

 PDF
PDF Citation
Citation Print
Print


Introduction
During embryonic development, diverse types of cells are generated by a precisely sequenced process in a highly conserved manner to form the functional brain. Such a process is finely guided by spatiotemporally-restricted gene expression and regulated by intrinsic and extrinsic factors, such as cell cycle kinetics, morphogen gradient, cell-cell interactions, and cell-extracellular matrix interac-tions.
Brain development begins with the neuroectoderm specification during gastrulation. Once it achieves its neural fate, the neuroectoderm undergoes a transition into neural tube formation. Through a series of expansions and the subsequent closure of the tube, the primary brain vesicles are formed, consisting of mesencephalon and rhombence-phalon. Further specification of positional identity and regionalization is achieved by morphogen gradients (Fig. 1). For instance, the SHH and BMP gradients determine the dorsoventral identity within the neural tube, acting as a ventralizing and dorsalizing factor, respectively. Indeed, the discrete region that releases growth factors and forms the gradients of such signaling molecules is referred to the signaling center. One of the well-known examples is the rostral patterning center, in which BMP4, SHH, and FGF8 synergistically generate the rostral forebrain.
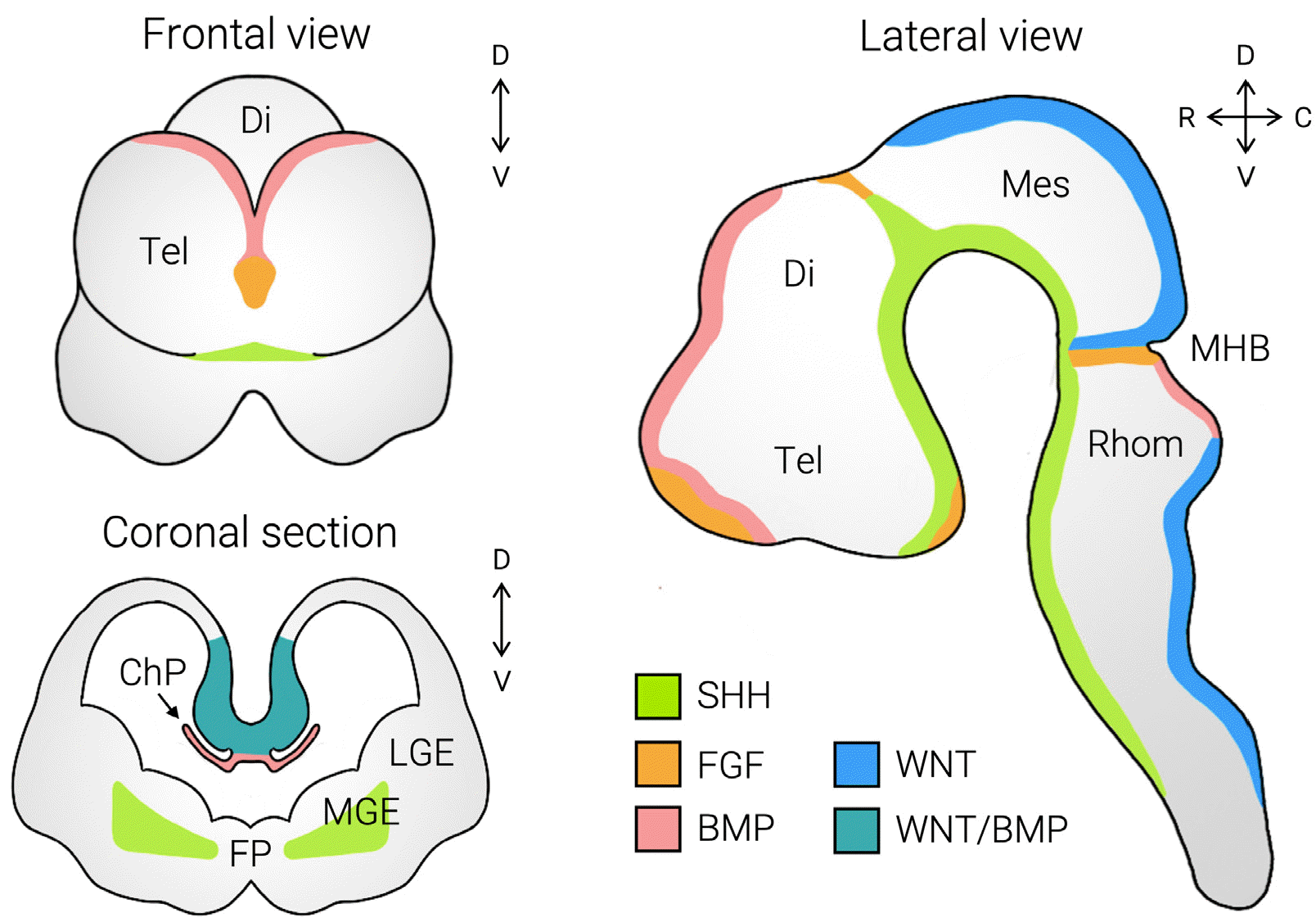 | Fig. 1Signaling dynamics in the embryonic brain development. The schematic illustration of signaling factors in mouse embryonic brain development. The combinatory effects of morphogens determine the regionalization of the brain. Tel: telencephalon, Di: diencephalon, Mes: mesencephalon, Rhom: rhombencephalon, MHB: midbrain-hindbrain boundary, ChP: choroid plexus, FP: floor plate, MGE: medial ganglionic eminence, LGE: lateral ganglionic eminence, SHH: sonic hedgehog, FGF: fibroblast growth factor, BMP: bone morphogenic protein.
|
