

 PDF
PDF Citation
Citation Print
Print


Introduction
With its diverse genetic and metabolic potential, the gut microbiota has a substantial influence on our health and disease (1). Beyond genetics, environmental factors predominantly shape the microbial composition (2), alteration of which is associated with various human diseases (3). Human microbiota transplantation to germ-free animals has revealed a causal role of the disease-associated human microbiota, moving from its associative role (4-9). This might suggest that the gut microbiota (i.e., human origin) can easily adapt to its surrogate host (i.e., mouse). However, microbial composition between the human donors and the recipient animals colonized with the human microbiota can be substantially different (10-12). Differences in the gut milieu such as dietary components, bile acid profiles, and anatomical and cellular differences in the intestine (13-16) might contribute to failure of the microbiota in colonizing different host species.
These specificities in choice referred to as host tropism limit our understanding of the pathologies of human pathogens. Traditionally, mouse models have been widely used to reveal pathological roles of virulence factors of human pathogens (17, 18). However, the inability of the human pathogens to colonize in non-human animal models often leads to the failure of replication of their pathologies. This includes several enteric pathogenic bacteria (i.e., Escherichia coli, Salmonella enterica, Shigella flexneri) and enteric viruses (i.e., human noroviruses, human rotaviruses) (19, 20). To overcome the host tropism, immortalized human cell lines have been used to model interactions between pathogens and the host. However, immortalized cell lines usually originate from a cancerous clone and tend to lose their original characteristics. Another drawback of immortalized cell lines is the lack of interactions among different cell types and extracellular environments, which can reduce cell or tissue-specific functions (21-23). Thus, modeling the role of pathogens in immortalized cell lines has not been optimal so far. The production of human organoids, the self-organizing, stem cell-based 3D culture systems have combated the issues of immortalized cell lines and non-human animal models since they resemble the in vivo organ architecture and landscape (24, 25).
In this review, we will describe current efforts to use human organoids to model the interactions between commensal microorganisms or pathogens and the host. Since many reviews have already described extensively the various organoids in the context of pathogen interactions (26-29), we will focus on the interactions between enteric microorganisms and gut epithelium with specific emphasis on the gut milieu.
Intestinal Organoids for Modeling Human Intestine
Since the term organoid was first used in 1987 and the discovery of leucine-rich repeat-containing G protein-cou-pled receptor 5 (Lgr5) as a marker for crypt-base columnal stem cells in 2007, Sato, Clevers, and colleagues have identified the key niche factors for the long-term culture of mouse small intestinal organoids, derived from intestinal stem cells (30, 31). These niche factors include epidermal growth factor (EGF), Noggin (a BMP signaling inhibitor), R-spondin (a Wnt signaling activator), often referred to collectively as ENR. Establishing various organoids derived from different tissues of mouse or human origin has become possible by modifying the ENR conditions (i.e., adding Wnt, a TGFβ signaling inhibitor, and a p38α MAPK signaling inhibitor for human intestinal organoids) (24, 32).
Two types of cell sources are used to establish human intestinal organoids, inducible pluripotent stem cells (iPSCs) or adult stem cells (AdSCs). These two types of organoids have their respective advantages and limitations. Establishing iPSCs-derived intestinal organoids does not require human intestinal biopsies. The iPSCs are generated from reprogramming of somatic cells (i.e., fibroblasts) by forced expression of pluripotency factors such as OCT4, SOX2, KLF4, and Myc (33). The iPSCs then need to undergo directed differentiation processes such as germ-layer specification towards endoderm, induction, and maturation into an intestinal organ type, which takes weeks to months (34). However, pluripotency factors are oncogenic or potentially oncogenic and reprogramming is often inefficient, thus leaving an epigenetic memory of somatic tissue of origin (35). In addition, the cell state of the iPSCs-derived organoids remains immature and generally lacks function (36). Similarly, these organoids do not reflect the region of interest in the intestine and also can contain mesenchymal cells as a byproduct of their differentiation (24, 37). In addition, iPSCs-derived intestinal organoids used in the most studies show small intestinal properties (38, 39). A recent study has applied the protocol inducing colonic organoids from hESCs in iPSCs derived from patients with familial adenomatous polyposis (FAP-iPSCs) harboring APC mutations (40). This study showed that FAP-iPSCs intestinal organoids have enhanced proliferation and Wnt activity, similar to those in colorectal cancers (40).
