

 PDF
PDF Citation
Citation Print
Print


Introduction
Genomic imprinting is an epigenetic event in which certain genes are mono-allelically expressed according to the parental origin. Most imprinted genes are clustered in a specific chromosomal loci that include protein-coding imprinted genes and non-coding RNA genes (1). The imprinted genes are regulated by highly ordered networks of epigenetic marks such as DNA methylation and histone modifications (2). In particular, imprinted control region (ICR) located in the imprinted cluster is differentially methylated to acquire mono-allelic expression. Differential methylation status of imprinted genes is resistant to the epigenetic reprogramming during preimplantation development in mammals (3). Instead, new methylation marks of the imprinted genes are acquired in the differentially methylated regions (DMRs) during gametogenesis. The newly acquired imprinted status is generally maintained throughout the life. Nonetheless, some imprinted genes show variable allelic expression patterns in a developmental stage- or tissue-specific manner (4). Studies about tissue-specific imprinting in human have mainly been performed in placenta, suggesting that tissue-specific expression of imprinted genes is crucial for fetus developments (5). Many imprinting disorders are closely related to abnormal loss- or gain-of imprinting in placental tissues (5). So far, approximately 50 to 100 imprinted genes have been identified in human and mouse, respectively (5). Recently, high-throughput sequencing in the mouse genome enables to identify differential methylation status of the newly identified imprinted loci in a base-level resolution (6). In spite of a small fraction in the whole genome, fine-tuned regulation of imprinted genes is crucial for mammalian development, growth, metabolism, and behaviors (7). Aberrant regulation of imprinted genes leads to various developmental disorders such as Prader-Willi /Angelman syndrome, Bechwith-Wiedemann syndrome, and Silver-Russell syndrome (7). Those imprinting disorders are attributed to genetic and epigenetic mutations in the imprinted genes or imprinting control region (ICR) (7). Thus, fine-tuned regulation of the imprinted gene expression is essential for mammalian development. Studies about genomic imprinting in mammals have been mainly performed in knock-out mice. The in vivo experiments are time-consuming and labor-intensive, and gene-targeting technique is not available in human in vivo. In addition, allelic expression status of some imprinted genes in mice is not concordant with human (8). Therefore, human embryonic stem cells (hESCs) can be alternative sources to study genomic imprinting underlying human development because they are able to differentiate to diverse cell types. In this study, we examined total transcriptional levels of 19 imprinted genes in hESCs during differentiation into specialized cell types. To compare expression levels of the imprinted genes between different developmental stages and lineages, hESCs were differentiated into ectodermal (neural progenitors and dopaminergic neurons), mesodermal (mesenchymal progenitors and osteoblasts), and endodermal lineage cell types (definitive endoderm cells and hepatocytes). Our results showed that respective imprinted genes were differently expressed in a cell type-specific manner during in vitro differentiation of hESCs.
Go to : 
Materials and Methods
Maintenance of human pluripotent stem cells (hPSCs)
Two hESC lines {H9-hESCs (passage No.: 36∼70) and CHA4-hES (passage No.: 52∼71)}, and two hiPSC lines {CRL-hiPSCs (passage No.: 32∼40) and HDF-hiPSCs (passage No.: 35∼40)} were maintained in the ESC mediun on mouse embryonic fibroblasts (MEFs) treated with Mitomycin-C (Sigma-Aldrich, MO, USA). The ESC medeiun is DMEM/F12 supplemented with 20% KO-SR (Invitrogen, CA, USA), 100 IU/ml penicillin (Invitrogen), 1 mM L-glutamine, 100 μg/ml streptomycin (Invitrogen), 1% non-essential amino acids (NEAA; Invitrogen), 0.1 mM β-mercaptoethanol (Sigma-Aldrich), and 4∼8 ng/ml basic fibroblast growth factor (bFGF; R&D Systems, MN, USA). The medium was daily changed, and passaging of hPSCs was performed at an interval of 5∼6 days.
Differentiation of hESCs into dopaminergic neurons, osteoblasts, and hepatocytes
For differentiation of hESCs into dopaminergic (DA) neurons (9), undifferentiated H9-hESCs were seeded on MS5 stromal feeder cells and cultured on insulin/transferrin/selenium (ITS, Invitrogen) medium with 0.2 mM ascorbic acid (Sigma-Aldrich). After 7 days of culture, the cells were passaged onto MS5-SHH cells and cultured for 14 days. Then, cell clusters were further differentiated into neural precursor cells (NP) by culturing in ITS medium supplemented with 20 ng/ml bFGF and 0.2 mM AA, and re-plated on poly-L-ornithine (15 μg/ml; Sigma-Aldrich)/ fibronectin (1 μg/ml; Sigma-Aldrich)-coated dishes. hESC-NP cells were passaged every 7 days in medium supplemented with bFGF. DA neuron differentiation from hESC-NP cells was induced by withdrawing bFGF from the medium and adding brain-derived neurotrophic factor (BDNF, 20 ng/ml; R&D Systems), glial cell line-derived neurotrophic factor (20 ng/ml; R&D Systems), and dibutyryl cAMP (0.5 mM; Sigma-Aldrich).
