

 PDF
PDF Citation
Citation Print
Print


Introduction
Alzheimer’s disease (AD) is the most common form of dementia among older persons. Pathognomonic hallmarks of the disease include the development of beta –amyloid (Aβ) senile plaques and deposits of neurofibrillary tangles. Thus, compounds that could interfere with Aβ formation may be potential therapeutic agents for treatment of AD (1).
Thymoquinone (TQ) is the main constituent of Nigella Sativa (black seed) oil with many pharmacological properties including anti-inflammatory, anticonvulsant, anti-tumour and antioxidant activity (2).
The primary therapeutic strategy for treating the cognitive dysfunction in AD has been cholinergic replacement strategy, based on researches which indicated that cholinergic neurons in the forebrain support information processing and cognition which become compromised with age especially in AD. Moreover, both nicotinic and muscarinic acetylcholine receptors are considered important therapeutic targets for improving cognition in AD (3).
A novel α7 nicotinic acetyl choline receptor (α7 nAChR) selective agonist have been identified to enhance the cognitive performance. PNU- 282987 has been shown to be a potent and most specific α7 nAChR agonist. Moreover, PNU had significant effects on memory thus improving performance (4).
An alternative treatment strategy via compounds known as nicotinic “positive allosteric modulators” (PAMs) has been reported. PAM of α7 nAChRs is known as PNU-120596 (3).
The present study aimed at investigating the combination of PAM of α7 nAChRs with PNU- 282987 (α7 nAChR agonist) OR with TQ as a possible treatment for AD in an animal model using histological, histochemical, immunohistochemical and morphometric methods.
Go to : 
Materials and Methods
Drugs and chemicals
Lipopolysaccharide (LPS) was obtained from (Sigma Aldrich, Germany) in the form of powder (1g vial) dissolved in phosphate buffered saline.
Thymoquinone (TQ) was obtained from (Sigma Aldrich, Germany) in the form of yellow crystals (1g vial) dissolved in tween 80.
PNU-282987 (α7 nAChR agonist) was obtained from (Abcam Biochemicals, USA) in the form of powder (10 mg vial) dissolved in phosphate buffered saline.
PNU-120596 (α7 allosteric modulator) was obtained from (Abcam Biochemicals, USA) in the form of powder (10 mg vial) dissolved in phosphate buffered saline.
Animals
Forty eight male albino rats aged 9 months weighing 200~250 g were used in the present study. The animals were housed in the Animal House of the German University in Cairo (GUC), under good hygienic conditions of air, temperature, fed ad libitum and allowed for free water supply. The animals were treated according to the ethical guidelines of GUC and Cairo University.
The animals were divided into four groups, kept in separate cages as follows
Group 1 (Control Group)
Included eight rats (each 2 were sacrificed with the rats of each experimental group and subgroup).
Two rats, each received 0.1 ml PBS by intraperitoneal injection (IPI) once.
Two rats, each received 0.1 ml PBS by IPI once then on the 3rd day each received 0.3 ml tween 80 by IPI for 5 days.
Two rats, each received 0.1 ml PBS by IPI once then on the 3rd day each received 0.1 ml PBS by IPI for 5 days.
Two rats, each received 0.1 ml PBS by IPI once then on the 3rd day each received 0.3 ml tween 80 and 0.1 ml PBS by IPI for 5 days.
The control rats were sacrificed 1 week from the start of the experiment.
Group 2 (LPS Group)
Included ten rats, each received LPS 0.8 mg/kg dissolved in 0.1 ml PBS by IPI once (5).
Group 3 (LPS and TQ Group)
Included ten rats, each was given LPS in the same dose, by the same route and for the same duration as in group 2. On the 3rd day TQ was given by IPI (10 mg/kg) dissolved in 0.3 ml tween 80 (2) for 5 consecutive days for each rat.
Group 4 (LPS and PNU-120596 Group)
Twenty rats, each received LPS in the same dose, by the same route and for the same duration as in group 2. On the 3rd day PNU-120596 was given by IPI (1 mg/kg) dissolved in 0.1 ml PBS (6) for 5 consecutive days for each rat.
