

 PDF
PDF Citation
Citation Print
Print


Introduction
Mesenchymal Stem Cells (MSCs) are undifferentiated multipotent stem cells residing mainly in the bone marrow but also in different adult tissues (1–4), with the function of replacing dying cells and preserving tissue homeostasis. Under appropriate culture conditions, MSCs are able to differentiate in vitro into mesengenic lineages (osteogenic, chondrogenic and adipogenic), but also to transdifferentiate into cells of nonmesodermal origin such as hepatocytes (5) and neural cells (6).
Recently, MSCs have been proposed as offering a promising therapeutic approach in regenerative medicine and tissue engineering because of their favorable biological characteristics: 1) MSCs can be easily isolated and expanded in vitro (7); 2) after in vivo administration MSCs can migrate to the sites of injury (8); 3) MSCs can be used autologously and their hypo-immunogenicity makes them suitable also for allogenic transplantation (9). Furthermore, other mechanisms could contribute to the therapeutic benefits of MSCs such as their immunoregulatory properties and their capacity to secrete a broad spectrum of bioactive macromolecules (10). Several preclinical and clinical studies have confirmed the therapeutic potential of MSCs and a number of phase I/II and III clinical trials have recently been completed or are underway (please see http://clinicaltrials.gov). However, literature on MSCs often reported controversial results, partly due to different culture conditions, but surely also due to the intrinsic variability existing both between MSCs derived from different species, and also among donors/strains of the same species.
The different behavior between MSCs of human and non-human origin may be an hurdle, since animal models are used as first approach to study the feasibility of MSCs for treatment of human diseases. In particular, several studies have set up isolation and differentiation protocols to allow the use of rodent MSCs into transgenic animal models (11, 12).
Gene expression profiling of rodent bone marrow-derived MSCs compared with hMSCs has revealed a high degree of concordance (13, 14), but in spite of the shared expression of some common surface antigens such as CD29, CD90, CD105, used for MSC characterization, rat MSCs (rMSCs) and human MSCs (hMSCs) essentially differ for what concerns the genomic stability (15, 16), the spontaneous expression of neural markers (17, 18), and the immunoregolatory capacities (19), and therefore the translation of the results has not a foregone conclusion, since the biological analogies between different species are not sufficient to directly shift the results obtained on animal MSCs to hMSCs.
In this paper we examined and compared the in vitro ability of MSCs isolated from human and rat bone marrow and cultured in their standard conditions to differentiate into mesengenic (osteogenic, adipogenic and chondrogenic) lineages, one of the most promising fields for MSCs’ therapeutic application, since a more extensive knowledge of the biological properties of MSCs derived from different species would give a clue to handle the results obtained in the different studies, and to better apply it in clinic.
Go to : 
Materials and Methods
When not otherwise indicated, materials were purchased from Sigma Co, Saint Louis, MO, USA.
MSCs’ isolation and culture
rMSCs (p4–8) were collected according to a previously published protocol (20) and all animal procedures were conducted in accordance with the European Communities Council Directive 86/609/EEC. In brief, bone marrow was flushed from the femurs and tibias of 10-week old Sprague Dawley rats (n=5; Harlan, Udine, Italy) and red blood cells were lysed. The remaining cells were plated in 75 cm2 culture flasks in culture medium, consisting of α-MEM (Lonza Group Ltd., Basel, Switzerland) supplemented with 2 mM L-glutamine, 100 U/ml penicillin, 100 μg/ml streptomycin, 250 μg/ml fungizone and with 20% Fetal Bovine Serum (FBS, Hyclone, Logan, UT). All supplements were purchased from Lonza.
hMSCs (p4–8) were isolated after patients’ understanding and informed consent from aliquots of heparinized bone marrow obtained in excess from individuals undergoing marrow harvest for allogenic transplantation at the San Gerardo Hospital (n=5; Monza, Italy). In brief, mononuclear cells were collected after centrifugation in a Ficoll-Hypaque gradient and seeded 160.000 cells/cm2 into culture flasks in culture medium, consisting of DMEM (Lonza) supplemented with 2 mM L-glutamine, 100 U/ml penicillin, 100 μg/ml streptomycin, 250 μg/ml fungizone and with 10% FBS.
rMSC and hMSC cultures were maintained at 37°C in a humidified atmosphere containing 5% CO2. After 48 hours the non-adherent cells were removed and, subsequently, the medium was changed every 3∼4 days. When rMSC and hMSC cultures respectively reached confluence, cells were detached by trypsinization and used in experiments.
