

 PDF
PDF Citation
Citation Print
Print


Introduction
Human embryonic stem cells (hESCs) have been regarded as a potential source of therapeutic cells for many diseases caused by tissue loss or dysfunction (1). Because of the self-renewal and pluripotency of hESCs, however, teratoma formation is a seriously raised concern whenever utilizing hESC-derived differentiated cells (2, 3). The number of hESC-derived cell transplantation studies into animal models have been done, but teratoma formation was not detected depending on experimental conditions (4–6). Longer in vitro differentiation of hESCs seems to reduce the risk of teratoma formation due to low survival rate of hESC-derived cells in animal models. However, survived dopaminergic implants in parkinsonian rats exhibit expanding cores of undifferentiated mitotic neuroepithelial cells, which can be tumorigenic (7). Furthermore, hESC-derived neural precursor cells have repeatedly induced tumors in animal models even though remaining hESCs were not detected (8, 9). Thus, the elimination of remaining undifferentiated hESCs is a major hurdle to hESC-derived cell transplantation.
Previously, we generated a panel of MAbs against surface molecules on undifferentiated hESCs (10). One of the MAbs, 57-C11 (IgG1, k), recognizes the phosphorylated form of adenovirus early-1B-55kDa associated protein 5 (E1B-AP5) or heterogeneous nuclear ribonucleoprotein U-like 1 (hnRNPUL1) (11). E1B-AP5 is a member of the heterogeneous nuclear ribonucleoprotein (hnRNP) family and is ubiquitously expressed in the nucleus and cytoplasm in all tissues (12). However, we showed for the first time that 57-C11-reactive E1B-AP5 is expressed on the cell surface of undifferentiated hESCs and cell surface-expressed E1B-AP5 is downregulated upon the differentiation of hESCs, suggesting that cell surface-expressed E1B-AP5 is a specific surface marker for undifferentiated hESCs (11). In this study, we have tested whether 57-C11 is able to eliminate undifferentiated hESCs from the mixture of undifferentiated and differentiated hESCs. To isolate 57-C11-positive hESCs, 57-C11 was biotinylated and added to the mixture of undifferentiated and differentiated hESCs. Sequentially, streptavidin-coated magnetic beads were added into the same tube. After applying magnet to the mixture, 57-C11-positive and negative hESCs were separated and analyzed. The isolated cells were further validated in in vivo animal model by injecting the isolated cells into right and left testes of the same NOD/SCID mice. The results showed that teratoma formation was not detected with 57-C11-negative hESCs whereas teratoma was detected with 57-C11-positve hESCs. The results indicate that 57-C11-positive hESCs are really pluripotent and tumorigenic while 57-C11-negative hESCs are not. The results also suggest that 57-C11-mediated isolation system will be useful for the elimination of residual undifferentiated hESCs from the preparation of hESC-derived differentiated cells.
Materials and Methods
Cell culture
H9 hESCs were cultured on irradiated mouse embryonic fibroblast (MEF) as described previously (13, 14). hESCs were fed daily with DMEM/F12 medium supplemented with 20% Knockout serum replacement (Life Technologies, Seoul, Korea), 0.1 mM 2-Mercaptoethanol, 1% Non-essential amino acid, 1 mM glutamine, 100 U/ml penicillin G, 100 μg/ml streptomycin, and 4 ng/ml basic fibroblast growth factor (PeproTech, Rocky Hill, NJ, USA). Feeder-free culture method was performed as described previously (13, 15). Briefly, 10 μM of Y27632 (Sigma-Aldrich, Seoul, Korea) were treated for 1 hour prior to detaching hESCs, the cells were harvested with trypsin-EDTA. After elimination of MEF cells on gelatin coated tissue culture dish, hESCs were cultured in conditioned medium supplemented with 10 μM of Y27632 on tissue culture dish coated with Matrigel (BD Biosciences, Seoul, Korea). For differentiation of hESCs, cells were treated 10−5 M retinoic acids (RA) in culture medium for 10~14 days. Colo205 cells were maintained following recommendation of ATCC.