The animals were further subdivided into 2 subgroups according to drug therapy:
Subgroup 4a
Ten rats, each given PNU-282987 in addition by IPI (1 mg/kg) dissolved in 0.1 ml PBS for 5 consecutive days for each rat (7).
Subgroup 4b
Ten rats, each given TQ in addition as in group 3.
In all experimental groups and subgroups, the rats were sacrificed one week from the start of the experiment.
The rats belonging to experimental groups, subgroups and corresponding control rats were sacrificed by cervical dislocation (8) following ether anaesthesia. The skull was opened by bone cutter at the temporal regions. The brain was exposed and immediately dissected out, the frontal lobes were separated following placing in 10% formol saline for 48 hours at room temperature. Frontal lobe specimens were embedded into paraffin wax. Coronal sections of 5 μm thickness were prepared and subjected to the following studies:
Immunohistochemical Study
Anti-phospho-cyclic adenosine phosphate (cAMP) response element binding protein (P-CREB) (11) immunostaining, a beta transcription factor that activates target genes through cAMP response elements, CREB plays a key role in neuronal survival, precursor proliferation and neuronal differentiation. Primary antibody (Ab) rabbit polyclonal supplier (Merck Milipore, Germany) catalog number (06-519) used as 0.1 ml diluted stored at 2~8°C.
CD44 Ab to detect the endogenous MSCs (12). 7 ml prediluted rabbit monoclonal 1ry Ab (IW-PA1021) (IHC World, Ellicott City, USA) stored at 2~8°C.
The primary Ab was applied to the sections for 60 minutes at RT for both markers. The +ve tissue control was a specimen of brain tissue for P-CREB and a specimen of human tonsil for CD44. P-CREB +ve cells showed nuclear reaction while CD44 +ve cells showed membranous reaction. The −ve tissue control was processed in the same way, but omitting the step of 1ry Ab.
Morphometric Study
Using Leica Qwin 500 LTD (Cambridge UK) computer assisted image analyzer. The area of deformed neurons, glial cells and that of amyloid plaques were measured. The measurements were done in 10 high power fields (HPF) using interactive measurements menu. Area % of P-CREB +ve nuclei and CD44 +ve cells were measured. The measurements were done in 10 HPF using binary mode.
Statistical analysis
Quantitative data were summarized as means and standard deviations (SD) and compared using one-way analysis-of-variance (ANOVA). p-values<0.05 were considered statistically significant. Calculations were made on social package of statistical study software version 16 (13).
Go to :
Results
None of the animals died during the experiment. Moreover, gross examination of the brains from all rats at the end of the experiment showed no apparent lesions.
Histological results
Sections in the frontal lobe of control group (Group 1) demonstrated multiple pyramidal neurons with typical main dendrite in the external pyramidal layer of the cerebral cortex. Few stellate neurons with dendrites projecting for short distances in all directions and few granule cells with pale nuclei and minimal cytoplasm were also seen. Few glial cells having elongated darkly stained nuclei were evident in the neuropil (Fig. 1A). In LPS administered group (Alzheimer induced group) (Group 2), some acidophilic masses exhibiting dark nuclei together with multiple deformed neurons, few pyramidal neurons, few granule neurons and multiple glial cells were detected (Fig. 1B). In LPS and TQ administered group (Group 3), acidophilic masses were less numerous and deformed neurons became few. Some pyramidal neurons, few stellate neurons and some glial cells were demonstrated (Fig. 1C). In LPS, PNU-282987 (Alpha7receptor agonist) and PNU-120596 (Alpha7allosteric modulator) administered subgroup (Subgroup 4a) recruited few deformed neurons, multiple pyramidal neurons, few stellate neurons and few glial cells (Fig. 1D). In LPS, TQ and PNU-120596 administered subgroup (Subgroup 4b), few small acidophilic masses with dark nuclei were detected. Few deformed neurons, multiple pyramidal neurons, few stellate neurons and few glial cells were also demonstrated (Fig. 1E).