MSC differentiation
hMSCs and rMSCs were analyzed for their capacity to differentiate towards osteogenic, adipogenic and chondrogenic lineages using specific protocols. hMSCs and rMSCs cultured in medium without any differentiating agent were used as a control.
Osteogenic differentiation
Cells were seeded at approximately 3,500∼4,000 cells/cm2 onto culture dishes in culture medium, α-MEM (for rMSCs) or DMEM (for hMSCs), and supplemented with 10% FBS until subconfluence occurred. Afterwards, cells were grown in culture medium alone (control cells) or in osteogenic medium (OS cells) consisting of culture medium supplemented with 100 nM dexamethasone, 10 mM β-glycerophosphate and 0.05 mM ascorbic-2-phosphate acid. The osteogenic differentiation was evaluated by Alizarin Red S staining. At pre-established times, rMSC (7, 14, 21,28 days) and hMSC (7, 14, 21, 28, 35, 42 days) cultures were fixed with 4% paraformaldehyde for 10 minutes. Then cells were incubated for 30 minutes at room temperature in 1% Alizarin Red S and 1% ammonium hydroxide. Following incubation, cultures were washed twice with water and air-dried. Alizarin Red S dye binds to calcium ions present in mineralized deposits resulting in a brilliant red staining. To perform a quantitative analysis, Alizarin Red S dye was solubilized with a solution of 5% SDS in 0.5M HCl and the optical density was measured with a spectrophotometer at 425 nm.
Adipogenic differentiation
rMSCs and hMSCs were seeded at approximately 15,000∼20,000 cells/cm2 onto dishes in culture medium (respectively α-MEM or DMEM) supplemented with 10% FBS. After 24 hours cells were induced by treatment with “Adipogenic Induction Medium” (AIM) consisting of 4.5 g/L glucose culture medium supplemented with 10% FBS and adipogenic supplements (10 μg/ml insulin, 500 μM isobutylmethylxanthine, 100 μM indomethacin and 1 μM dexamethasone). When lipid droplets were observed throughout the cell culture, the medium was switched to “Adipogenic Maintainance Medium” (AMM), consisting of 4.5 g/L glucose culture medium plus 10% FBS supplemented with 10 μg/ml insulin. Adipogenic differentiation was evaluated by Oil Red O staining. At pre-established times, rMSC (7, 14, 21, 28, 35 days) and hMSC (7, 14, 21, 28, 35, 42 days) cultures were fixed with 4% paraformaldehyde for 10 minutes. Then, cells were incubated in 60% isopropanol for 5 minutes at room temperature and stained with 1.8% Oil Red O in 60% isopropanol for 15 minutes. Following incubation, cultures were washed twice with water and photographed. Oil Red O binds to lipids, in particular to triglycerides, resulting in a red staining.
To perform a quantitative analysis, Oil Red O was solubilized with 100% isopropanol and the optical density was measured with a spectrophotometer at 516 nm.
Statistical Analysis
Experiments for the quantitative assessment of osteogenic and adipogenic differentiation were performed three times in cells from different donors. The results were normalized to the absorbance of untreated control cells and they are expressed as mean±Standard Deviation. Statistical analysis was performed using the one-way ANOVA test and Tukey's multiple-comparison test as a post-test with the Graph Pad Prism statistical package (GraphPad Software, San Diego, CA, USA).
