their proliferation potential, thus suggesting that VSELs are a quiescent cell population and that some mechanisms must exist to prevent their proliferation in a normal physiological condition. Similarly to VSELs, PGCs in cultures freshly isolated from embryos proliferate for a few days only and then disappear either because they differentiate or die (14). However, when PGCs are cultured over the feeder layer and with appropriate growth factors cocktails, they restore the proliferation potency and can be established into EG cells. Therefore, it is possible that two stem-cell populations could employ similar molecular mechanisms to regulate their pluripotency and to prevent unleashed cell proliferation.
The hallmark for epigenetic reprogramming during PGC development is erasure of genomic imprinting. Genomic imprinting is an epigenetic process ensuring paternal-specific, mono-allelic expression of so-called imprinted genes. Approximately 80 imprinted-genes (expressed from maternal or paternal chromosomes only) have been reported in mouse genome and their proper mono-allelic expression regulates totipotency and pluripotency of the zygote and developmentally early stem cells, respectively (40). Furthermore, most imprinted genes, such as insulin-like growth factor 2 (Igf2), H19, Igf2 receptor (Igf2R), and p57KIP2 (also known as Cdkn1c), are directly involved in embryo growth and development. As the majority of imprinted-genes exist as gene clusters enriched for CpG islands, their expression is coordinately regulated by the DNA methylation status on CpG-rich cis-elements known as differently methylated regions (DMRs) (41). The differential methylation status of DMRs is mediated by DNA methyltransferases (Dnmts), depending upon the parental allele origin. Depending on the developmental period of methylation, there are two types of DMRs, i.e. “primary DMRs” which are differentially methylated during gametogenesis, and “secondary DMRs” which acquire allelic specific methylation after fertilization. To date, 15 primary DMRs have been identified in mouse genome. Interestingly, most primary DMRs are maternally methylated ones and only three DMRs, i.e. Igf2-H19, Rasgrf1, and Meg3 loci, are paternally methylated (42). In addition to DNA methylation on DMR, histone modifications also contribute to the monoallelic expression of the imprinted genes.
Shortly after PGCs specification at E7.25, PGCs initiate the epigenetic reprogramming programs resulting in progressive global DNA demethylation and changes in histone modifications (43). As a result, DNA methylation marks for genomic imprinting in both parental chromosomes are erased during PGCs migration into the genital ridge and new genomic imprints are established during terminally maturation into gametes in the sex-dependent manner. The erasure of genomic imprints could be a potential mechanism used to restrict the Oct4 expressing germ-line stem cells from unleashed cell proliferation and teratoma formation. For example, while the nuclei of early migrating PGC at E8.5–9.5 can be successfully used as donors for nuclear transfer, nuclei from post-migratory PGC after E11.5 are unable to support full-term development (44).