Osteoblasts were differentiated from hESC-derived mesenchymal progenitors (MPs) as previously described (10). Briefly, hESCs were dissected into small clumps, transferred to a petri dish, and cultured in embryoid body (EB) medium (EBM; DMEM/F12, 10% SR without bFGF and 0.1 mM β-mercaptoethanol) supplemented with 10 μM SB431542 (SB) (Sigma-Aldrich). After 10 days of culture, hEBs were transferred to fibronectin-coated dishes in serum-free medium (DMEM/F12 supplemented with 1% ITS, 2% B27, and 1% concentrated lipids (Invitrogen)) containing 1 μM SB431542 (SB1-treated cells). The attached EBs were cultured to confluence, followed by expansion over three or four passages. To induce MPs, SB1-treated cells were seeded on FN-coated dishes at a density of 104 cells/cm2 in α-MEM medium containing 10% FBS. Medium was refreshed every other day. After 20 days in culture, induced cells showing homogeneous morphology were characterized. For osteogenesis, MPs were seeded at a density of 0.5×104 cells/cm2 into culture vessels containing α-MEM medium supplemented with 10% FBS. Cells were incubated at 37°C, 5% CO2 for 1 day. When the cells reached to 50∼60% confluence, medium was replaced by pre-warmed osteogenesis complete medium containing STEMPRO Osteogenesis Supplement (Invitrogen). Medium was refreshed every 3 days. After 2∼ 3 weeks, cells were analyzed by Alizarin Red staining and Von Kossa staining.
Derivation of hepatocytes from hESCs was carried out as previously described (11), Briefly, hESCs were cultured to confluence in conditioned medium on Matrigel for 2 days. To induce definitive endoderm (DE), hESCs were cultured in RPMI-1640 (HyClone, UT) containing 1% B27 (GIBCO) and 50 ng/ml activin A (Peprotech) for 5 days. To further differentiate into hepatocytes, DE cells were cultured in hepatocyte culture medium (HCM, Lonza) containing 30 ng/ml FGF4 (Peprotech) and 20 ng/ml BMP2 (Peprotech) for 5 days and further incubated in HCM supplemented with 20 ng/ml hepatocyte growth factor (Peprotech) for 5 days. Finally, hESC-derived hepatocytes were matured by culturing in HCM supplemented with 10 ng/ml oncostatin M (R&D Systems) and 0.1 μM dexamethasone (Sigma-Aldrich) for 5 days.
Total RNA extraction and qRT-PCR
Total RNAs were extracted from undifferentiated hPSCs and hESC-derived cells using TRIzol® (Invitrogen) according to the manufacturer’s procedure. Then, extracted RNAs were treated with DNase I to eliminate genomic DNA. cDNAs were synthesized by using M-MLV reverse transcriptase (Enzynomics, Korea), and diluted 1:5 ratio with RNase- and DNase-free water. The cDNAs were analyzed by real-time quantitative PCR (qPCR) on a Bio-RAD iQ5 cycler real-time PCR Detection System (Bio-Rad, CA, USA) using the Primer Q-Master Mix (GenetBio, Chungnam, Korea). PCR conditions were 10 min at 94°C (one cycle), 30 sec at 94°C, 30 sec at 60°C and 30 sec at 72°C (50 cycles). Melting-curve analysis was performed to ensure specific amplification. All sets of reactions were conducted in triplicate. Expression levels of imprinted genes were normalized to that of GAPDH as an internal control, and calculated using the comparative CT method (ΔΔCT). The primers used in this study were designed using Primer3Plus and are listed in Supplemental Table 1.
Table 1.
Information for Quantitative real-time PCR primer sets
![]()
Statistics
Statistical analyses were performed using Prism 5.01 (GraphPad Software, USA). Also, one-way ANOVA and Dunnett’s post-hoc test were used to confirm the statistical significance. The differences of gene expression between control and experimental groups were considered significant if the p value was less than 0.05.
Go to :
Results
Conservative expression of imprinted genes in undifferentiated hPSC lines
To check whether imprinted genes are expressed in un-differentiated hPSCs, 24 imprinted genes which are known to be mono-allelically expressed in human tissues (http://igc.otago.ac.nz/home.html) were tested in two hESC lines and two hiPSC lines. Of 24 imprinted genes, 19 imprinted genes were expressed in H9-hESCs (data was not shown). Transcriptional expression of 19 imprinted genes was also observed in other hPSCs (Fig. 1). The relative expression levels of imprinted genes did not show distinct difference between hPSC lines (2 hESC lines and 2 hiPSC lines) except some genes such as DIO3, RTL1, OSBPL5, and NDN. These results indicate that the relative expression levels of imprinted genes are generally conserved in undifferentiated hPSC lines.