Chondrogenic differentiation
For chondrogenic differentiation, cells were grown into a pellet in 15 ml tubes, at approximately 250,000 cells/tube, in culture medium (control cells) or in chondrogenic medium for 7 weeks. rMSCs’ chondrogenic medium consisted of culture medium supplemented with 15% FBS, 1 mM piruvate, 100 nM dexamethasone, 37.5 μg/ml ascorbic-2-phosphate acid and 10 ng/ml Transforming Growth Factor-β3 (TGF-β3, Peprotech, Rocky Hill, NJ, USA). hMSCs’ “chondrogenic medium” consisted of 4.5 g/L glucose culture medium serum-free and supplemented with ITS+premix (1:100, Becton Dickinson, Franklin Lakes, NJ USA), 1 mM piruvate, 100 nM dexamethasone, 50 μg/ml ascorbic acid 2-phosphate, 10 ng/ml TGF-β3. Chondrogenic differentiation was evaluated by Safranin O staining. hMSC and rMSC pellets, treated with chondrogenic medium or with culture medium only, were fixed with 10% neutral buffered formalin for 30 minutes, paraffin embedded by standard methods and cut into 7 μm sections. Sections were stained with Hematoxylin-eosin and Safranin O 0.1%. The slides were mounted and photos were taken using a Nikon Coolscope instrument (Nikon Instruments S.p.A., Italy). Safranin O is a histological cartilage-specific staining that binds to proteoglycans and glycosaminoglycans resulting in a red/orange staining.
Go to :
Results
MSCs’ characterization
In accordance with the Mesenchymal and Tissue Stem Cell Committee of the International Society for Cellular Therapy (21), and as previously demonstrated (15, 16, 22), the MSCs isolated from rat and human bone marrow used in our experiments, were: a) plastic-adherent and capable of extensive proliferation when maintained in their standard culture conditions; b) positive for several antigens such as CD29, CD90, CD105, CD73, and lacking in the expression of hematopoietic surface molecules CD34, CD45; c) able to differentiate into osteogenic, adipogenic and chondrogenic lineages under specific in vitro differentiating conditions. The different abilities of MSCs of human and rat origin to differentiate into the three mesengenic lineages was compared.
Osteogenic differentiation