Antibody biotinylation
Antibody was biotinylated by using DSB-XTM biotin labeling kit (Life Technologies) according to manufacturer’s procedure.
Isolation of 57-C11-positive and -negative hESCs
Cell isolation was carried out by using Dynabeads FlowComp Flexi kit (Life Technologies) according to the manufacturer’s protocol. The cells were detached with dissociation solution for human ES/iPS cells (ReproCELL). To prevent physiological cell damage, dissociation buffer was immediately neutralized by complete medium. The mixture of undifferentiated and RA-treated hESCs were suspended in 500 μl of cold isolation buffer (phosphate buffered saline (PBS), pH 7.4, 5% Knockout serum replacement, 1 μg/ml Doxycyclin, 0.5 mM Mg2+, 1 mM Ca2+) in Eppendorf tube and incubated with 20 μg of biotinylated 57-C11 for 30 min at 4°C with rolling and tilting. Cells were harvested after addition of 500 μl of cold isolation buffer. Cells (2×106) were then resuspended in 500 μl of cold isolation buffer and incubated for 30 min at 4°C with rolling and tilting after addition of 50 μl (7.5×107 beads) of Dynabeads (Life Technologies). 57-C11-positive hESCs were collected by placing the tube in a magnet for 2~3 min. The supernatant containing 57-C11-negative hESCs was carefully removed in the magnet, and the bead/cell complexes were cleaned up by a total of three washes. Cells were then removed from the magnet by incubation of 1 ml of FlowComp Release Buffer (Life Technologies) for 10 min at room temperature (RT) with rolling and tilting. The supernatant with 57-C11-positive hESCs was carefully transferred to a new tube and put on the magnet again to remove residual beads. The supernatant with 57-C11-positive hESCs was subjected to centrifugation at 600 × g for 10 min, and the bead free-cells were resuspended by stepwise addition of a 1:300 mixture of PBS: Matrigel (BD Bioscience) to a final total volume of 50 μl before injection.
Teratoma formation
The procedure of teratoma formation experiments was performed as described (16). All procedures were carried out under guidelines approved by the Institutional Review Board (SJU-2015-008) and Institutional Animal Care and Use Committee (IACUC No. SJ-20140801) of Sejong University. Male NOD/SCID mice (8~12 weeks) were used for all experiments. All procedures were carried out in a biological safety cabinet under sterile conditions using sterile materials. Injection of hESCs into testes was done under anesthesia. Mice received the following anesthetics and analgesics: Zoletil 50 (30 mg/kg)/Rompun (9.32 mg/kg). For testis injection, a sterile 1 ml syringe with a 31 G needle was injected into testes with hESC suspension (50 μl). After successful injection, the needle was retracted swiftly, the injection site was pressed with a cotton swab for about 1 min, and then a sterile, resorbable wound pad was placed at the injection site. Teratomas were surgically removed at 12~18 weeks postinjection under sterile conditions. The sizes of teratomas were measured by a ruler.
Flow cytometric analysis
Flow cytometry analysis was performed described as previous study (10). Briefly, the undifferentiated H9, RA-induced differentiated H9 cells or Colo205 cells were incubated with 20 μg of DSB-X biotin-labeled 57-C11. After isolation process described above, the cells were washed with PBS (pH7.4) supplemented with 1% bovine serum albumin (PBA). The cells were incubated with 57-C11 or biotin-labeled 57-C11 for 30 min, followed by fluorescein isothiocyanate-conjugated anti-mIgG or streptavidin-phycoerythrin, depending on the isotypes of the primary antibodies. After washing with PBA, Propidium iodide-negative cells were analyzed by FACScalibur (BD biosciences).