The brain involves unique and extraordinarily complex developmental features compared to other organs. The mechanisms underlying brain development have been extensively studied, mostly on model organisms, such as mice. Although decades of work have elucidated the evolutionarily conserved biology of the developing brain and neurodevelopmental diseases, the species-specific features still hinder the complete comprehension of human brain development (1). Therefore, the need for experimental models that accurately recapitulate the developing human brain has increasingly emerged.
The establishment of human embryonic stem cells (ESCs) (2) and induced pluripotent stem cells (iPSCs) (3) granted scientists a tremendous opportunity to investigate mechanisms of normal brain development and human brain disorders. Yet, the two-dimensional (2D) monolayer culture of human pluripotent stem cells (PSCs) often suffers from a lack of architectural complexity, such as three-dimensional (3D) cell-cell interactions and segregation into a structural and functional unit as in vivo. Over the past several years, a new model system that could recapitulate the cellular and molecular characteristics of brain development and mimic the 3D-cytoarchitecture of the brain has been developed, also known as brain organoids. Brain organoids can be generated from human PSCs in a dish, enabling researchers to overcome the previous hurdles in deciphering embryonic development and avoid ethical issues related to obtaining human samples (4, 5).
Conventionally, the generation of brain organoids is mediated by the capability of the human PSC-derived embryonic body (EB) to self-assemble and self-pattern to form distinct brain regions in a 3D structure (5). A method with minimal external signaling factors to induce lineage specification, or ‘unguided differentiation,’ yields various brain regional domains, including the retina, forebrain, midbrain, and hindbrain (6). Additionally, the addition of small molecules in the culture media to direct the differentiation into a particular brain region of interest, or ‘guided differentiation,’ has been developed, such as the forebrain, midbrain, hypothalamus, hippocampus, and cerebellum organoid (Fig. 2) (7-9).