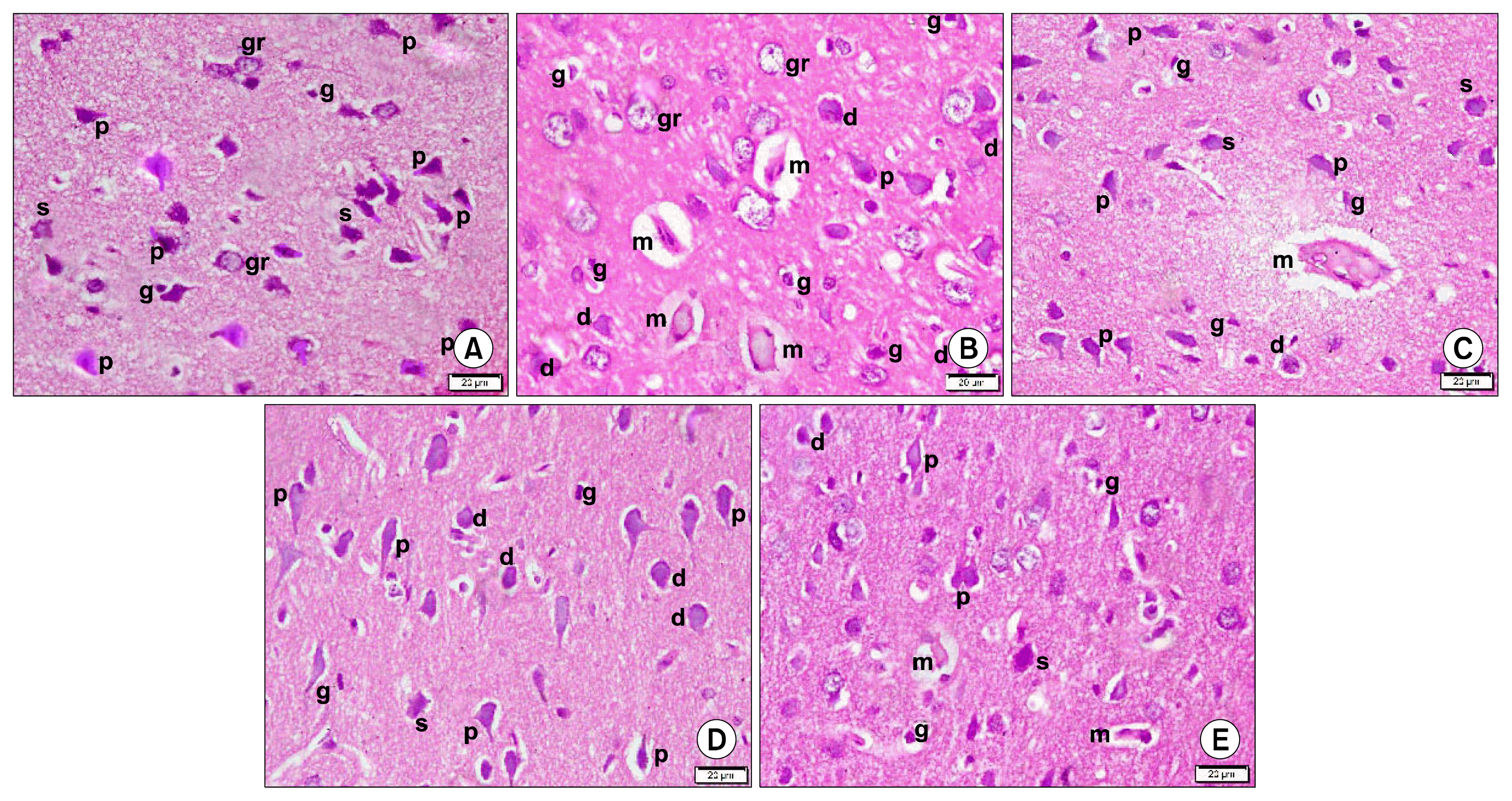 | Fig. 1Photomicrographs of sections in the frontal area of cerebral cortex (EPL) (H&E, ×400). (A) of a rat in group 1 showing multiple pyramidal (p), few stellate (s) and few granule (gr) neurons. Note few glial cells (g) in neuropil. (B) of a rat in group 2 showing 4 acidophilic masses (m) exhibiting dark nuclei and surrounded by a clear space. Multiple deformed (d), few pyramidal (p) and granule (gr) neurons are seen. Note multiple glial cells (g). (C) of a rat in group 3 showing an acidophilic mass (m) exhibiting dark nuclei and surrounded by a clear space. Few deformed (d), some pyramidal (p) and few stellate (s) neurons are seen. Note some glial cells (g). (D) of a rat in subgroup 4a showing few deformed (d), multiple pyramidal (p) and few stellate (s) neurons. Note few glial cells (g). (E) of a rat in subgroup 4b showing 2 small acidophilic masses (m) exhibiting dark nuclei and surrounded by a clear space. Few deformed (d), multiple pyramidal (p) and few stellate (s) neurons are seen. Note few glial cells (g).