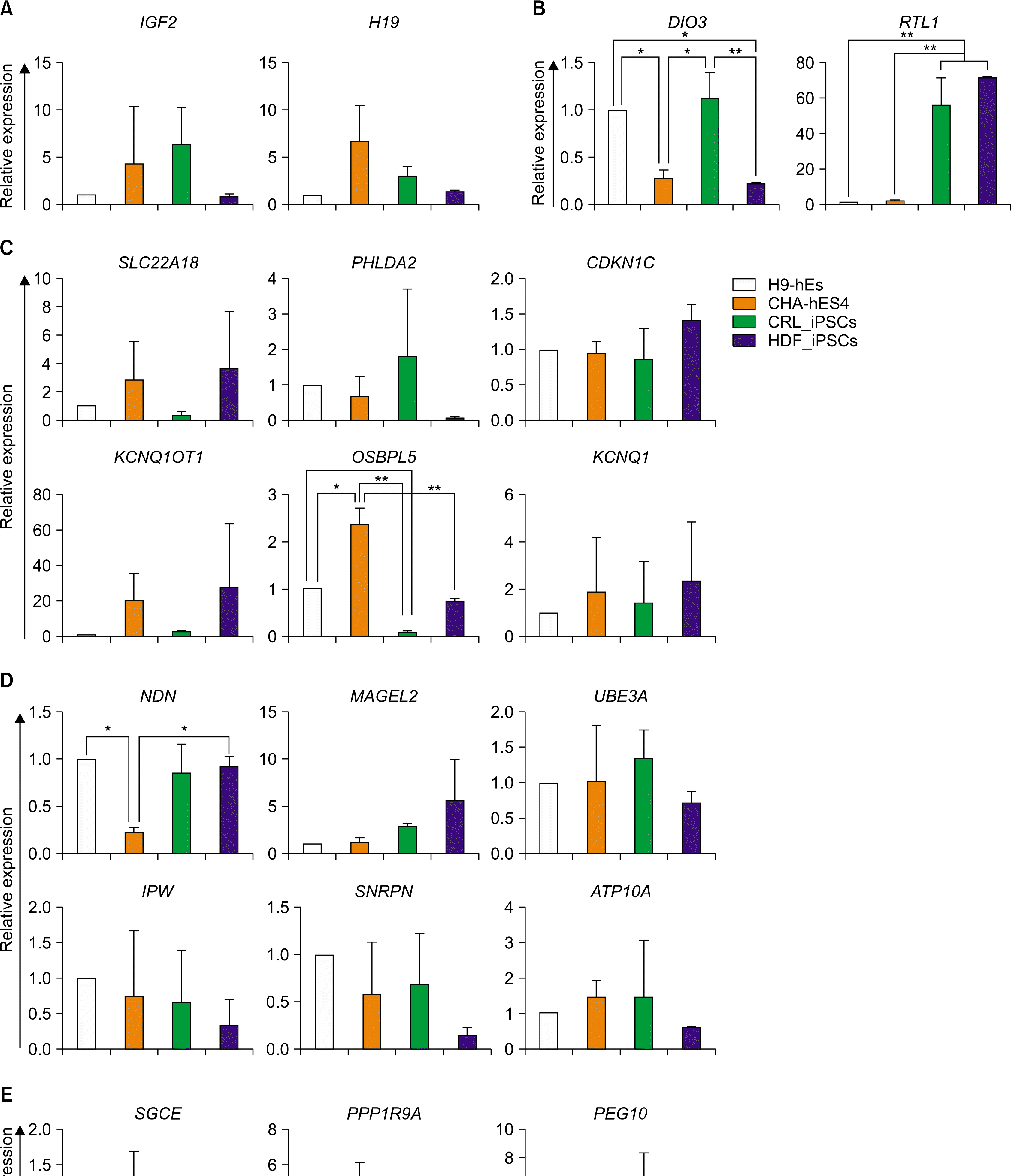 | Fig. 1.Transcriptional expression of imprinted genes between four hPSC lines. Transcriptional levels of the imprinted genes that are included in IGF2-H19 (A), DIO3-DLK1 (B), KCNQ1 (C), PWS/AS (D), and PEG10 (E) domains were examined in undifferentiated hPSCs by qRT-PCR. The error bars indicate±SD for three independent experiments (n=3, *p<0.05, **p <0.01). Abbreviations: CRL_iPSCs; hiPSCs derived from CRL foreskin fibroblasts, HDF_iPSCs; hiPSCs derived from human dermal fibro-blasts.
|
Quantitative expression of the imprinted genes in hESCs during in vitro differentiation
To investigate the expression level of imprinted genes during lineage-specific differentiation, H9-hESCs were differentiated into dopaminergic neurons (ectoderm), osteoblasts (mesoderm) and hepatocytes (endoderm) as representative cell types of 3 germ layers, respectively. Characterization and functionality of differentiated cell types from hESCs were previously reported (12). In this study, we analyzed the transcriptional expression of 19 imprinted genes that were commonly expressed in 4 different hPSC lines. These 19 imprinted genes were categorized in well-known imprinted domains, including IGF2/H19 domain (Fig. 1A), DIO3-DLK domain (Fig. 1B), KCNQ1 domain (Fig. 1C), PWS/AS domain (Fig. 1D), and PEG10 domain (Fig. 1E). Total transcriptional levels of the imprinted genes were investigated in three developmental stage cell types such as pluripotent stem cells, intermediate progenitors, and terminally-differentiated cells, respectively.
IGF2/H19 domain: IGF2/H19 (chr.11p15) imprinting domain contains paternally-expressing IGF2 gene and maternally-expressing non-coding RNA gene H19. The expression of these imprinted genes is regulated by an insulator binding protein CTCF (CCCTC-binding factor) (13). CTCF binds to unmethylated DMRs on maternal allele, and blocks the expression of IGF2. In contrast, expression of H19 is modulated by downstream enhancer. Since CFCF does not bind to methylated DMRs, the enhancers activates IGF2 gene in the paternal allele, not H19 gene (Fig. 2A). Transcription of both IGF2 and H19 was up-regulated in hESCs during differentiation to neurons (Fig. 2B) and hepatocytes (Fig. 2D). However, transcriptional expression of H19 reduced in hESCs during osteoblastic differentiation (Fig. 2C).