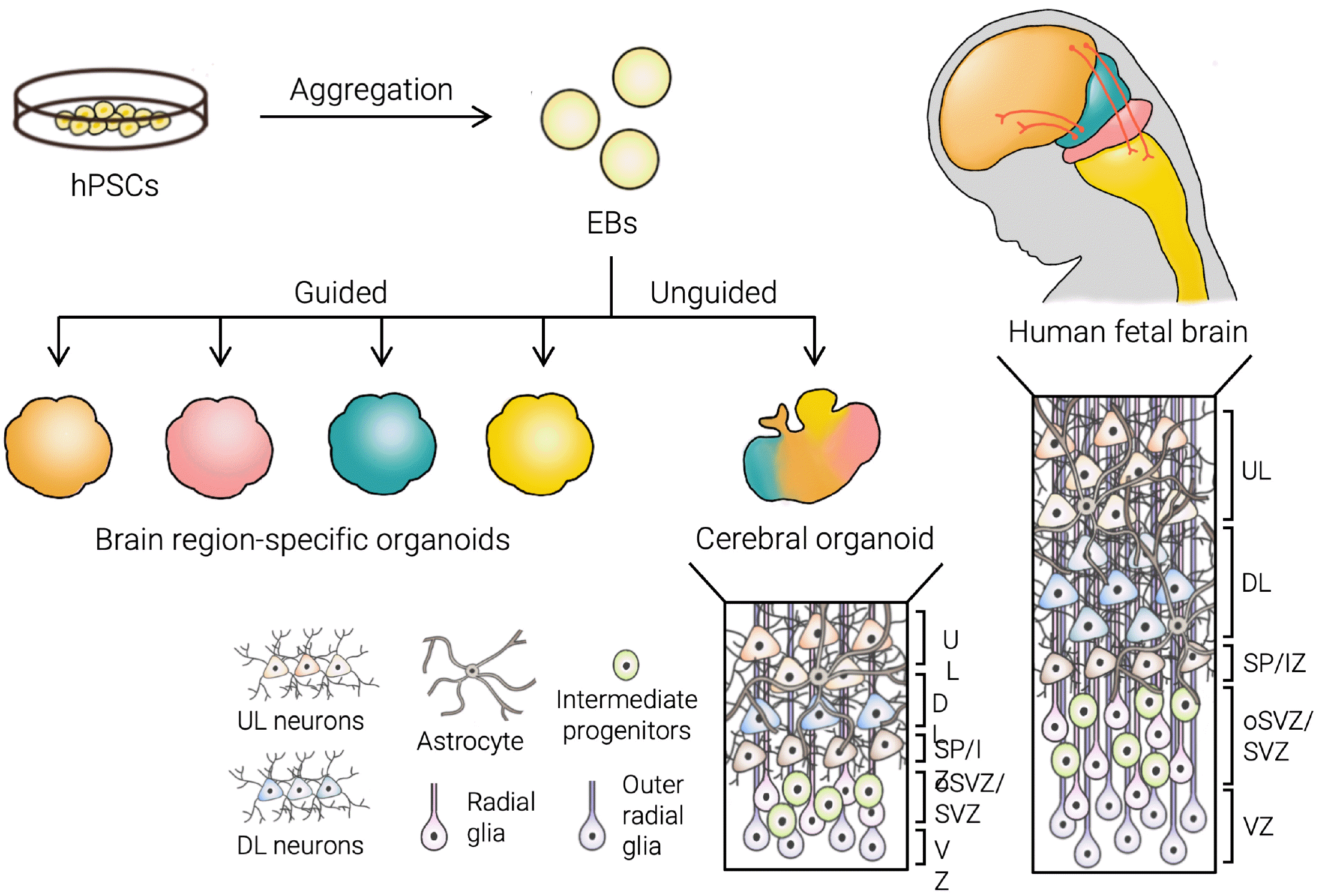 | Fig. 2Generation of brain organoids. The process of brain organoid generation consists first of aggregation of hPSCs into EBs and then further differentiation to organoids. EBs can undergo guided differentiation with small molecules into region-specific organoids or unguided differentiation to generate cerebral organoids. Though cerebral brain organoids successfully replicate many aspects of the human fetal brain, they are still limited with regard to many factors such as the depth and complexity of the cortical lamination.
|
Altogether, these brain models successfully replicate the aspects of the developing brain, including cellular diversity, micro-architectural features, and in vivo developmental trajectories. Until now, brain organoids undoubtedly have advanced our understanding of human brain development and provided unprecedented opportunities to study brain disorders in human-specific experimental settings (10).
Although brain organoids stand as the state-of-the-art model for the human brain, there are still challenges to be addressed. One of the major hurdles for brain organoid technology is the ‘batch effect,’ which is the heterogeneity across the different batches of samples or different culture methods. Second, the conventional protocol to generate brain organoids mainly involves the uniform bath application of signaling molecules in the culture, which fails to recapitulate spatiotemporal signaling dynamics that regionalize the brain for its complex structure and function. Third, although transcriptome analysis between brain organoids and fetal brains revealed remarkably similar developmental trajectories and even showed early postnatal signatures (11), the maturation status of the current brain organoids is yet far away from advanced neural functions in the adult brain. In addition, progressive cellular stress during long-term in vitro culture may impair the cell fate transition and normal cellular functions (12). Therefore, an improved long-term culture method to enhance the maturation of brain organoids is necessary to model the later stages of development. Lastly, the current organoid culture methods provide an incomplete microenvironment, lacking an extracellular matrix and non-neural cell types, which significantly contribute to the critical steps of brain development in vivo.