Results
Biotinylation and characterization of 57-C11
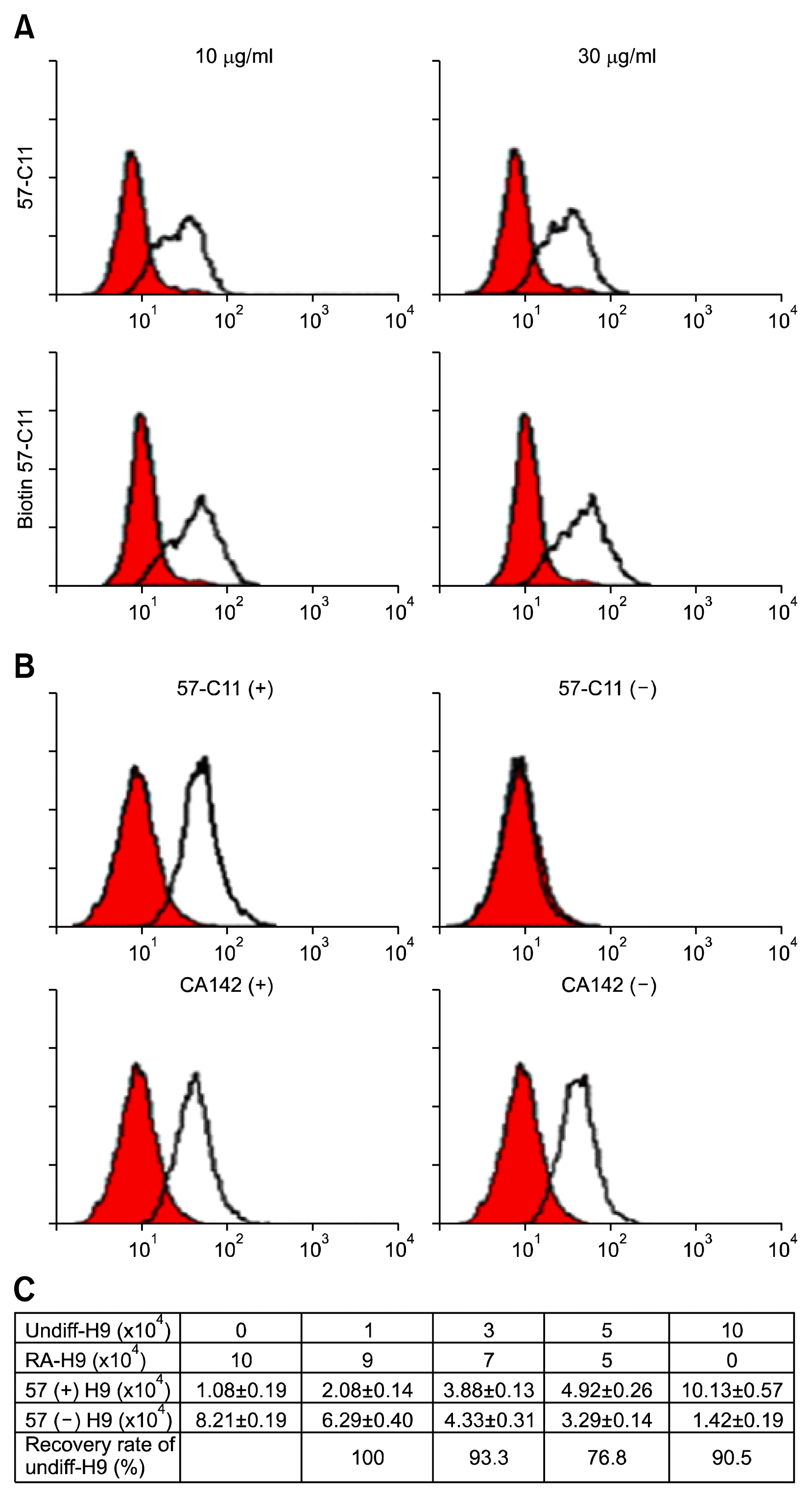
Previously, we found that 57-C11-reactive E1B-AP5 is closely associated with pluripotent and undifferentiated H9 hESCs and the surface expression of 57-C11-reactive E1B-AP5 is downregulated during the differentiation of H9 cells (11). To test whether 57-C11 is able to specifically eliminate undifferentiated H9 cells from the mixture of undifferentiated and differentiated H9 cell population, in this study, we have developed a 57-C11-mediated magnetic cell separation system, based on the specificity of 57-C11 to undifferentiated H9 cells (11). First, 57-C11 was biotinylated and 57-C11-bound H9 cells were analyzed by flow cytometry analysis with streptavidin-phycoerythrin conjugate. As shown in Fig. 1A, H9 cells (106 cells) were readily detected with biotin-labeled 57-C11 at final concentrations of 10 or 30 μg/ml. As compared with that of 10 μg/ml of biotin-labeled 57-C11, 57-C11 binding to H9 cells were not significantly increased with 30 μg/ml of biotin-labeled 57-C11, indicating that 10 μg/ml of biotin-labeled 57-C11 is enough to analyze one million hESCs. To confirm whether biotin-labeled 57-C11/streptavidin-magnetic beads were able to specifically isolate undifferentiated H9 cells from the mixed cell population, CA142 (IgG1, k), a MAb specific to surface antigen on Mycoplasma hyorhinis, was also biotinylated and used as a control isotype antibody (17). When the mixture of undifferentiated H9 hESCs and Colo205 cells were incubated with 10 or 20 μg/ml of biotin-labeled 57-C11 and subjected to the streptavidin-magnetic beads-associated isolation procedure, 57-C11-positive H9 cells were clearly separated with 20 μg/ml of biotin-labeled 57-C11 (data not shown). To reflect the possible scenario of contaminating hESC in mixed cell populations, the mixture of undifferentiated and differentiated hESCs were incubated with 20 μg/ml of biotin-labeled 57-C11 and subjected to the same isolation procedure (Fig. 2A). 57-C11-positive undifferentiated H9 cells were mostly removed with 20 μg/ml of biotin-labeled 57-C11 while the same H9 cells were not removed with the irrelevant antibody CA142 (data not shown and Fig. 1B). Thus, it seems that 57-C11/streptavidin beads is able to specifically isolate 57-C11-positive undifferentiated hESCs from the mixed cell population.