|
Histochemical results
Congo red stained sections of group 1 revealed dull staining of neurons and neuropil (Fig. 2A). Group 2 demonstrated some strongly +ve masses (Fig. 2B), while group 3 showed few small +ve masses (Fig. 2C). Subgroup 4a was comparable to control with dull staining (Fig. 2D), while subgroup 4b showed accidental small +ve masses (Fig. 2E).
 | Fig. 2Photomicrographs of sections in the frontal area of cerebral cortex (EPL) (CR ×200). (A) of a rat in group 1 showing dull Congo red (CR) staining of neurons (N) and neuropil (n). (B) of a rat in group 2 showing 4 CR +ve masses (arrowheads). (C) of a rat in group 3 showing 3 small CR +ve masses (arrowheads). (D) of a rat in subgroup 4a showing dull Congo red (CR) staining of neurons (N) and neuropil (n). Note multiple blood vessels (v). (E) of a rat in subgroup 4b showing a small CR+ve mass (arrowhead) (CR ×200).
|
Immunohistochemical results
P-CREB immunostained sections of group 1 showed multiple +ve nuclei of neurons (Fig. 3A), while in group 2 few +ve nuclei were detected (Fig. 3B). On the other hand, group 3 showed some +ve nuclei (Fig. 3C), subgroups 4a and 4b showed multiple +ve nuclei (Figs. 3D and 3E).