In this review, we will introduce the up-to-date engineering methods to improve brain organoid technology. By applying the various engineering strategies, we may pave the way to fully reproduce the in vivo contextual environment and eventually generate brain organoids harboring functionally mature neural circuitry with an intact cytoarchitecture and inter-regional connection.
Go to : 
Challenges and Potential Breakthroughs to Overcome the Current Limitations of Brain Organoid Technology
Towards consistent and homogenous brain organoid generation
One of the major limitations of brain organoids is the batch effect. Such inconsistent generation of brain organoids may yield substantial phenotype variance, hindering our understanding of the accurate brain development process or disease underlying mechanisms. This is largely due to the lack of an in vivo like environment in which various factors, such as signaling molecules, regulation of gene expression, and extracellular interaction, work together to control brain development precisely. Indeed, the brain organoid generated through unguided differentiation, which largely depends on the self-patterning and self-organization of PSCs, yields marked variation in structure, cellular diversity, and transcriptomes (6, 13, 14). To reduce the heterogeneity in brain organoid formation, several culture methods and bioengineering approaches have been introduced.
Engineering size and geometry of the embryonic body: The formation of EBs, which have the potential to differentiate into all three germ layers, is mediated by the self-assembly of PSCs and is the first step to generate brain organoids. Previous studies have shown that the size of EBs, thus the initial number of cells to aggregate, matters when it comes to the ectoderm-specific differentiation. In fact, a decreased size of EBs yielded a relatively effective differentiation into the neuroectoderm (15), which may be largely due to a lower surface-area-to-volume ratio. Nowadays, many protocols involve the use of cell-non-adhesive U-/or V-bottom plates to assemble EBs in controlled and uniform manners (Fig. 3A). An alternative strategy may be a physical dissociation of brain organoids to reform them into smaller pieces using the narrow opening of a Pasteur pipette, resulting in a smaller size and consistent formation of a single rosette structure (16). Furthermore, it was shown that not only the size but also the geometry of EBs matters when it comes to brain organoid development (15). The elongated EBs assembled with poly(lactide-co-glycolide) copolymer (PLGA) fiber microfilaments as a scaffold showed an enhanced neuroectoderm lineage specification. Several bioengineering approaches have been introduced to modulate the size and the geometry of EBs, including microwells (17-19) and micropatterned extracellular matrix (20, 21), showing potential to be applied to brain organoid to better guide into the neural fate (Fig. 3A).