In rMSC cultures the Alizarin Red staining evidenced the presence of mineralized calcium only in osteogenic medium-treated cells after 14 days of treatment, and the intensity of the staining increased after 21 and 28 days of induction (Fig. 1A). These results were confirmed by the quantitative analysis of osteogenic differentiation, which evidenced a statistically significant increase in mineralized matrix deposition starting from 14 days of culture (Fig. 1B). On the contrary, untreated control cells resulted negative at all the examined time points for the Alizarin Red staining (Fig. 1A).
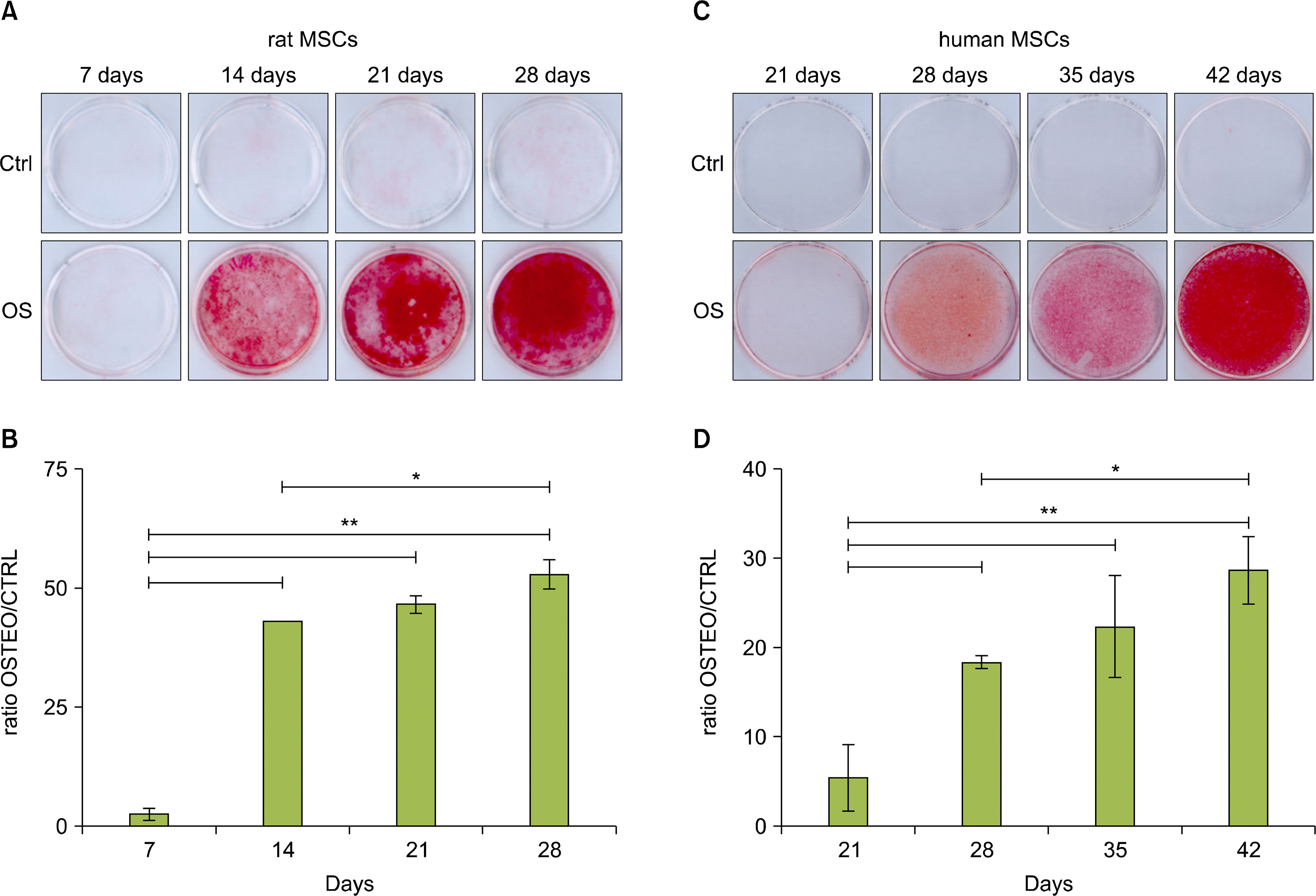 | Fig. 1.Osteogenic differentiation of rMSCs and hMSCs. Control (CTRL) and osteogenic induced (OSTEO) rMSCs (A) or hMSCs (C) were stained with Alizarin Red respectively at days 7, 14, 21 and 28 or at days 21, 28, 35 and 42 after induction and micrographs were taken. Quantitative analysis of Alizarin Red dye accumulation in rMSCs (B) and in hMSCs (D) cultures. The results were normalized to the absorbance of untreated control cells and they are expressed as mean±Standard Deviation. **p<0.001; *p<0.05.
|
In hMSC cultures, the Alizarin Red positive staining was evident only after 28 days of treatment with osteogenic medium and the intensity of the staining increased at the later times examined, as also demonstrated by the quantitative analysis of mineralized matrix deposition (Fig. 1C and D). However, in some hMSC cultures from different donors, this Alizarin Red S positive mineral deposition was observed later, after 35 or 42 days of osteogenic medium treatment. In control hMSC cultures Alizarin Red staining was negative (Fig. 1C).
Adipogenic differentiation
Small translucent Oil Red O positive lipid droplets were evident in the cytoplasm of some adipogenic treated rMSCs starting form 14 days of induction (Fig. 2A). The number of cells with intracellular lipid droplets increased slightly but not in a significant way after 21 days of treatment and it was estimated as being about 5% of the treated cells. The number of cells with lipid droplets did not change at later times but, at 35 days post-induction, the droplets’ size increased in a few cells. The quantitative analysis of Oil Red O accumulation evidenced that, after the initial increased after 14 days of treatment, the dye amount reached a steady level which was maintained for all the observation time (Fig. 2B). In control rMSC cultures, no cells with Oil Red O positive lipid droplets were observed (Fig. 2A).