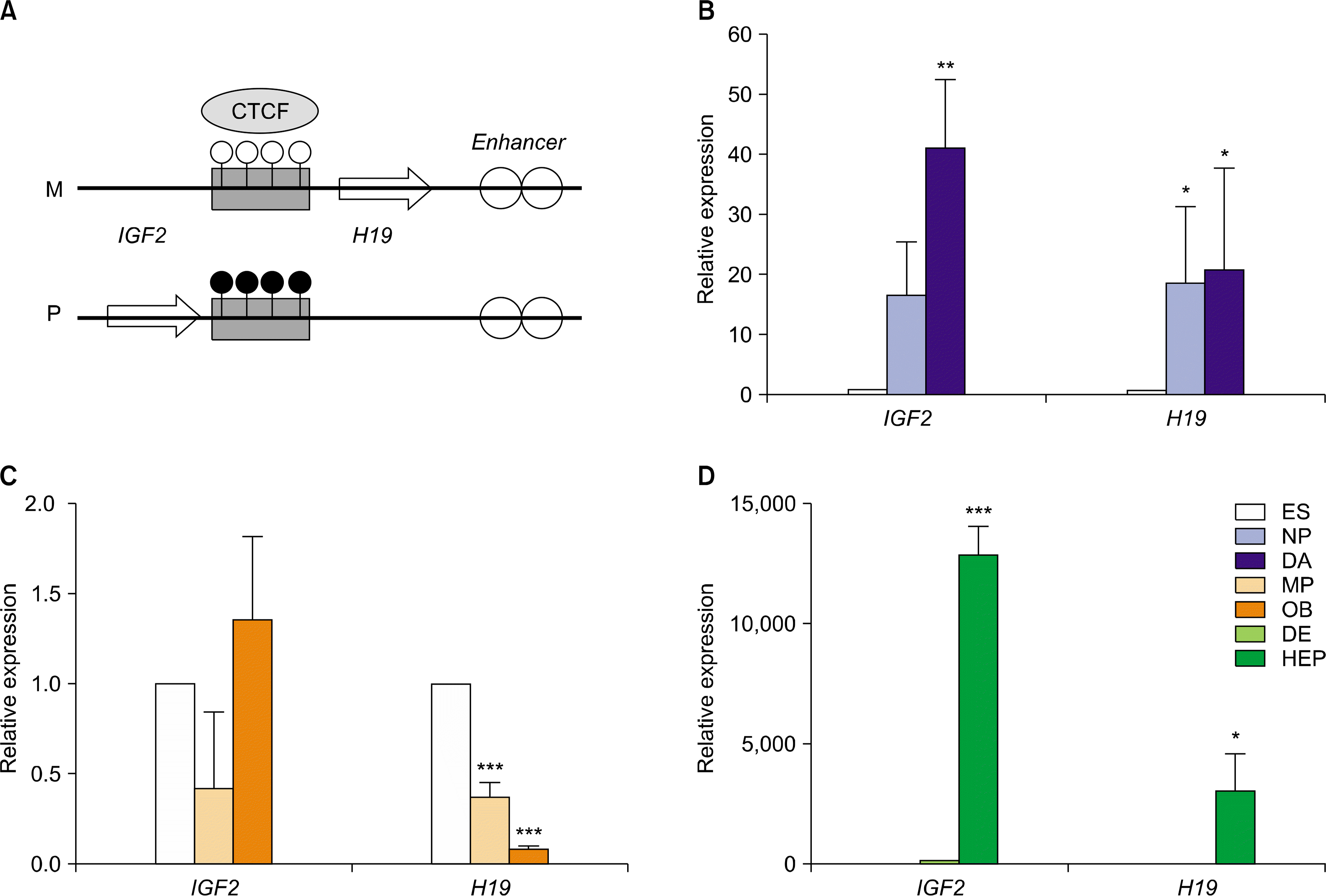 | Fig. 2.Relative expression levels of imprinted genes in IGF2/H19 domain in hESCs during in vitro differentiation. (A) A schematic diagram of IGF2/H19 domain (Ch 11p15). Relative expression level of IGF and H19 imprinted genes in hESCs during in vitro differentiation to DA neurons (B), osteoblasts (C), and hepatocytes (D). The error bars indicate±SD for three independent experiments (n=3, *p<0.05, **p<0.01, ***p<0.001). Abbreviations, ES: H9-hESCs; NP: neuronal progenitors; DA: dopaminergic neurons; MP: mesenchymal progenitors; OB: osteoblasts; DE: definitive endoderm; HEP: hepatocytes.
|
KCNQ1 domain: KCNQ1 domain (ch.11p15) has 6 maternally-expressing genes and a paternally-expressing KCNQ1OT1 (Fig. 3A). The genomic imprinting at this domain is controlled by maternally-expressing non-coding RNA gene KCNQ1OT1 (Fig. 3A). Quantitative expression level of imprinted genes in this domain showed distinct patterns in a cell-type dependent manner during in vitro differentiation (Fig. 3B∼D). During neuronal differentiation of hESCs, expression levels of OSBPL5, PHLDA2, and KCNQ1OT1 did not changed in neural progenitors and DA neuron (Fig. 3B). However, relative expression level of CDKN1C and KCNQ1 was enhanced in neural progenitor (NPs) and DA neurons, respectively. Besides, all imprinted genes in KCNQ1 domain significantly decreased in hESCs during differentiation to osteoblasts (Fig. 3C). In contrast, most of the imprinted genes showed enhanced expression in hESC-derived hepatocytes (Fig. 3D). These results indicate that transcriptional expression of the imprinted genes in KCNQ1 domain may be regulated in a cell type-dependent manner.