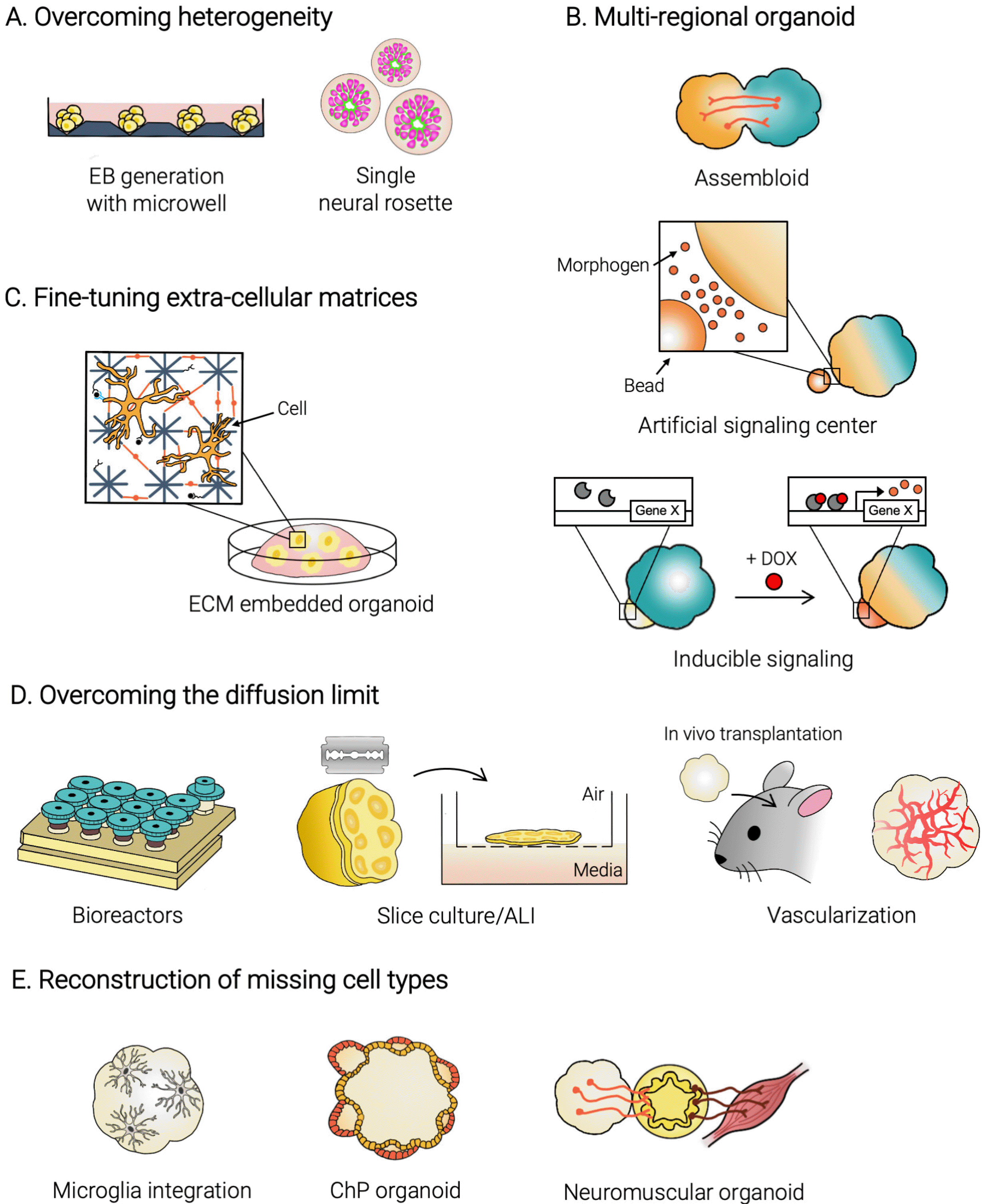 | Fig. 3Engineering advances to overcome major hurdles of brain organoid technology. (A) Efforts to overcome the “batch effect” include using standardized microwells to generate homogenous EBs and generating organoids with single neural rosettes. (B) The tradeoff between the multi-regional complexity but low reproducibility of unguided cerebral organoids with high fidelity but low complexity of region-specific organoids may be overcome by generating mutli-regional organoids. Multi-regional organoids can be generated through assembloid production, morphogen gradients created by artificial signaling centers, and chemical and/or light inducible systems. (C) Mimicking the neural ECM in brain organoid culture systems is a challenge that has been addressed by modulating the properties of both naturally-derived and synthetic biomaterials. (D) Various approaches may be used to overcome the “diffusion limit” to enhance the long-term culture and mature of brain organoids. These include the use of bioreactors, organoid slice culture at the air-liquid interface, and vascularization through in vivo transplantation. (E) Organoids lack many important connections among various cellular subtypes within the human brain. These missing cell types may be accounted for through microglia integration (neuro-immune), CFS producing ChP organoid generation (meninges-brain), and neuromuscular organoid generation (PNS-CNS).
|
Guiding the differentiation of brain organoids: A series of protocols that make use of signaling factors to instruct the organoid differentiation into distinct brain regions have been developed, the so-called ‘guided differentiation.’ In these protocols, small molecules or growth factors, which were previously discovered in model organisms to direct the brain regionalization in vivo (22), are added to the culture media. Such patterning with certain factors delineates the brain organoid development while improving the stochastic and spontaneous differentiation of EBs. The dual SMAD inhibition, which blocks the SMAD pathway, is widely used for neuroectoderm lineage specification, and its prolonged exposure led organoids to have dorsal forebrain characteristics with fidelity (7, 23). Moreover, the additional treatment of SHH, a ventralizing factor determining the ventral to dorsal axis of the brain, produced the ventral forebrain organoid as well (24, 25). Until now, numerous region-specific brain organoid protocols have been introduced, resembling the midbrain (7, 26), striatum (27), hippocampus (8), choroid plexus (28), thalamus (29), hypothalamus (7), hindbrain (30), cerebellum (9), and spinal cord (31).