The AdSCs-derived intestinal organoids can be directly generated from human intestinal biopsies, which do not require endoderm specification (24). In addition, they retain intestinal regional identities due to the maintenance of stable epigenetic signatures such as DNA methylation throughout long-term culture (41). Thus, colon organoids can be established directly from colon tissues, different from iPSCs-derived small intestinal organoids (32). The AdSCs-derived organoids only have epithelial cell types, simplifying disease modeling but instead limiting its usage to model complex interactions among cells originated from different germ layers. Another drawback is inaccessibility to human tissues. Nevertheless, as the number and accessibility of biobanks increases, it will become easier to obtain human tissues for AdSCs-derived organoids (42).
Introducing Microorganisms into the Intestinal Organoids
Microinjection
The intestinal organoids have closed 3D structures where the apical surface of the epithelium is located inside the organoids, thus limiting the epithelial interactions with luminal contents (43). Therefore, microinjection is required to induce contacts between the microorganisms and the apical surface of the intestinal epithelium while mimicking in vivo physiology. This technique has led to support luminal growth of facultative or obligate anaerobic bacteria, such as E. coli, Clostridium difficile, or fecal matter containing complex microbiota (44-47). C. difficile, a leading cause of nosocomial antibiotic-associated diarrhea, is an obligate anaerobic bacterium in its vegetative form but oxygen tolerant via dormant spore formation (44, 48). Microinjection of anaerobic C. difficile into the lumen of intestinal organoids has been successful (45, 49). This suggests the hypoxic environment of the organoids with a sealed lumen and a mucus layer (29). However, given that the lumen of organoids is not in a perfect hypoxic state, it is difficult to sustain a stable co-culture with obligate anaerobic bacteria for a long period (43). In addition, it is hard to perform high-throughput experiment with the manually performed microinjection method; however, the recently reported system would allow us to approach high-throughput experiments (44, 50).
Reversed polarity
Co and colleagues have described a method to face the apical surface of the epithelium outside the organoids, referred to as reversed polarity while maintaining a 3D organoid structure (51). Upon removal of extracellular matrix proteins and subsequent continuous suspension culture, reversed polarity can be induced, enabling easy access to the apical side of the epithelium in the organoids (51). Moreover, these apical-out organoids can differentiate into the major intestinal epithelial cell types. Modeling infections of enteric pathogens such as Salmonella enterica serovar Typhimurium, Listeria monocytogenes, and transmissible gastroenteritis virus (TGEV) have been successful with the apical-out intestinal organoids (51, 52). Compared to laborious microinjection, this technique is relatively easy to demonstrate interactions with pathogens or bacterial metabolites (43, 51). With this approach, differential preference of each pathogen towards a polarized epithelium was identified (i.e., S. Typhimurium on the apical side, L. monocytogenes on the basal side) (51). However, it is questionable whether the reversed organoids in suspension were tolerable to study long-term interactions with pathogens. In addition, investigating anaerobes in the apical-out intestinal organoids is not feasible due to the oxygen-rich environment.
2D monolayer
3D organoids can be used to generate monolayers with the apical side facing upward (43, 53). Briefly, dissociated 3D organoids are plated as monolayers in a transwell, which enables separation between the apical and basal compartment where we can easily introduce microorgani-sms in the apical side (43, 54). However, this system is not optimal for culturing both obligate anaerobes and oxygen-requiring epithelial cells (43). To overcome this limitation, an anaerobic transwell system has been developed in which the apical chamber is anaerobic while the basal chamber is in aerobic conditions. Briefly, an anaerobic environment is generated by sealing the apical chamber with a plug and thus the epithelial monolayer could be co-cultured with obligate anaerobes such as Bifidobacteria, Clostiridium, and Akkermansia (55). However, the long-term culture of organoids as 2D monolayers generates low oxygen tension, causing cellular stress and reduced cell differentiation (56).
Air-liquid interface
Oxygenation at the apical side in 2D monolayers can be improved by removing the medium from the upper chamber while keeping the basal side submerged in the medium (57). This air-liquid interface (ALI) method can supply oxygen and thus healthy epithelial monolayers can be maintained for a long period by reducing oxygen stress. Moreover, incorporating underlying stromal elements into the ALI is possible, thus preserving the epithelial-mesenchymal interactions (58). The ALI culture system provides an accessible model to study long-term host-pathogen interactions, as evidenced by robust growth and complete development of the gastrointestinal parasite Cryptosporidium with this system (59). However, the ALI system does not form a physiological lumen (60) and to culture obligate anaerobes in the ALI culture system, the apical compartment needs to be anoxic while host cells need to be oxygenated basolaterally.