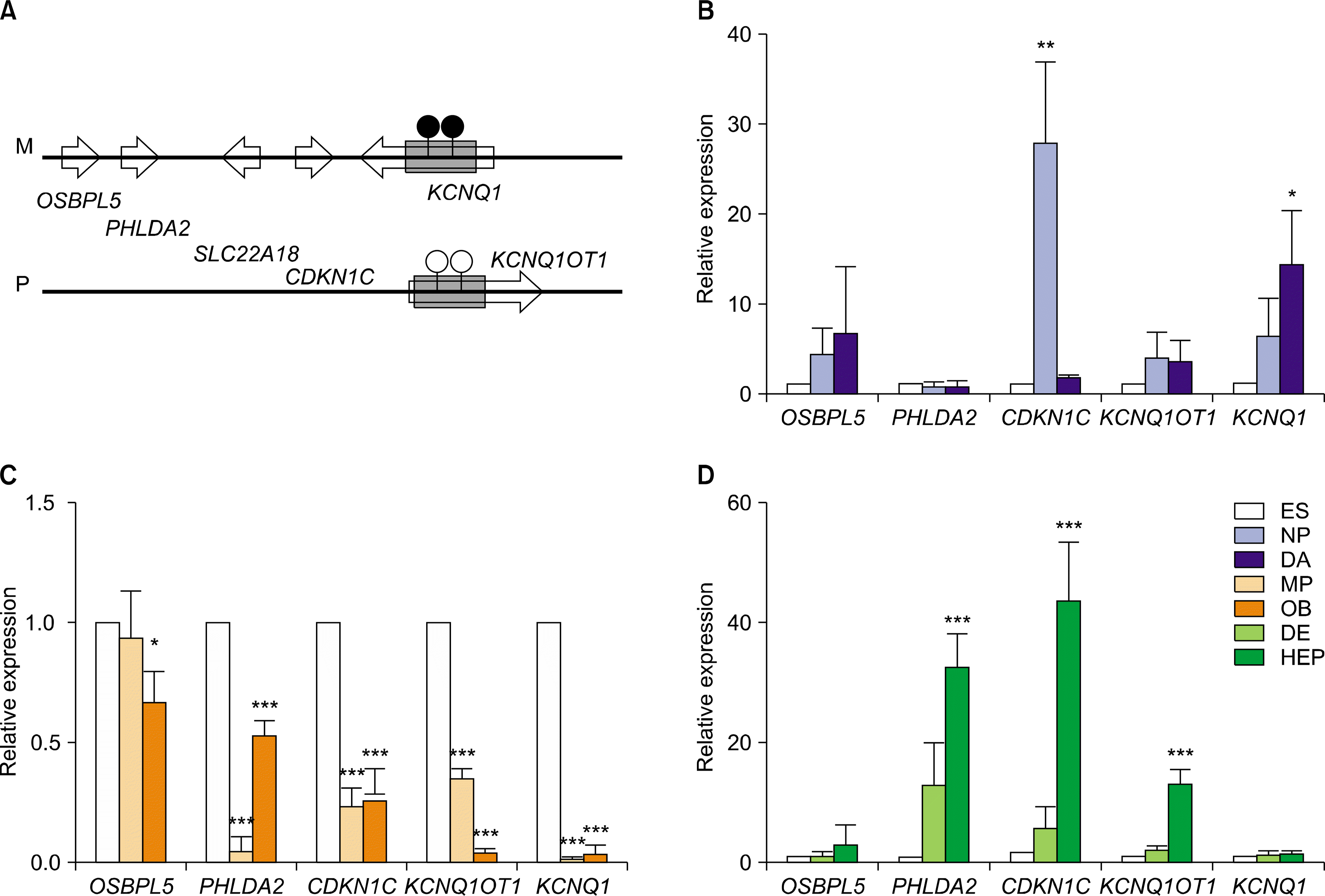 | Fig. 3.Relative expression levels of imprinted genes in KCNQ1 domain in hESCs during in vitro differentiation. (A) A schematic diagram of KCNQ1 domain (Ch 11p15). This KCNQ1 domain contains several imprinted genes, including OSBPL5, PHLDA2, CDKN1C, KCNQ1, and KCNQ1OT1. Relative expression level of several imprinted genes in hESCs during in vitro differentiation to DA neurons (B), osteoblasts (C), and hepatocytes (D). The error bars indicate±SD for three independent experiments (n=3, *p<0.05, **p<0.01, ***p<0.001).
|
PWS/AS domain: PWS/AS domain (chr. 15q11–q13) contains four paternally-expressing genes and three maternally-expressing genes (Fig. 4A). In this domain, several imprinted genes represented distinct expression patterns in hESCs during in vitro differentiation. During differentiation of hESCs into DA neurons and hepatocytes, two genes (MAGEL2, NDN) were up-regulated and three genes (SNRPN, IPW, and UBE3A) revealed constant expression levels (Fig. 4B, D). However, MAGEL2, SNRPN, and IPW were significantly down-regulated in hESCs during osteoblastic differentiation (Fig. 4C).