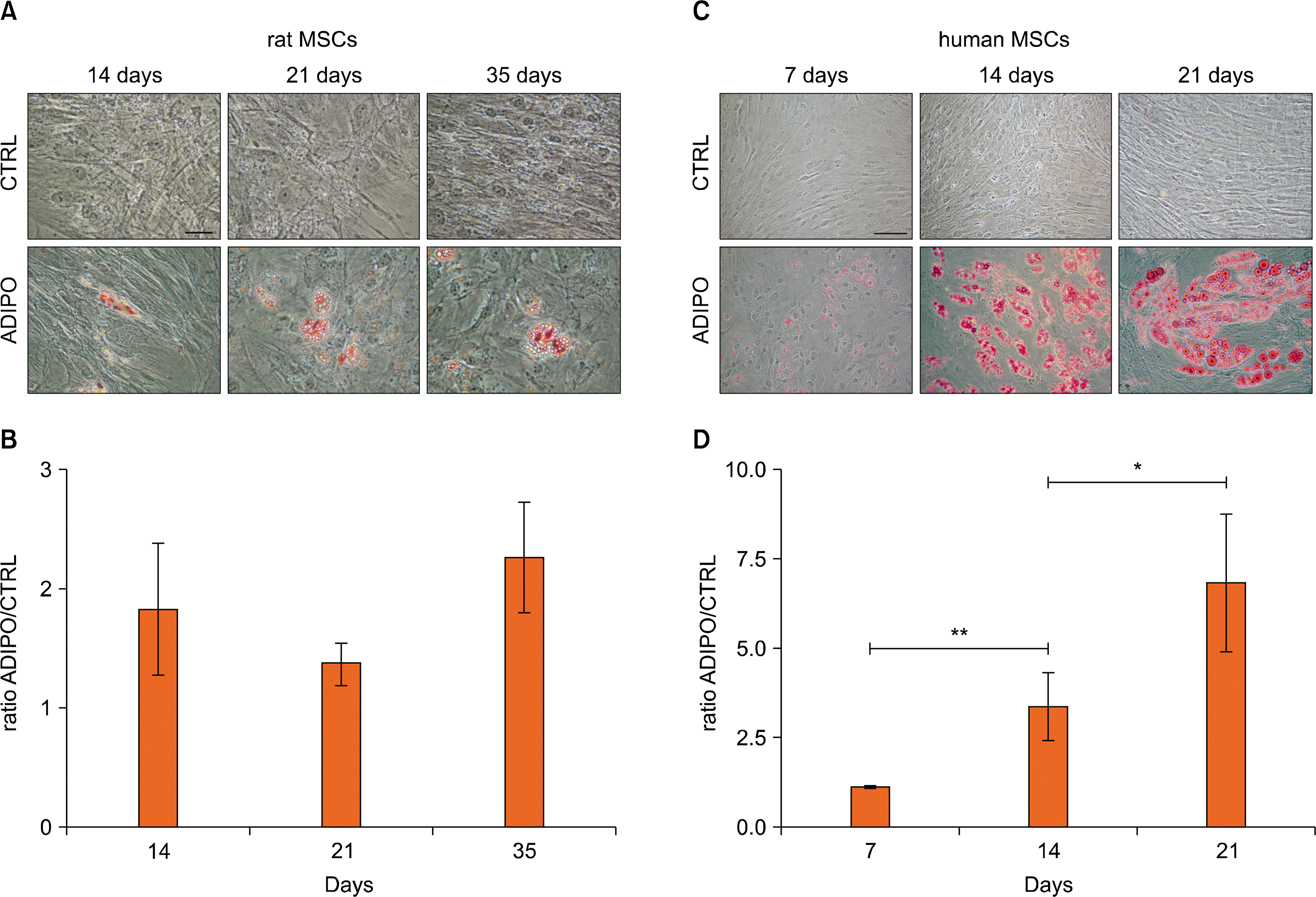 | Fig. 2.Adipogenic differentiation of rMSCs and hMSCs. Control (CTRL) and adipogenic induced (ADIPO) rMSCs (A) or hMSCs (C) were stained with Oil Red O respectively at days 14, 21 and 35 or at days 7, 14 and 21 after induction and micrographs were taken. Bars=50 μm. Quantitative analysis of Oil Red O dye accumulation in rMSCs (B) and in hMSCs (D) cultures. The results were normalized to the absorbance of untreated control cells and they are expressed as mean±Standard Deviation. **p<0.001; *p<0.05.