Human Intestinal Organoid as a Research Tool for Studying Host-Microorganism Interactions
Escherichia coli
Escherichia coli is one of the most well-known bacterial species, which is a member of the intestinal microbiome. Non-pathogenic E. coli produces vitamin K and B12 which provide benefits to the host, but certain pathogenic E. coli strains cause diseases (46). Co-culture of non-pathogenic E. coli with human intestinal organoids via microinjection has shown to induce hypoxia in the intestinal lumen, leading to maturation of the mucus layer, antimicrobial peptide production, and improved barrier function (47, 61). Pathogenic E. coli strains have been also applied to the human intestinal organoids, which includes enterohemorr-hagic E. coli (EHEC), a foodborne pathogen that causes bloody diarrhea and hemolytic uremic syndrome and enterotoxigenic E. coli (ETEC), the most common cause of children’s and traveler’s diarrhea in the developing countries (62, 63). Infecting pathogenic EHEC into 3D human intestinal organoids caused hypoxia and intestinal maturity, like those infected with non-pathogenic E. coli (61). However, unlike non-pathogenic E. coli, EHEC severely destroyed the epithelial barrier, induced reactive oxygen species (ROS) production, and initiated inflammatory responses (61). Infecting pathogenic E. coli including EHEC and ETEC in 2D intestinal organoid monolayers confirmed that E. coli infection occurred on the apical surface of the intestinal epithelium (54, 64). In addition, with 2D human colonoid monolayers, EHEC-induced early molecular events such as mucin layer reduction and microvillar effacement have been identified (54). Furthermore, a human enteroid-macrophage co-culture system has revealed that macrophages facing the basal side can kill luminal pathogens such as ETEC attached to the apical side across the intestinal monolayer (65).
Shigella flexneri
Shigella, an intracellular pathogen, causes intense inflammation in the colonic and rectal epithelium (66). In human intestinal organoid monolayers derived from the small intestine and colon, basolateral infection and intracellular replication of S. flexneri have been recapitulated (67, 68). S. flexneri invasion in monolayers increased NF-κB-mediated inflammation signaling pathway and production of MUC2 (26, 67, 68). However, the apical invasion of S. flexneri can be increased by M cell induction or tight junction disruption in monolayers (67, 68).
Salmonella
Salmonella, a major foodborne pathogen, causes enteric fever and acute self-limiting gastroenteritis (69). Apical infection preference of S. Typhimurium has been demonstrated in the reversed epithelial polarity organoids (51). S. Typhimurium was able to invade the epithelial barriers and increase the expression of innate immune responses, inflammation, and cytokine-mediated signals upon apical infection via microinjection (70). In addition, infection of S. Typhimurium in 2D monolayers induces transdifferen-tiation of enterocytes into functional M cells (71). However, basolateral infection of S. Typhimurium seemed to be effective in disrupting tight junctions and enhancing NF‐κB signaling in 3D murine intestinal organoids (72). Infection of another serovar strain of Salmonella enterica, S. Typhi in human intestinal organoid-derived monolayers has shown to reproduce phenotypes observed in the infected tissue such as microvilli destruction, cytoskeleton rearrangement, and vesicle-contained bacteria (69, 73).
Vibrio cholerae
Vibrio cholerae causes human diarrheal pandemic disease cholera. V. cholerae has two major virulence factors, Cholerae toxin (CT) and toxin-coregulated pili (TCP) (74). CT is sufficient to induce severe diarrhea, which is caused by CT-induced electrolyte imbalance (75). Specifically, CT binding to GM1 receptors induces the entry of CT into the epithelium and after the activation of several signal transduction pathways, cAMP levels are increased. Increased cAMP then leads to increased chloride secretion into the lumen via the activation of cystic fibrosis transmembrane receptor (CFTR) and reduced sodium uptake via the inhibition of sodium-hydrogen exchanger 3 (NHE3). This net electrolyte imbalance leads to watery diarrhea characteristic of cholera (76, 77). Therefore, as a strategy to prevent diarrhea caused by CT, blocking of GM1 binding using random polymers carrying galactose and fucose has been reported in the human enteroids (78). In vivo pathology of CT has shown to be recapitulated in human organoids such as GM1 or NHE3-dependent CT-induced swelling of organoids (79, 80). Moreover, intestinal organoids derived from O-blood group and A-blood group provided a direct clue as to why O-blood group is associated with more severe cholera infections (81).