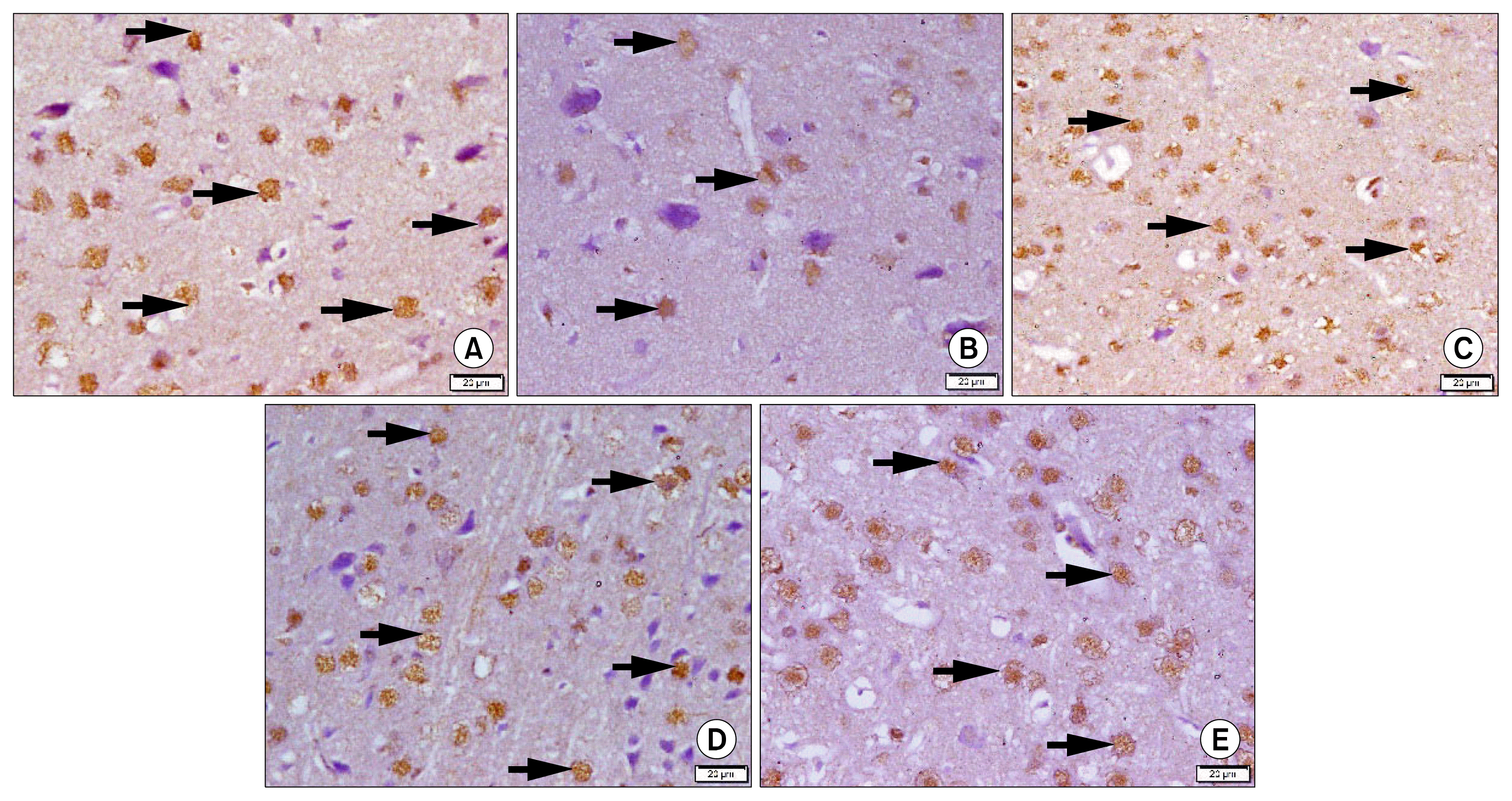 | Fig. 3Photomicrographs of sections in the frontal area of cerebral cortex (EPL) (P-CREB immuno-staining, ×400). (A) of a rat in group 1 showing multiple +ve nuclei of neurons (arrows). (B) of a rat in group 2 showing few +ve nuclei of neurons (arrows). (C) of a rat in group 3 showing some +ve nuclei of neurons (arrows). (D) of a rat in subgroup 4a showing multiple +ve nuclei of neurons (arrows). (E) of a rat in subgroup 4b showing multiple +ve nuclei of neurons (arrows).
|
CD44 immunostained sections revealed −ve immunostaining in sections of group 1 (Fig. 4A). Group 2 showed some CD44 +ve spindle cells in the neuropil and within blood vessels (Fig. 4B). Sections of group 3 recruited multiple +ve spindle cells in the neuropil and fused with deformed neurons (Fig. 4C). Subgroup 4a showed few +ve spindle cells in the neuropil and fused with accidental deformed neurons (Fig. 4D). In subgroup 4b multiple +ve spindle cells were detected in the neuropil (Fig. 4E).