Evaluation of separation efficiency of 57-C11-positive hESCs
To examine the applicability and efficiency of this separation strategy, we prepared several pools of undifferentiated and differentiated H9 cells at different ratios (0, 10, 30, 50, or 100% undifferentiated H9 hESCs) and performed the same 57-C11-mediated isolation procedure (Fig. 2A). Approximately 10% of hESCs were recovered from 1×105 cells of differentiated H9 cells after the isolation procedure, suggesting that some cells are isolated from the differentiated H9 cells during the isolation procedure (Fig. 1C). Approximately 91% of hESCs were recovered from 1×105 cells of undifferentiated H9 cells after the isolation procedure (Fig. 1C). Our previous studies showed that approximately 89~96% of undifferentiated H9 cells were 57-C11-positive (11). Therefore, it seems that undifferentiated hESCs are almost completely recovered through the isolation procedure. When undifferentiated and differentiated hESCs were mixed at different ratios, approximately 77~100% of undifferentiated H9 hESCs were recovered after the isolation procedure (Fig. 1C). Thus, the spiking experiments suggest that the recovery rate of 57-C11-positive hESCs from mixed cell populations is approximately 77~100%.
Teratoma formation of 57-C11 positive hESCs in NOD/SCID mice
Before we validated whether isolated 57-C11-positive H9 cells were tumorigenic in NOD/SCID mice, we tested how many undifferentiated hESCs were able to form teratomas in NOD/SCID mice. Undifferentiated hESCs rapidly undergo apoptosis after the preparation of single cell suspensions from the colonies. Therefore, more than one million hESCs are recommended to use for teratoma formation in immunodeficient mice (16, 18). When we injected with 1×105, 4×105, 1×106, or 2×106 cells of undifferentiated and RA-differentiated H9 cells into the left and right testes of the same mice, respectively, we found that undifferentiated H9 cells could form teratomas with 4×105 cells under our experimental condition 8~18 weeks after injections while RA-differentiated H9 cells were not able to do (Fig. 3).