Clostridium difficile
Clostridium difficile, the cause of severe diarrhea and colitis, can spread via the fecal-oral route (82). The pathogenicity of C. difficile is mediated by two exotoxins, TcdA and TcdB, which disrupt the cytoskeletal structure and the tight junction of the intestinal epithelium (83). In vivo pathologies of C. difficile such as epithelial barrier disruption were recapitulated in the human intestinal organoids infected with C. difficile, or injected with TcdA (44, 48). In the human jejunal enteroids, TcdA receptor levels were higher than TcdB receptors (84). Similarly, TcdA was 10-fold more effective in disrupting cytoskeletal rearrangement than TcdB in the 2D jejunal enteroid monolayers. Moreover, bacitracin, an antibiotic known to inhibit the cell wall synthesis of gram-positive bacteria, can protect human intestinal organoids from C. difficile-induced destruction of F-actin (85). This suggests that the human enteroid system is a great tool to investigate the clinical efficacy of drugs against C. difficile infections.
Norovirus
Human norovirus, the most common causative agent of acute gastroenteritis, causes vomiting and diarrhea worldwide (86). Since the discovery of norovirus, many resear-chers have attempted to cultivate human norovirus in vitro, but most efforts were unsuccessful (87, 88). Interestingly, however, norovirus can be replicated when co-cultured with human small intestinal organoid-derived monolayers (89). Moreover, human intestinal organoids are effective to identify norovirus inactivation factors (89, 90).
Rotavirus
Rotavirus is the leading cause of serious gastroenteritis in young children worldwide (91). The growth of human rotavirus is limited in most transformed cell lines and animal models. However, rotavirus can also be cultured in human and murine intestinal organoids (92, 93). Further-more, organoids infected with rotavirus can produce virus particles, and the viral replication in organoids was inhibited by interferon-alpha and ribavirin treatment (92). Importantly, human intestinal organoids were more permissive to human rotavirus infection than mouse intestinal organoids, demonstrating host specificity of human rotavirus (93).
Commensals
Since the human microbiome project began, important roles of commensal bacteria in the gut homeostasis, such as the supply of essential nutrients, metabolism of indigestible compounds, and stimulation of the immune system have been emphasized (94, 95). For example, Lactobacillus, Bifidobacterium, Akkermansia, and Eubacterium spp. have been suggested to be associated with good intestinal health (96-98). Lactobacillus spp. such as L. rhamnosus, L. reuteri, and L. plantarum have been shown to enhance maturation, proliferation, and barrier function of the intestinal organoids (99-101). In addition, Lactobacillus spp. growth was more supported by mature human intestinal organoids than immature human intestinal organoids (101). This suggests that intestinal organoids can be used to investigate host effects towards microorganisms as well as the effects of microorganisms on the intestinal epithelium.
Compared to Lactobacillus spp, facultative anaerobes, obligate anaerobic bacteria such as Bifidobacterium, Akkermansia, and Eubacterium, the dominant species in the large intestine require a hypoxic environment. Co-culture of B. adolescentis and E. hallii with Caco-2 cells has shown to be successful with the anoxic-oxic interface-on-a-chip for up to 1 week (102). However, the co-culture of human colonic organoids with obligate anaerobes has been challenging until Sasaki and colleagues established the 2D-intestinal hemi-anaerobic coculture system (iHACS), consisting of a hypoxia apical chamber and a normoxic basal chamber (55). In this iHACS system, B. adolescentis significantly increased the expression of markers for goblet and stem cells. Unlike B. adolescentis, A. muciniphila uses mucin as a carbon source instead of glucose. Differentiated human colonic organoids but not undifferentiated organoids can support the growth of A. muciniphila, which supports the role of goblet cell-derived mucin as a carbon source for this bacterium (55). This indicates that proper use of organoid systems would enable us to model competitive or beneficial interactions between human epithelium and commensals.