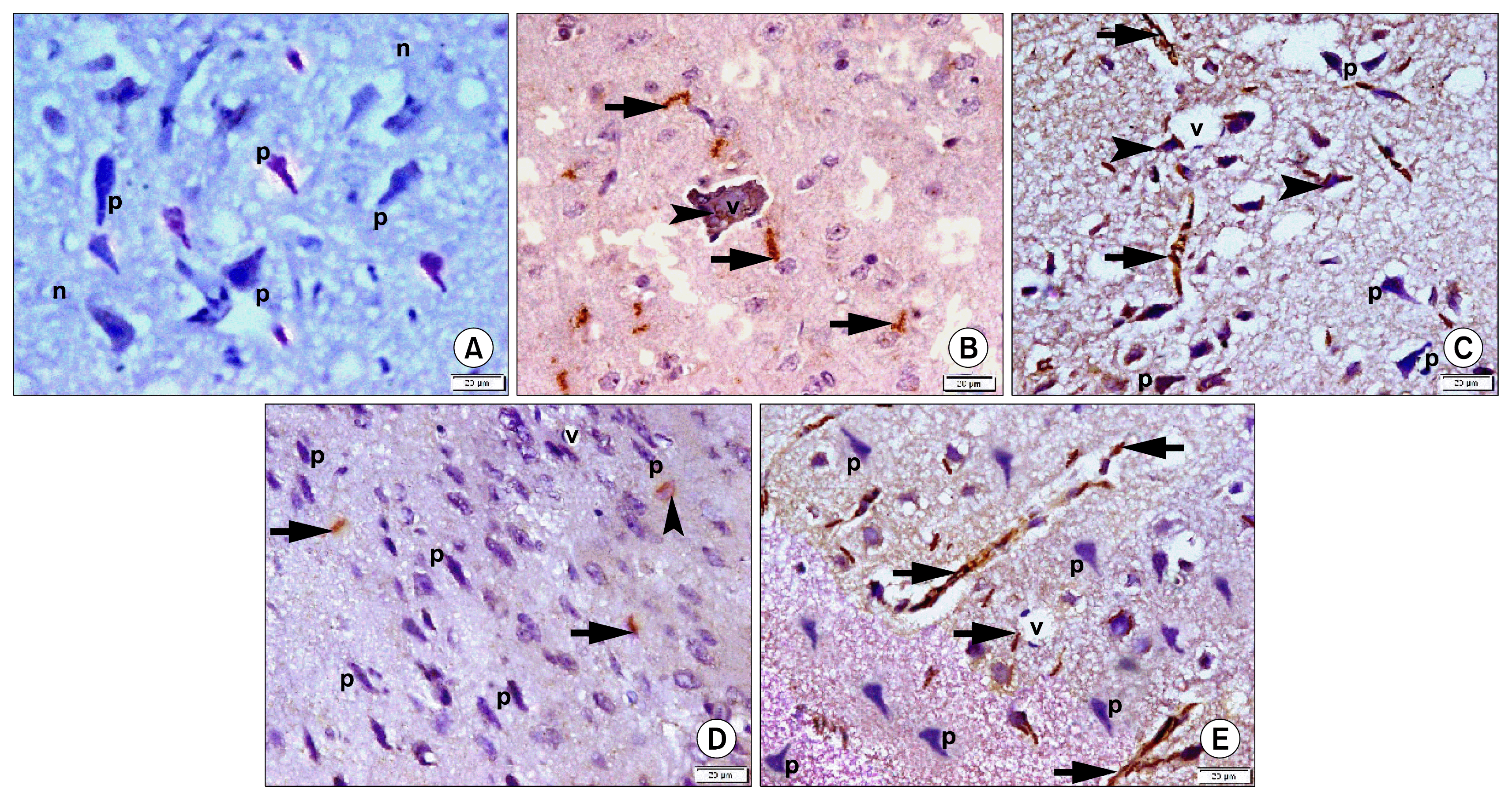 | Fig. 4Photomicrographs of sections in the frontal area of cerebral cortex (EPL) (CD44 immuno-staining, ×400). (A) of a rat in group 1 showing −ve immunostaining of pyramidal neurons (p) and neuropil (n). (B) of a rat in group 2 showing some +ve spindle cells around (arrows) and inside (arrowhead) a blood vessel (v). (C) of a rat in group 3 showing multiple +ve cells in neuropil (arrows) and fused with neurons (arrowheads). Note some pyramidal neurons (p). (D) of a rat in subgroup 4a showing few +ve cells in neuropil (arrows) and fused with a neuron (arrowhead). Note multiple pyramidal neurons (p) and a vessel (v). (E) of a rat in subgroup 4b showing multiple +ve cells in neuropil (arrows) and multiple pyramidal neurons (p).
|
Morphometric results
Table 1: Area of deformed neurons, area of glial cells, area of plaques, area% of P-CREB and area % of CD44 immunoexpression in control and experimental groups±SD.