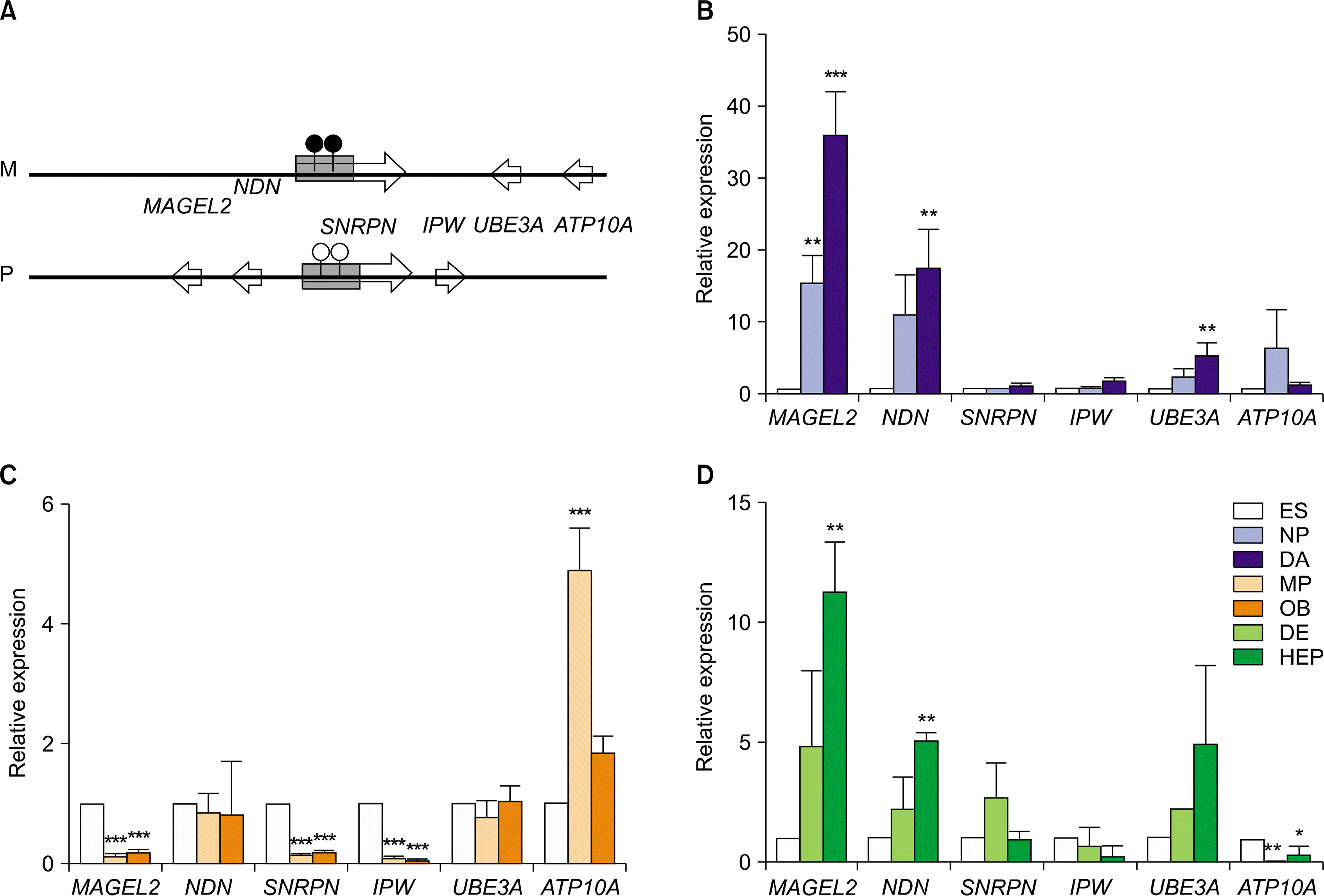 | Fig. 4.Relative expression levels of imprinted genes in PWS/AS domain in hESCs during in vitro differentiation. (A) A schematic diagram of PWS/AS domain (Ch. 15q11-q13). This domain contains several imprinted genes, including MAGEL2, NDN, SNRPN, IPW, UBE3A, and ATP10A. Relative expression level of several imprinted genes in hESCs during in vitro differentiation to DA neurons (B), osteoblasts (C), and hepatocytes (D). The error bars indicate±SD for three independent experiments (n=3, *p<0.05, **p<0.01, ***p<0.001).
|
PEG10 domain: PEG10 domain (chr.7q21.3) has two paternally-expressing imrinted genes and one maternally-expressing gene (Fig. 5A). The paternally-expressing gene 10 (PEG10) is a retrotransposon-derived gene that is important to placental development (14). This imprinting locus is controlled by DMR located in PEG10 promoter region (Fig. 5A). The imprinted genes in PEG10 domain also exhibited distinct expression patterns between different cell types derived from hESCs. Even though PPP1R9A showed invariable expression levels in all three cell types, SGCE and PEG10 were differently regulated in all differentiated cell types. Although transcription of SGCE was relatively constant in the neuronal lineage, transcripts of SGCE were significantly enhanced in osteoblasts and hepatocytes, respectively (Fig. 5B∼D). Expression of PEG10 was significantly upregulated in hESCs during differentiation to DA neurons and hepatocytes, but down-regulated during osteoblastic differentiation (Fig. 5B∼D).