Points to Consider for the Proper Use of Human Intestinal Organoid to Study Host-Microorganism Interactions
Tissue tropism
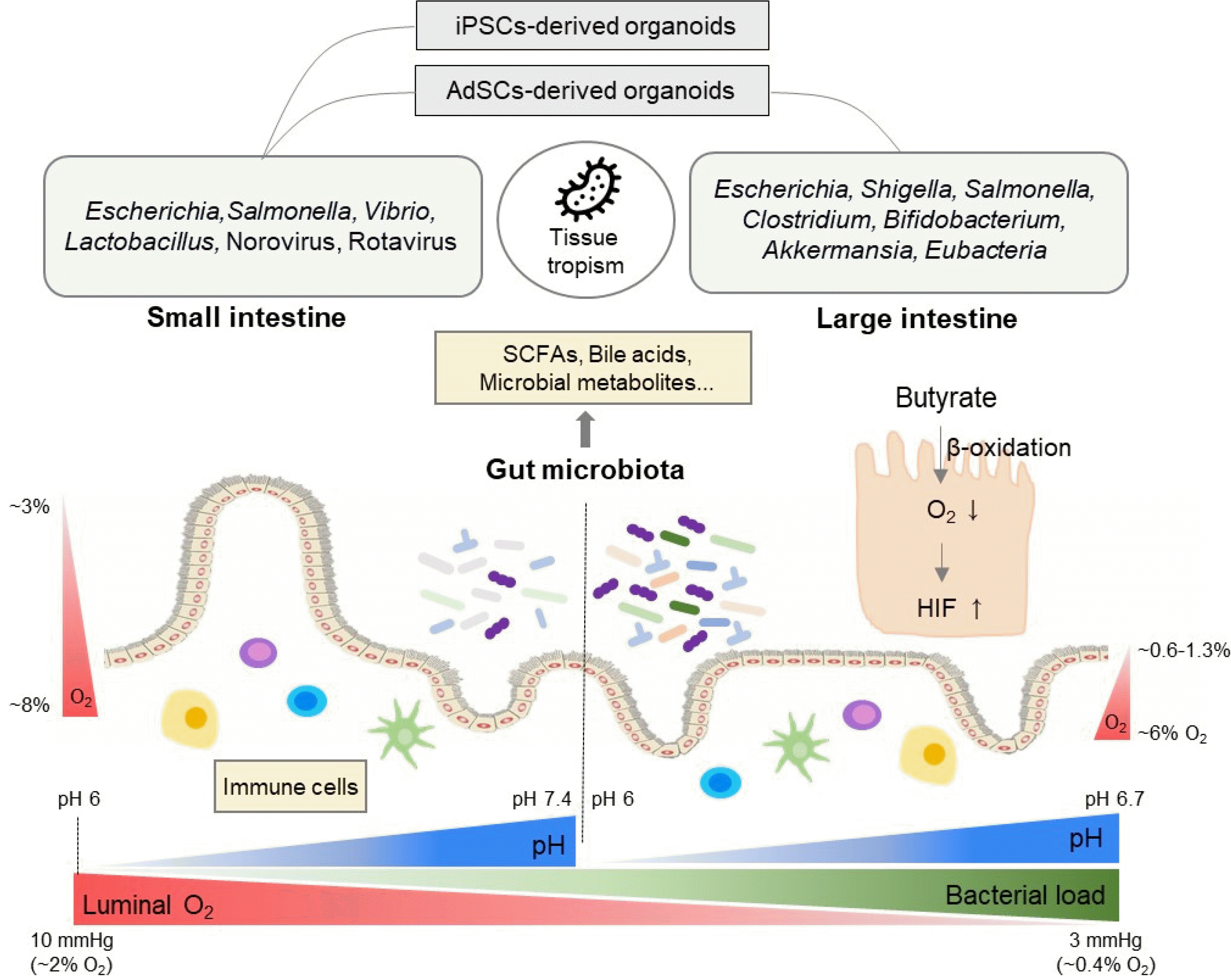
Microorganisms including pathogens often have a tissue tropism (i.e., small intestine versus large intestine) (Table 1, Fig. 1). As mentioned earlier, the AdSCs-derived intestinal organoids retain their own highly stable gut segment-specific epigenetic regional identities (41). However, the iPSCs-derived intestinal organoids might retain epigenetic memory of non-intestinal tissue of origin and showed small intestine properties in the most studies (35, 38, 39). Thus, it is necessary to choose proper intestinal organoid systems based on the regionality of microorganisms (Table 1, Fig. 1). For example, Shigella, S. Typhi, and C. difficile show the highest adhesion to the large intestine (68, 103, 104). However, some in vitro models of these pathogen infections have been established in organoids with small intestinal properties (39, 48, 68, 73, 84, 85). On the other hand, in some cases, pathogens with a preference towards the small intestine are studied in the large intestine-derived organoids (79, 81, 92, 99). If the tissue tropism of microorganisms is not considered, it might lead to wrong interpretation. For example, pathogenic enterohemorrhagic E. coli (EHEC) with preference towards the large intestine increased expression of Muc2 and tight junction proteins in the hESCs-derived small intestinal organoids whereas it reduced Muc2 expression and integrity of tight junction in human colonoids (54, 61). Similarly, EHEC infection in vivo redistributed the tight junction protein (105). Therefore, for better modeling of the interactions between human intestinal epithelium and microorganisms, the tissue tropism of microorganisms and it’s matched organoid systems should be considered.