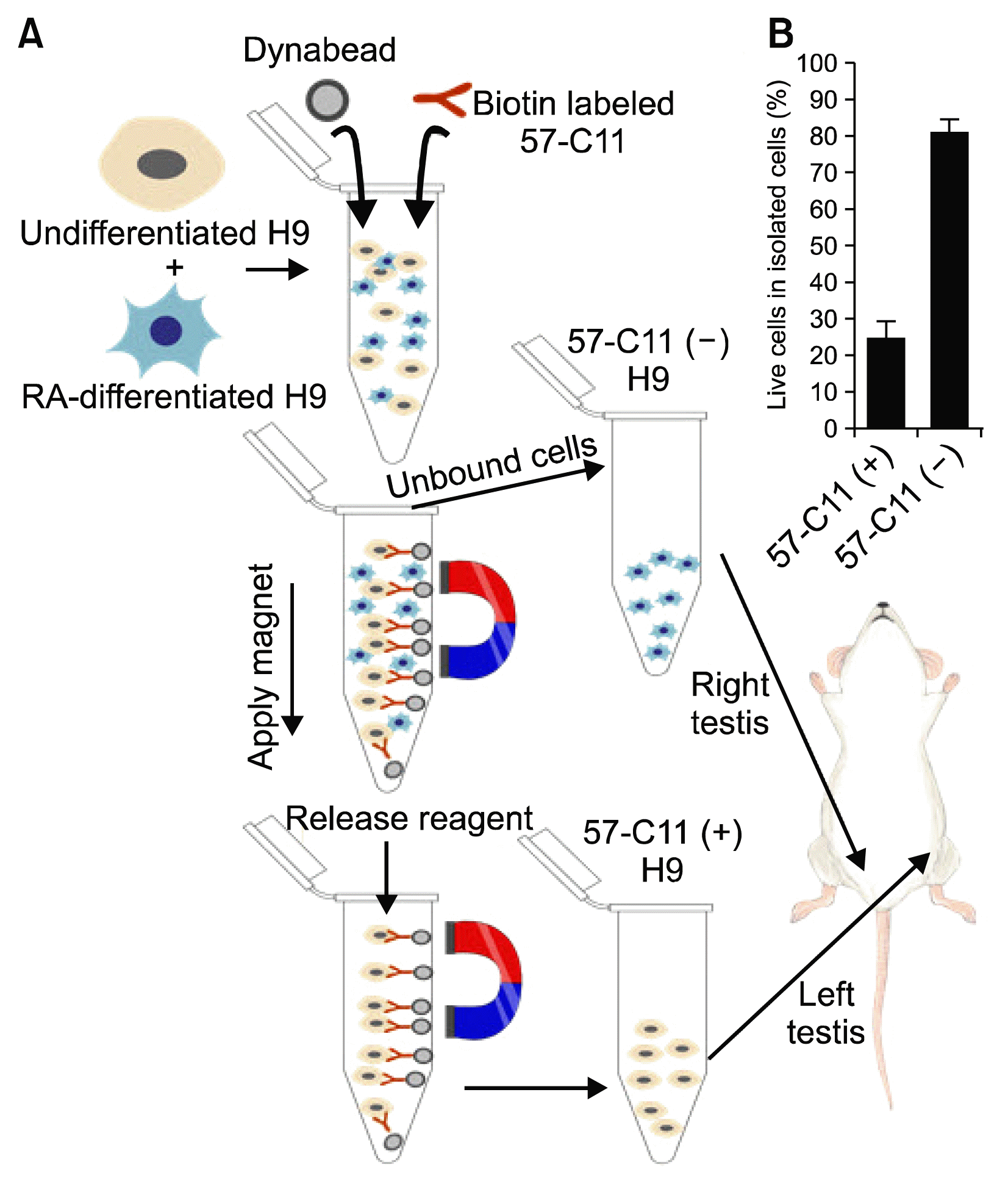
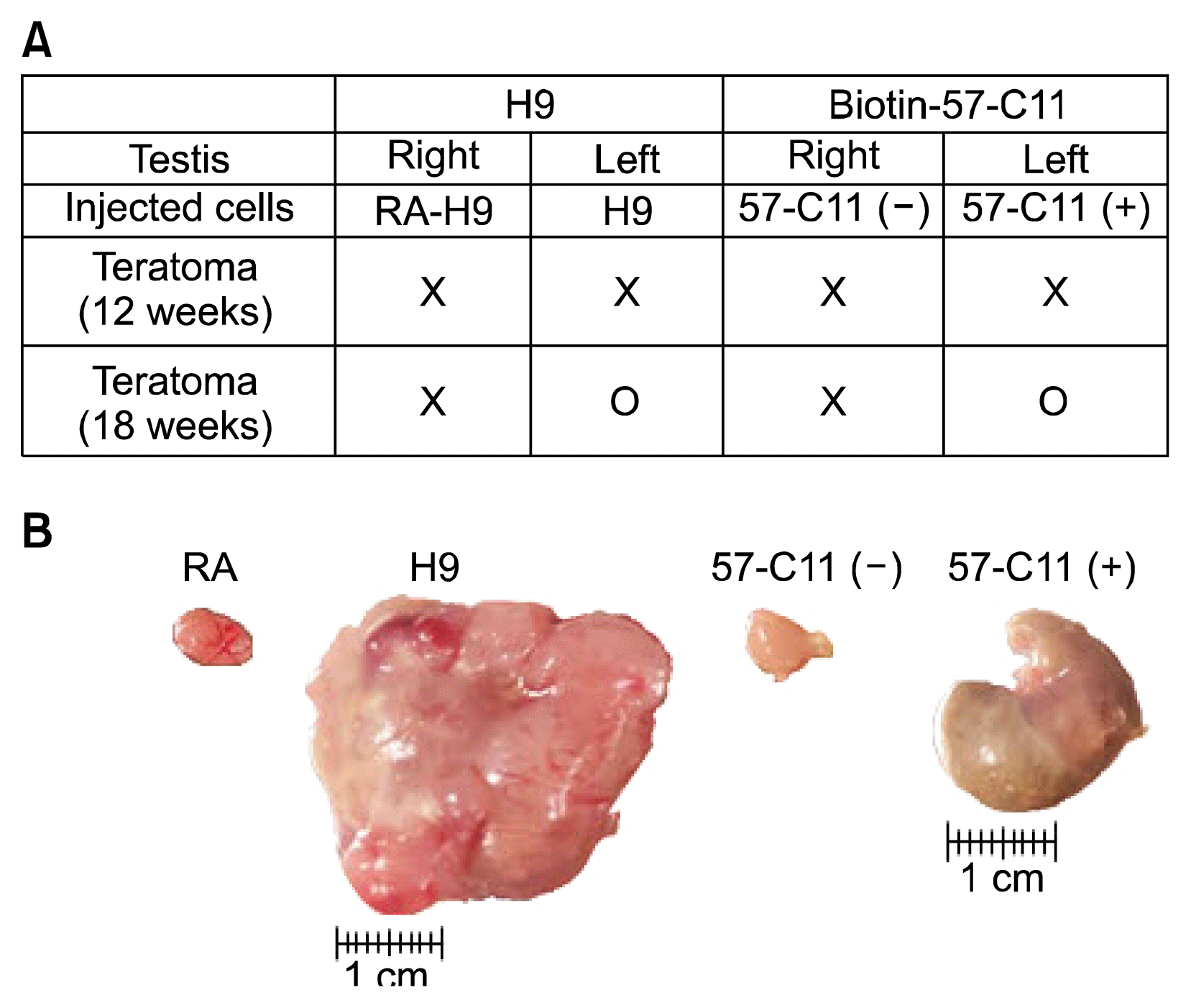
The overall scheme of the experiment including magnetic cell separation of 57-C11-positive and -negative H9 hESCs from mixed cell populations and teratoma formation of isolated H9 hESCs in NOD/SCID mice is shown in Fig. 2A. Before the injection of 57-C11-positive and -negative hESCs into NOD/SCID mice, the viability of isolated H9 cells were tested. After the isolation procedure, approximately 85% of 57-C11-negative cells remained alive while only 25% of 57-C11 positive cells remained alive (Fig. 2B). It seems that the releasing processes from magnetic beads are harmful to 57-C11-positive H9 cells. Therefore, the viability of injected hESCs were checked before injection and 1×106 cells of 57-C11-positive and negative hESCs were injected into left and right testes of NOD/SCID mice. Untreated and RA-differentiated H9 cells were also injected into NOD/SCID mice as a control experiment in the same way. When the first set of mice were opened at 12 weeks postinjection, teratoma formation was not observed even with untreated H9 cells (Fig. 4A). When the second set of mice were opened at 18 weeks postinjection, teratoma formation was observed with 57-C11-positive H9 cells or untreated H9 cells whereas it was not observed with 57-C11-negative H9 cells or RA-treated H9 cells (Fig. 4A and 4B). The results suggest that 57-C11-positive hESCs are undifferentiated and pluripotent hESCs whereas 57-C11-negative hESCs are not.