Table 1
Area of deformed neurons, area of glial cells, area of plaques, area% of P-CREB and area % of CD44 immunoexpression in control and experimental groups±SD
| Group | Area of deformed | Area of glial cells | Area of plaques | Area % of | Area % of |
|---|---|---|---|---|---|
| neurons | +ve P-CREB | +ve CD44 | |||
| Group 1 | - | 0.76±0.09 | - | 25.01±6.52 | - |
| Group 2 | 7.54±1.72 | 3.81±0.51# | 733.16±31.23 | 8.07±1.62$ | 2.98±0.53 |
| Group 3 | 3.31±0.49* | 0.94±0.12 | 148.44±18.01^ | 17.39±4.08 | 6.44±0.92@ |
| Subgroup 4a | 1.98±0.12** | 0.74±0.05 | - | 26.08±3.22 | 1.67±0.21 |
| Subgroup 4b | 1.81±0.05** | 0.80±0.20 | 68.74±5.94^^ | 20.75±4.22 | 7.36±1.05@ |
![]()
Go to :
Discussion
Despite decades of laboratory and clinical research, AD is still the leading cause of dementia with poor therapeutic effects of the current treatments (14). This necessitates the exploration of new therapeutic strategies. Thus, the current work was carried out to investigate the combined treatment in LPS-induced AD at histological level.
In the present work, frontal lobe was selected as it was reported to play a crucial role in cognition which is known to be affected in AD (15). The current work used lipopolysaccharide (LPS) to induce AD. There are several models of AD based on neuroinflammation which is considered the key component in the neurodegenerative process of AD. LPS-induced neuro-inflammation represents the current standard paradigm to study neuro-inflammation both in vivo and in vitro (16).
Group 2 revealed acidophilic masses exhibiting dark nuclei, multiple deformed neurons, few pyramidal neurons with multiple glial cells. These findings are supported by significant increase in area of deformed neurons and that of glial cells in comparison to other groups. The observations are indicative of apoptosis and neuronal degeneration. The degenerating neurons are referred to as red dead neurons with eosinophilic cytoplasm and small shrunken nuclei. The acidophilic masses with dark nuclei might represent apoptotic neuronal cells that fuse together and form masses. Pyramidal neurons in EP layer are the principal neurons that mediate many cognitive functions and they appear to be the main substrate for neurodegenerative changes in AD (12). Glial cells increase secondary to the inflammatory process and as an attempt for clearing the degenerated neurons, supported by Nazem et al. (16).
The current study used CR stain to demonstrate amyloid Aβ plaques. It was confirmed that CR is used for identification of beta sheet nature of amyloid plaques (17). Group 2 demonstrated the presence of some CR positive plaques that were absent in subgroup 4a and became fewer in group 3 and subgroup 4b. Increase in the mean area of plaques was found in group 2. In accordance, Aβ aggregation has been identified as one of the major causes of neurodegenerative processes in AD (18) and were demonstrated in cerebral sections of AD induced in mice (19).
Phospho-CREB immunostaining was carried out in the current work. P-CREB is a constitutively expressed nuclear factor (20). It was added that P-CREB signaling is essential for learning and memory (21). On the other hand, group 2 revealed few +ve nuclei of neurons, proved by a significant decrease in the area% of P-CREB immunostaining. This might be explained on the basis of LPS induced neuroinflammation and accompanied oxidative stress followed by neuron degeneration. Hand in hand, it was reported that oxidative stress induced down regulation of P-CREB in cultured rat neurons. Moreover, inverse correlation between Aβ and P-CREB protein levels was proved (20). The decreased CREB immunostaining in the current model of AD prove and reinforce the histological findings of H&E stained sections indicative of apoptosis.
In the present work, group 2 demonstrated some CD44 +ve spindle cells in neuropil and in blood vessels. This could be explained as an attempt of MSC for healing of neurodegeneration, being attracted by inflammatory mediators and products of damaged tissue. The inflammatory mediators and the changes in the microenviroment in AD result in mobilization and differentiation of MSC that can be tissue residents or recruited from the bone marrow (12).