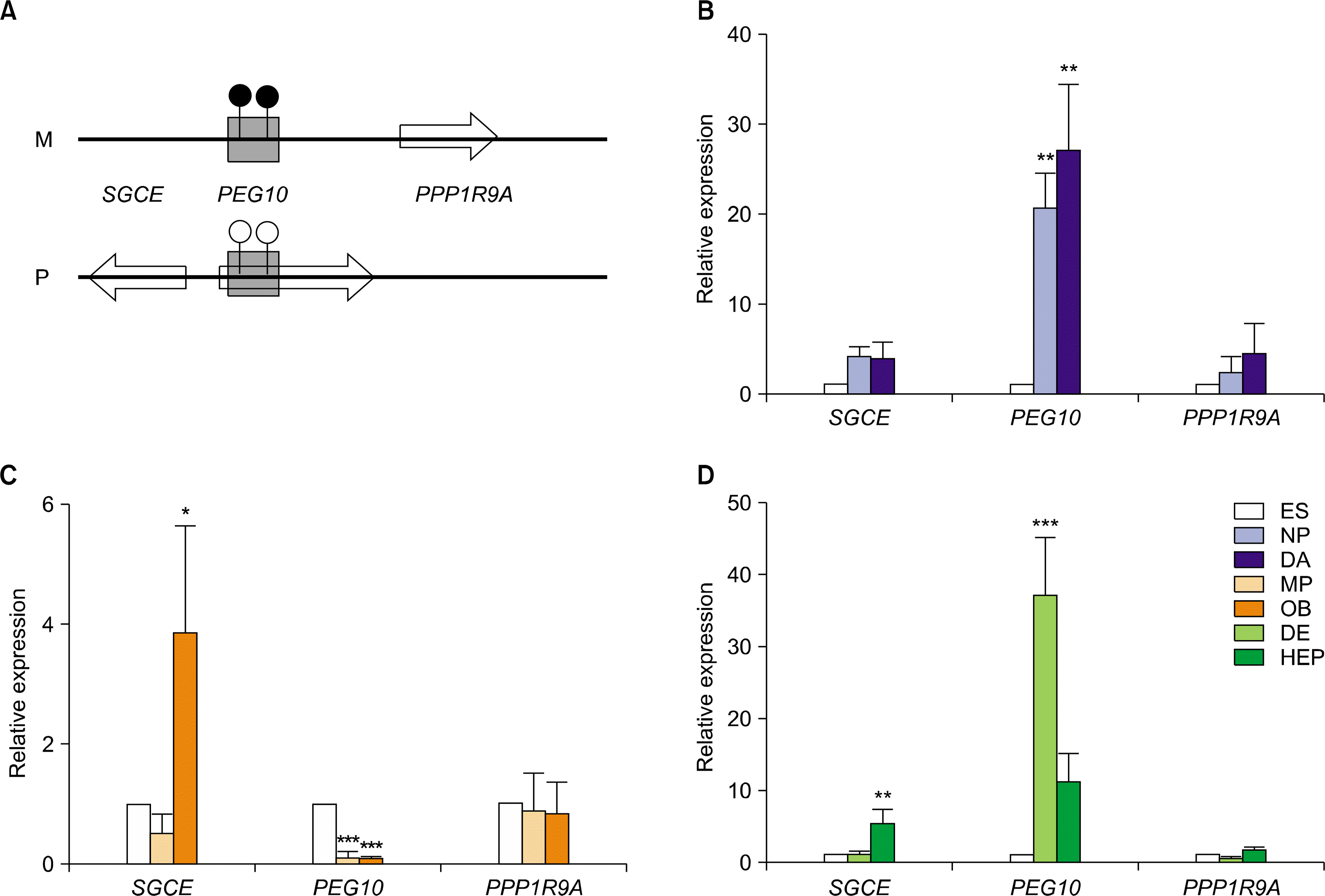 | Fig. 5.Relative expression levels of imprinted genes in PEG10 domain in hESCs during in vitro differentiation. (A) A schematic diagram of PEG10 domain (Ch. 7q21.3). This domain contains three imprinted genes such as SGCE, PEG10 and PPP1R9A. Relative expression level of these imprinted genes in hESCs during in vitro differentiation to DA neurons (B), osteoblasts (C), and hepatocytes (D). The error bars indicate±SD for three independent experiments (n=3, *p<0.05, **p<0.01, ***p<0.001).
|
DIO3-DLK1 domain: In addition to above imprinted genes, we also examined the quantitative expression patterns of DIO3, RTL1, MEG3 and DLK1 that are clustered in chromosome 14q32.2 region (Fig. 6A). Of these imprinted genes, only DIO3 and RTL1 were expressed in un-differentiated hESCs. Transcription of DIO3 was activated in hESCs during differentiation to neuroectodermal and endodermal lineages, not mesodermal lineage, respectively (Fig. 6B∼D). Interestingly, expression of RTL1 was enhanced in hESC-derived differentiated cells such as DA neurons, osteoblasts, and hepatocytes (Fig. 6B∼D).