Since VSELs exhibit a similar molecular signature to PGCs, we have hypothesized that VSELs might employ the epigenetic reprogramming of genomic imprinting to prevent them from unleashed proliferation and this mechanism may be responsible for maintaining their quiescent status in adult tissue. Indeed, VSELs freshly isolated from murine BM erase the paternally methylated imprints, e.g. Igf2-H19 and Rasgrf1 loci, however, at the same time they hypermethylate the maternally methylated ones, e.g. Igf2 receptor (Igf2R), Kcnq1-p57KIP2, and Peg1 loci (28, 34). Because paternally expressed, imprinted-genes (Igf2 and Rasgrf1) enhance the embryo growth and maternally expressed genes (H19, p57KIP2, and Igf2R) inhibit cell proliferation (28, 34), the unique genomic imprinting pattern observed on murine VSELs demonstrates that the pattern of genomic imprints in these cells is favorable to the repression of cell proliferation. Accordingly, VSELs highly express growth-repressive imprinted gene transcripts, H19, p57KIP2, and Igf2R, and down-regulate growth-promoting ones, Igf2 and Rasgrf1, and which explains the quiescent status of VSELs (28). Importantly, cells isolated from VSEL-DSs which proliferate and differentiate progressively recover all of the growth repressive patterns of genomic imprinting. These results suggest that epigenetic reprogramming of genomic imprinting should be primary mechanism for maintaining the quiescence of the most primitive pluripotent adult stem cells, e.g. Oct4+ VSELs, deposited in the adult body and protect them from premature aging and tumor formation (28). Indeed, we demonstrated that the epigenetic changes of some imprinted genes, Igf2-H19, Igf2R, and Rasgrf1 loci regulate the responsiveness to the insulin factor signaling pathway in murine VSELs. It is well-known that the attenuation of the insulin/insulin-like growth factor (Ins/Igf) signaling that positively correlates with longevity (45). Thus, our experimental data using several animal model mice for longevity and Ins/Igf signaling demonstrate that the pool and regeneration potency of VSELs deposited in adult tissue gradually decrease throughout life in an Ins/Igf signaling-dependent manner and the ageing associated dysfunction of VSELs could be an important mechanism of aging (46–48). Therefore, the investigation of potential modulation of mechanisms controlling genomic imprinting in VSELs is crucial for developing more powerful strategies to unleash the regenerative potential of these cells for efficient use in the clinical setting.
To get a better molecular insight about the quiescent state of VSELs, we recently performed the genome-wide analysis of murine BM-derived VSELs. Accordingly, the attenuation of mitogenic growth-factor signaling pathways also has a crucial role in their quiescence and ageing (49). Of particular interest, VSELs down-regulate genes involved in the UV radiation response, mRNA processing, and mitogenic growth factor signaling, e.g. from Igf-1 and TRKA receptors as well as for the ERK and PI3K pathways. Using leading-edge, subset analysis and real-time, quantitative PCR assays, we observed that several genes, such as Grb2, Sos1, Shc1, Map2k1, Akt3, Elk1, Rps6ka3, Gsk3β, and Csnk2a1, which are involved in mitogenic growth-factor signaling pathways, were commonly down-regulated in VSELs. In contrast, we observed that Oct4+ VSELs up-regulates tissue-specific gene sets and a gene set encoding the complement-coagulation cascade. Thus, we are investigating the role of these putative molecular targets for regulating the quiescence and ageing of VSELs.
Go to :
Conclusions
Several attempts have been made in the past few years to purify a population of PSCs from adult tissue. The very primitive VSELs in adult tissue could function as a precursor for monopotent TCSCs. Pluripotency of murine VSELs is already supported by their i) ability in vitro to differentiate in co-culture systems into three germ line-ages, ii) the presence of primitive, unorganized euchromatin in nuclei, iii) “true expression” of Oct4 and Nanog supported by demonstration of the hypomethylated promoter status and its association with acetylated histones, and iv) the presence of bivalent domains on homeodomain-containing, developmental transcription factors. However, these cells do not grow teratomas and are ineffective in blastocyst complementation assay due to modulation of methylation on regulatory sequences in developmentally important, imprinted genes, similarly seen in PGCs, to prevent them from unleashed cell proliferation and teratoma formation. Furthermore, VSELs could be specified into several tissue residing TCSCs, e.g. MSCs, HSCs, and cardiac stem cells, in response to tissue/organ injury. Taken together, it is very likely that these cells have a physiological role in the rejuvenation of a pool of TCSCs under steady state conditions. VSELs developmentally originate from epiblast-derived, migrating PGCs and they could be deposited in adult organs early in the development as a reserve pool of primitive stem cells for tissue repair and regeneration. Therefore, VSELs isolated from adult tissue are an alternative and not ethically controversial source of stem cells for regenerative medicine. To successfully use VSELs in the realm of regenerative medicine, it is very important to establish experimental protocols for the reprogramming of growth-repressive, genomic imprinting status in VSELs into a somatic pattern in order to unleash their regenerative potential.
Go to :
 XML Download
XML Download