|
Small translucent lipid droplets positive for Oil Red O staining were evident in the cytoplasm of some adipogenic treated hMSCs (<5%) after 7 days of treatment. The number of these cells increased at later days, reaching the maximum (about 20∼25%) at day 14 (Fig. 2C). The number of cells with lipid droplets did not change at later times but their size increased, probably as a result of a process of droplet fusion. The quantitative analysis of Oil Red O staining demonstrated a progressive and significant increase of dye accumulation during the differentiation period (Fig. 2D). In control hMSC cultures, no cells with Oil Red O positive lipid droplets were observed at any time (Fig. 2C). Variations in terms of the timing of differentiation were observed depending on the donors, since in some cases the differentiation process started to be evident later (14 days rather than 7 days).
Chondrogenic differentiation
Analysis of paraffin-embedded rMSC pellets treated for 5 weeks with chondrogenic medium revealed the presence, in a few areas of the sections, of big oval or polygonal cellular structures with abundant cytoplasm (Fig. 3). After 6 weeks of chondrogenic treatment, similar structures were present throughout the sections. The matrix in which such cells were located stained positively for Safranin O confirming the presence of proteoglycans and glycosaminoglycans. Moreover, many cells were located in small areas that resembled cartilaginous lacunae, sometimes with the presence of more than one cell, similar to cartilage isogen groups. Control pellets did not show the presence of any specific cellular structure and were negative for Safranin O staining (Fig. 3).
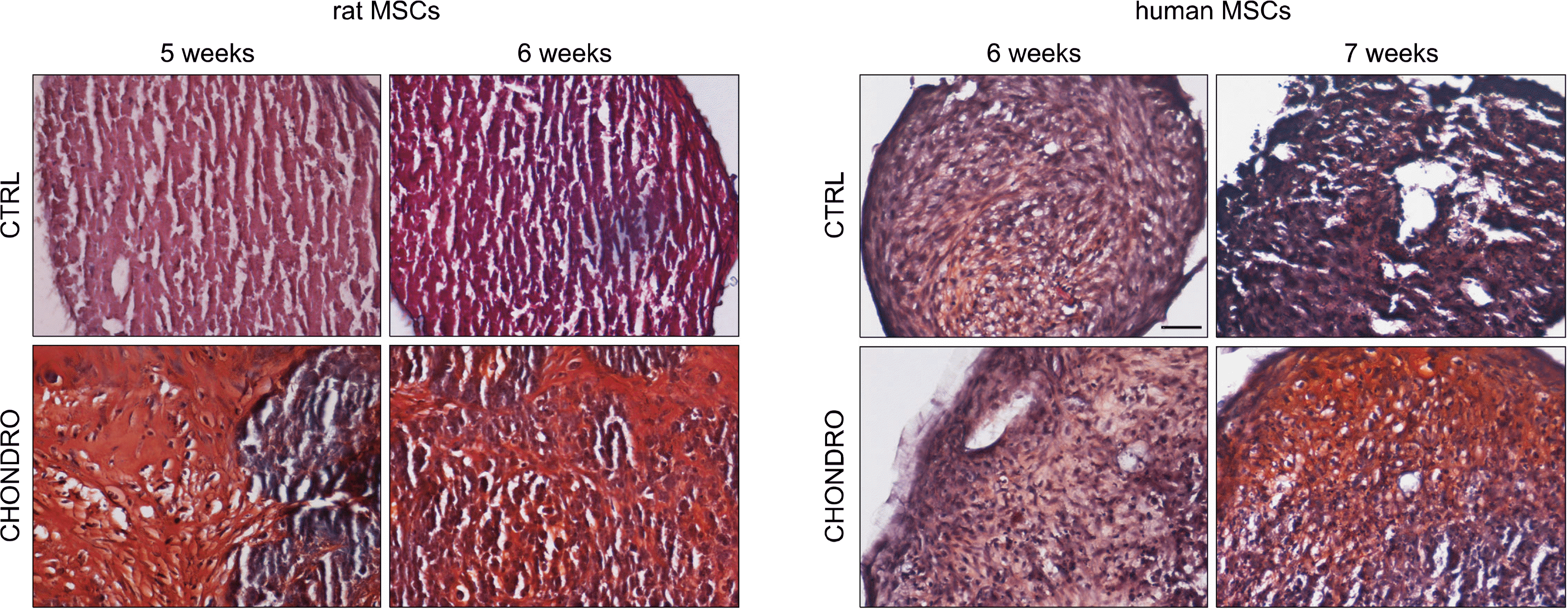
Cartilaginous-like structures positive for Safranin O were not observed in hMSC treated pellets up to 6 weeks of treatment with chondrogenic medium, and only after 7 weeks of chondrogenic induction, they became evident (Fig. 3). Control hMSC pellets were always Safranin O negative and did not show cartilaginous-like structures.