Discussion
Previously, we generated the MAb 57-C11 that recognizes the phosphorylated form of E1B-AP5 on the surface of undifferentiated hESCs (10, 11). Further studies revealed that the expression of 57-C11-reactive E1B-AP5 has a close relationship with pluripotent marker expression and is downregulated during the differentiation of hESCs (11), suggesting that 57-C11-reactive E1B-AP5 will be a specific surface marker for undifferentiated hESCs. To prove whether 57-C11-reactive E1B-AP5 is a specific surface marker for undifferentiated hESCs in vivo, we isolated 57-C11-positive and -negative hESCs by magnetic separation system and tested the capacity of teratoma formation in NOD/SCID mice. The results showed that 57-C11-positive hESCs had capacity to form teratoma whereas 57-C11-negative hESCs had not, suggesting that 57-C11-positve hESCs are really pluripotent and tumorigenic. The result also suggests that 57-C11 would be a good tool to separate remaining undifferentiated hESCs from the mixed cell population.
In the first set of injection into 2 NOD/SCID mice, teratoma formation was not observed even with untreated H9 cells at 12 weeks postinjection (Fig. 4A). To generate consistent teratomas in immunodeficient mice, many researchers have injected more than one million undifferentiated hESCs. Generally, hESCs rapidly undergo apoptosis after the preparation of single cell suspensions from the colonies even with Y-27632, the selective inhibitor of Rho-associated, coiled-coil containing protein kinase (19). Therefore, we assumed that although injected cells were alive, they were damaged and were not healthy enough to form teratomas at 12 weeks postinjection. In the second set of injection with the same cell preparation, therefore, the hESCs were given more time to recover from the damage. At 18 weeks postinjection, teratomas were observed in both untreated H9 and 57-C11-positive H9 cells. However, the teratoma size of 57-C11-positive hESCs were still smaller than that of untreated hESCs (Fig. 4B). In the first step of the isolation of 57-C11-positive hESCs, 57-C11-positive hESCs were readily separated by streptavidin-magnetic beads from mixed hESC population. After further separation of 57-C11-positive hESCs from magnetic beads, however, only 25% of isolated 57-C11-positive hESCs remained alive (Fig. 3B). Therefore, it is tempting to speculate that 57-C11-positive hESCs suffer from cytotoxic damage during the releasing processes from magnetic beads. More investigation needs to safely separate captured hESCs from magnetic beads.

 XML Download
XML Download