The current work investigated the possible role of Thymoquinone in an animal model of AD. The acidophilic masses, the deformed neurons and the glial cells became few, while the pyramidal cells became more apparent. Few CR +ve masses, some P-CREB+ve nuclei and multiple CD44 +ve cells in neuropil were detected. The previous findings were confirmed by a significant decrease in mean area of deformed neurons and that of plaques with significant increase in mean area of CREB and CD44+ve cells as compared to group 2. The recorded improvement at histopathological level could be attributed to the anti-inflammatory and antioxidant effect of thymoquinone that might interfere with neuro inflammation, Aβ plaques formation, aggregation and consequently neurodegeneration. It was reported that oxidative stress is a common mechanism contributing to neuronal damage (22). TQ was reported to inhibit Aβ aggregation in vitro and confer neuroprotection to cultured rat cortical neurons (23). Hand in hand with the current findings, antioxidant and anti-inflammatory activities of TQ were proved through inhibition of lipid peroxidation and dramatic reduction the of malondialdehyde level in cerebral cortex of rats chronically exposed to toluene (2). In addition, TQ was recorded to reduce apoptosis by reducing activity of caspase-8 and 9 (18).
Subgroups 4a and 4b demonstrated few deformed neurons, CR+ve masses were absent in subgroup 4a, small and accidental in subgroup 4b, this was confirmed by a significant decrease in the mean area of both compared to groups 2 and 3. Glial cells mean area was also reduced. On the other hand, mean area of P-CREB +ve nuclei was increased in relation to group 2. The demonstrated findings could suggest the efficiency of combined treatment in AD model. These findings could be explained on the basis of stimulation of immunologic response by α7 nAChR agonist with reduction of inflammation and consequently neurodegeneration.
Group 4 of the current work was created to evaluate the combined treatment that, to the best of our knowledge, was designed primarily and wasn’t tested before. Group 4a demonstrated the combined therapy of PNU-282987 (α7 nAChR agonist) and PNU-120596 (α7 nAChR allosteric modulator) also termed +ve allosteric modulator (PAM). While group 4b demonstrated the combined administration of Thymoquinone and PNU-120596 (PAM).
The use of PNU-282987 was based on the knowledge that α7 nAChR is expressed in key brain regions, the pre and frontal cortex (24). Moreover, It is expressed on immune cells such as microglia (25) and was significantly reduced in patients of AD (26). The combination of PNU-120596 (PAM) with either PNU-282987(agonist) or TQ was in consistent with the demonstration that PNU-120596 when combined with donepezil, it increases the drug effective dose range. Moreover, PNU-120596 restored the agonist response of desensitized receptors (3).
Selective α7 nAChR agonists demonstrated neuroprotective effects in primary neuronal cultures and rat pheochromocytoma cells PC12. Moreover, PAM affected both potency and efficacy of the agonist (27).
It was demonstrated that α7 nAChR binds to Aβ peptide (24), the interaction between them induces neuron apoptosis and reduction of α7 nAChR expression (26). So we could suggest that the agonist and the modulator together interfere with α7 nAChR /Aβ binding thus leaving the receptor free for signaling and hence the desirable effect at histopathological level. This suggestion is supported by the demonstration that the PAM allosteric modulators aimed at unleashing the factors that hinder agonist mediated α7 nAChR channel activation (1).
Group 3 and subgroup 4b revealed a significant increase in the mean area% of CD44+ve cells compared to group 2 and subgroup 4a, indicating activation of migration of MSCs from the bone marrow to the frontal area of cerebral cortex. The related apparently improved histological picture of group 4b in relation to group 3 might be explained via TQ capacity to reduce Aβ aggregation, as previously mentioned, thus α7 nAChR are free for signaling and activity, hence could be targeted by the modulator. Thus, PAM could enhance the neuroprotective effect of Thymoquinone. The activation of migration can be supported by the study conducted by Kim et al. (28), who studied the increase in functional capacity and beneficial effect of bone marrow BM-MSCs on ischemic rat cortical neurons. Activated cell migration and appears to increase their ability to release prosurvival signals and trophic factor to rescue ischemic cortical neurons. Mohd-Ali et al. (29) confirmed that activity of BM-MSCs leads to enhanced cell proliferation, migration as well as decrease in growth arrest and apoptosis.
It can be concluded that, at histological level, the early combined treatment in AD can be more efficient than single drug treatment. TQ or α7 nAChR agonist combined with PAM can have an important role in treatment of AD that is superior to thymoquinone alone. Exceptionally, TQ single or combined with PAM proved activation of MSC. Further investigations are recommended to prove the safety and efficacy of the use of combined treatment at clinical level.
Go to :
 XML Download
XML Download