Go to :
Discussion
In view of a therapeutic application of MSCs for tissue repair and regeneration, many studies have been performed to evaluate the differentiation potential by using MSCs from various species, mainly from rodents. Among rodents, the rat model has several advantages with respect to mouse (23) representing a valid and widely used experimental model for diseases affecting tissues of mesengenic origin (24). However, the results obtained from MSCs of different species are not fully comparable, since intrinsic differences exist and often do not allow the transfer of the findings observed (25).
The results of the present study describe some differences in differentiation ability between human- and rat-derivedMSCs. For osteogenic and chondrogenic lineages, the time taken to achieve a differentiated phenotype represents the most evident difference between human and rat MSCs. On the contrary, for adipogenesis the differentiation ability differs greatly, both in terms of time and above all of yield, since only a very few rat MSCs undergo adipogenic differentiation, in spite of a larger number of MSCs of human origin.
A dissimilar differentiation potential among MSCs of different species has already been reported (25): sheep-and rabbit-derived MSCs have been reported as having a greater tendency to differentiate towards chondrogenic lineage with respect to human MSCs which take a longer time to differentiate. The diversity among MSCs is not limited to the differentiative abilities, but it reflects a slightly different molecular pattern, since it has been reported that MSCs from different species do not express the same surface antigens (19), do not have the same alkaline phosphatase activity (26), do not respond to the growth factor BMP-2 in the same way (27), have a different proliferation rate (14) and have different immunoregolatory capacities (19), and they are also characterized by a different genomic stability (15, 16). Also in terms of isolation there are some differences between species since some MSCs are more difficult both to isolate from bone marrow and to expand in culture than human or rat MSCs (11).
The results emerging from our study also evidenced a certain degree of variation in terms of differentiation time among MSCs of human origin. This is probably due to the intrinsic variability of each individual (28), since many factors influence the differentiation process, such as age, sex of the donors (29), or simply a different “propensity” of some bone marrow to produce mesengenic-derived tissues. The biological individual variability of MSCs of human origin raises a potential problem for the clinical use of autologous cells, since the MSCs from one patient may take longer to differentiate with respect to those from another donor, thus making it difficult to set up a standard protocol suitable for clinical application. Recently, many studies have been directed at speeding up the differentiation process. In particular, the use of certain chemicals or some trophic factors, such as FGF4, seems to improve the MSCs’ expansion rate (30), while other factors, such as FGF, PDGF and TGFβ, support the mesengenic differentiation process (31). Moreover, several authors have demonstrated that culturing MSCs under hypoxic conditions can enhance their production, enrichment and their overall differentiation (32) towards osteogenic (33), adipogenic (34) and chondrogenic lineages (35). All these approaches may help to improve MSCs’ usability overcoming the donor-dependent variability of human MSCs.
There are great differences also among MSCs from different rodent strains in terms of growth and differentiation (12). However, the rat MSCs used in our study came from animals of the same age, sex and strain and no variability in differentiation ability was observed.
In conclusion, on the basis of our data, it is important to carefully evaluate the results of pre-clinical studies performed with animal MSCs before attempting to transfer them into clinic. The knowledge of the peculiar differentiative features of MSCs derived from different sources can help to select the populations the more fitting to reach the proposed aim, since the different cellular source may be a misleading factor, and to better apply the results into clinical practice.
Go to :
 XML Download
XML Download