Gut milieu
Although there has been enormous progress in the use of human intestinal organoids for modeling interactions with microorganisms, it is still at an immature stage in applying various gut environmental factors other than anaerobic conditions. Here, we will describe various gut milieu (Fig. 1), which needs to be considered in future intestinal organoid research.
Microbial fermentation activity in the gut can influence oxygen levels and pH. Short-chain fatty acids (SCFAs) such as acetate, propionate, and butyrate are products of microbial fermentation of dietary fibers (106). Primarily, SCFAs are used as energy substrates in oxidative phospho-rylation, where oxygen is consumed to produce ATP. For example, microbial butyrate limits the bioavailability of oxygen and maintains intestinal homeostasis via PPARγ-mediated β oxidation of butyrate or hypoxia-inducible factor (HIF) stabilization (107, 108). Without microbial butyrate, oxygen levels were high in the intestinal epithelium in germ-free mice, whereas butyrate supplementation restored oxygen levels in antibiotics-treated mice (108, 109). Different from epithelial oxygen levels, luminal oxygen levels in germ-free mice and conventional mice were nearly identical along the intestinal tract (110). This suggests the existence of a host-derived oxygen consumption mechanism (i.e., lipid oxidation) other than microbial respiration although the oxygen consumption rate was slower in the germ-free mice than conventionally-raised mice (110).
In addition, the oxygen concentrations of the intestine need to be considered in two directions - longitudinal and cross-sectional. First, there is a longitudinal steep decrease in the oxygen gradient from the proximal small intestine to the distal colon. Luminal pO2 in the small intestine is about 10 mmHg (∼2% O2), which decreases rapidly along the gut axis and reaches less than 3 mmHg in the sigmoid colon (∼0.4% O2) (111). Second, there is a steep decrease in the oxygen from the base of the epithelium to the lumen. In the small intestine, pO2 at the intestinal barrier is about 59 mmHg (∼8% O2), about 22 mmHg (∼3% O2) at the tip of the villus, and less than 10 mmHg (∼2% O2) in the lumen (111). In the large intestine, pO2 of the colonic muscle wall is around 42∼71 mmHg (∼6-9% O2), about 42 mmHg (∼6% O2) in the vascularized submucosa, 5∼10 mmHg (∼0.6-1.3% O2) at the crypt-lumen interface, and less than 3 mmHg (∼0.4% O2) in the lumen of the sigmoid colon (111, 112). Therefore, strict anaerobes could grow in this anaerobic environment in the large intestine (111).
Microbial fermentative products, SCFAs, also affect pH in the colon, sharply dropping to 6 in the cecum and rising back to 6.7 in the rectum (113-115). The pH of the small intestine can be affected by stomach acid (pH 1.4) and pancreatic juice (pH 8.6). The pH of the duodenum is about 6.0, which becomes 7.1 in the jejunum and 7.4 in the ileum (116, 117). The intestinal pH can be altered by intake of food or water, and these variations in pH affect microbial community and metabolism (118).
Beyond SCFAs, the gut microbiota can metabolize primary bile acids, synthesized in the liver, into secondary bile acids and the amino acid tryptophan into indole-containing compounds (119). Despite the successful replication of previously non-cultivatable human noroviruses in human organoids, not every norovirus strain can replicate. Interestingly, bile acids have been shown to induce strain-dependent norovirus replication such as GI.1, GII.3, GII.17 and enhance infectivity (89, 90, 120). In addition, microbial indole-3-propionic acid and indole-3-al-dehyde have been shown to increase the expression of IL-10 receptor ligand-binding subunit, mediating anti-inflammatory IL-10 action in human intestinal organoids (121, 122). These suggest that microbial metabolites need to be considered as important gut milieu to better mimic in vivo gut, in addition to oxygen and pH.