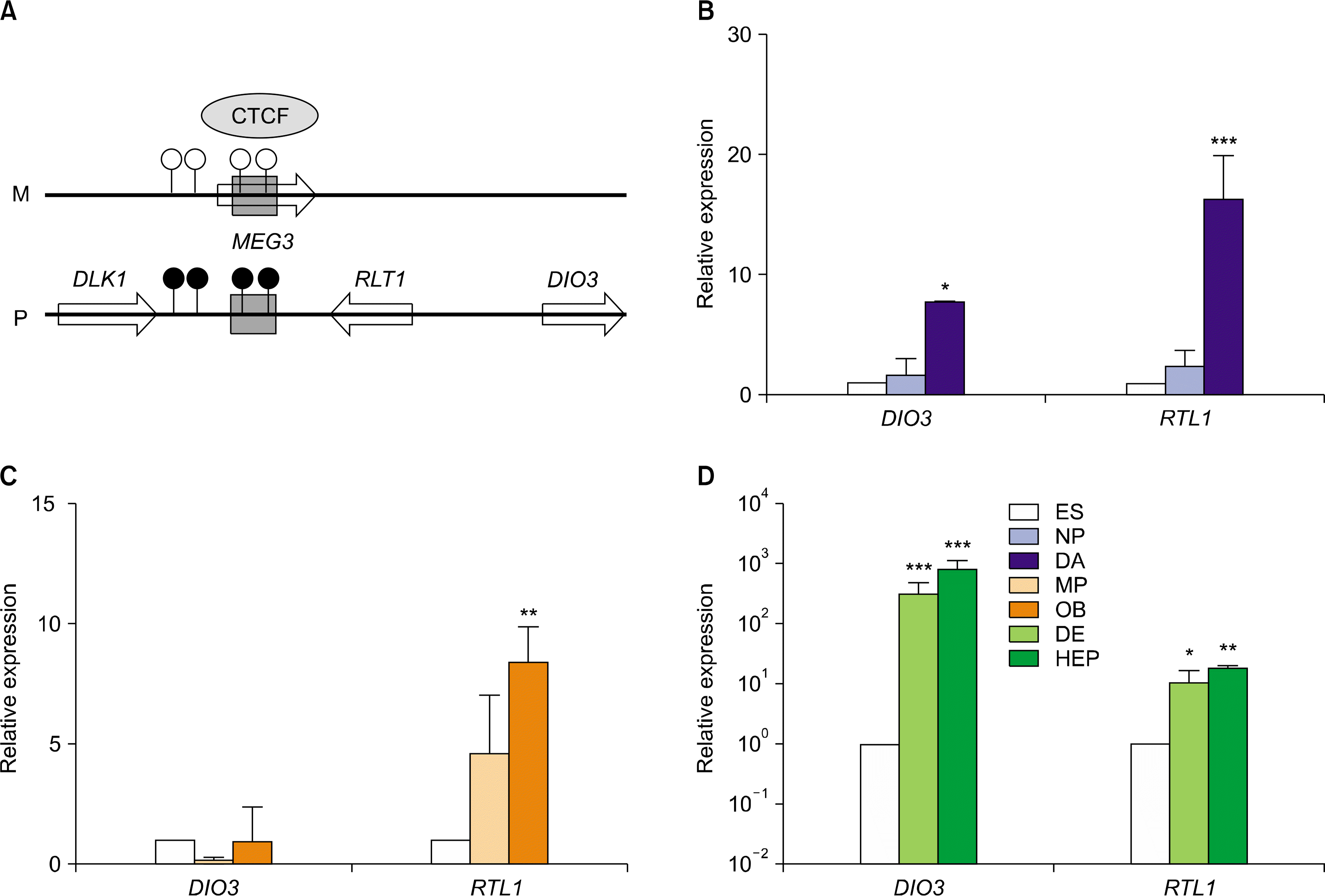 | Fig. 6.Relative expression levels of imprinted genes in DIO3-DLK1 domain in hESCs during in vitro differentiation. (A) A schematic diagram of DIO3-DLK1 domain (Ch. 14q32.2). This domain contains four imprinted genes such as DLK1, MEG3, DIO3 and RTL1. Relative expression level of DIO3 and RTL1 imprinted genes in hESCs during in vitro differentiation to DA neurons (B), osteoblasts (C), and hepatocytes (D). The error bars indicate±SD for three independent experiments (n=3, *p<0.05, **p<0.01, ***p<0.001).
|
Go to :
Discussion
We have previously reported that allele-specific expression of imprinted genes is changed in a cell-type specific manner in human pluripotent stem cells (hPSCs) during in vitro differentiation (12). However, total expression levels of imprinted genes were not still reported in hPSCs during in vitro differentiation. Here we demonstrate that total transcript levels of imprinted genes are varied in hPSCs during in vitro differentiation. In general, transcription of imprinted genes is down-regulated in somatic cells (15). In contrast, expression of some imprinted genes (PEG10, NDNL1, IGF2, GNAS, DCN, and H19) is up-regulated in hESCs during the formation of embryoid body (EB) (16). However, there is a limitation to understand cell type- or tissue-specific imprinting because EBs consist of mixed cell populations, which include different cell types (ectoderm, endoderm, and mesoderm) as well as undifferentiated cells. Imprinting of some imprinted genes is changed in a developmental stage- or tissue-specific manner (17). Therefore, hESCs undergoing differentiation into specialized cell types, rather than EBs derived from hESCs, would be better in vitro models for studying tissue-specific imprinting. In this study, the transcriptional expression of imprinted genes was varied in a cell type-dependent or a domain- independent manner in hESCs during in vitro differentiation. Transcriptional expression of imprinted genes within IGF2/H19 and KCNQ1 domain was enhanced in hESC- derived neuronal progenitors (18). Similarly, transcript levels of some imprinted genes (IGF2 and H19 in IGF2/H19 domain, CDKN1C and KCNQ1 in KCNQ1 domain) were significantly increased during differentiation of hESCs into the neuronal lineage (Fig. 2B, 3B). Consequently, this study provides a possibility that hESCs can be employed as a valuable source for in vitro modeling to understand the behavior of imprinted genes in the embryonic development and/or cellular differentiation in human.
Go to :
 XML Download
XML Download