Another important gut milieu to consider is immune cells interacting with the intestinal epithelium and the microbiota. Immune cells such as T cells, innate lymphoid cells, dendritic cells, and macrophages have shown to affect differentiation and barrier function of the intestinal epithelium, which in turn can prevent infections (123, 124) and might affect microbial composition (125). Intestinal dendritic cells have suggested to compartmentalize the commensal microbiota via sampling of the gut bacteria for antigen presentation (126). Moreover, the immune system maintains a homeostatic relationship with microbiota by a mucosal firewall, a structural and immunological component made in cooperation with mucosal cells, IgA, antibacterial peptides, and immune cells (127). For example, a diversified and selected IgA has suggested to maintain balanced microbiome and to prevent inflammation (126). Thus, immune cells are important gut milieu when modelling the real interactions between the intestinal epithelium and commensals or pathogens.
Perspective
Human intestinal organoids have made it possible to establish models that mimic actual organs (50). Hence, it can overcome the limitations of intrinsic anatomical and cellular differences in the intestine between human and animal models and is a promising human-microorganism interaction model. However, culture conditions for intestinal organoid establishment may not match the environment for the survival of microorganisms in the gut (50, 128).
The oxygen concentration is one of the representative mismatched conditions. Intestinal oxygen concentration is controlled by microbial and epithelial oxygen consumption. In the human newborn intestine, the facultative bacteria such as Escherichia, Streptococcus, and Enterobacteriaceae, colonizing during the first 2 weeks, can consume oxygen and lower the redox potential to negative values (129, 130). This enables colonization of the obligate anaerobes such as Bifidobacterium, Clostridia, and Bacteroides (130, 131). Subsequently, saccharolytic bacterial fermentation activity of normal intestinal microbiota generates beneficial SCFAs, lowering pH. Specifically, butyrate reduces epithelial oxygenation levels via β-oxidation of butyrate and thus prevents dysbiotic expansion of aerotolerant pathogens such as V. cholerae, S. enterica, and S. flexneri (26, 132, 133).
Intestinal organoid culture conditions in vitro, however, are enriched with high glucose, but devoid of butyrate. Colonocytes in vivo use butyrate as a primary energy source, but energy sources can be converted into glucose when butyrate is depleted (107, 134). Thus, even if we add butyrate as an alternative energy source in the presence of high levels of glucose in intestinal organoid cultures, glycolysis will occur more predominantly than oxidative phosphorylation. This can lead to the accumulation of butyrate, enough to act as a histone deacetylase (HDAC) inhibitor (106, 133, 135). Indeed, butyrate and valproic acid, structurally similar to butyrate, have been shown to affect intestinal organoid stem cell function via HDAC inhibition under the typical intestinal organoid culture conditions (136, 137). Therefore, optimizing glucose and butyrate concentrations would be required to induce β-oxida-tion of butyrate and, as well as sustain functional intestinal organoid cultures where subsequently induced physiological hypoxia would generate a suitable environment for microorganisms-intestinal epithelium interactions.
Given that the gut milieu such as oxygen, pH, metabolites, and immune cells are regulated by the interaction of gut microbiota and intestinal epithelial cells, the gut microbiota transplantation into intestinal organoids may help to simulate actual intestinal physiological activity (45). However, intestinal organoid culture conditions in vitro are also enriched with amino acids, but devoid of fibers. Under the traditional organoid culture conditions, ssacharolytic bacteria cannot ferment fibers but assacharolytic bacteria can use amino acids as their carbon source, which potentially generates toxic metabolites (138, 139). It would thus be critical to use physiologically relevant culture conditions to maintain transplanted microbiota in intestinal organoids.
From the perspective of pathogens, changes in environmental signals during host infection can trigger an adaptive response to survive and affect the expression of virulence genes (140). For instance, anaerobic conditions can enhance S. flexneri invasion by the activation of the type 3 secretion system (141). Bile acids can increase the survivability and pathogenicity of some enteric pathogens (142, 143). To sum up, when studying host-pathogen interaction, efforts should be made to generate an environment in which pathogenicity can be expressed for each pathogen by reflecting the actual environment as much as possible.
 XML Download
XML Download