Guided differentiation demonstrates an increased reproducibility of generating organoids with respect to structures and cell types (32) and further diminishes the batch variation even across different cell lines (33). However, the amount of factors present in the media must be carefully determined because the intrinsic threshold may be programmed differently in individual cell lines (34, 35). The use of external factors is still artificial, and it may often lead to a distorted developmental scenery, potentially misleading our understanding of the intrinsic properties of brain development.
A brain organoid with multi-regional identities
There is a trade-off between the unguided and guided differentiation of brain organoids; the former leads to the emergence of various regions interacting together for development and functionality but suffers from low reproducibility, while the latter produces a region-specific brain organoid with high fidelity but does not resemble the multi-regional brain structure in vivo. The brain is composed of interconnected circuits that span through the various brain regions, and many brain disorders involve malformation or malfunction of those circuits. Therefore, the investigation of the interaction and connectivity among various brain areas is important. In fact, a recent article reported the generation of brain organoids with primordial eye fields, consisting of conserved developmental properties for both tissues and light-responsive functional connectivity (35). Similarly, it is increasingly required to develop advanced culture techniques that could resemble multiple regional identities in a single brain organoid (Fig. 3B).
Assembloid - assembly of organoids with different regional identities: One of these techniques is called the ‘assembloid,’ which means the fusion of organoids representing different brain regions. The assembloid has shown successful assembly of organoids to mimic the inter-regional phenotypes, which could be used in studies for cell migration pattern or functional neural circuitry between particular brain regions (24, 25, 29, 36). In the case of the cortico-striatal, the migration of interneurons from the dorsal to ventral forebrain has been observed with the integrated electrophysiological connectivity, similar to the case of in vivo development (24, 25, 27). Furthermore, other various regional interconnections, such as thalamo-cortical (29). and hypothalamic-pituitary (37), have been successfully modeled via assembloid technology as well. More interestingly, a three-compartment assembly, the so-called cortico-spinal-muscle assembloids (38), was made which showed the functional connection to modulate muscle activity. The significance of the assembloid is that it not only resembles the in vivo-like circuitry but also enhances the neuronal maturation compared to single-region organoids, suggesting that the co-existence of distinct brain regions may be a key to further accelerating the developmental aspects of brain organoids.
Incorporation of an engineered signaling center for regionalization: During brain development, the signaling center has a substantial role in the regionalization of the brain, which mediates the dynamic gradients of morphogens to modulate its subsequent activation of gene expression essential for acquiring distinct positional identities (39). Such signaling centers have been observed along the boundaries of areas with distinct regional signatures in the unguided brain organoid (5), implying that the reproduction of signaling dynamics is critical for determining regional identity within the brain organoid. Indeed, the brain organoid culture that involves establishing morphogen gradients in a developmental context-relevant manner has received increasing attention in recent years. An engineered signaling center incorporated into brain organoids may create the morphogen gradients rather than using signaling molecules that are equally bathed in the culture media. As a result, the brain organoids are able to be spatially structured in response to the signaling gradient, displaying positional information or several distinct brain regions within a single brain organoid.
The simplest technique to introduce an artificial signaling source is to use morphogen-soaked beads, which is a common strategy for manipulating signaling in classical developmental studies (40). The degree of signaling may differ based on the concentration of the signaling molecules absorbed in the beads or the distance from the beads, through which a signaling gradient is established with diffused morphogens. Indeed, a recent study utilized a WNT agonist- and BMP4-soaked beads to successfully direct dorsoventral and anteroposterior topography in brain organoids (41). The spatial topography was determined by the concentration and position of the beads, in which stronger exposure to WNT and BMP yields more anterior/rostral molecular signatures. Such straightforward technology to mimic the signaling center in human brain development showed potential to generate brain organoids with controllable regional specification.
A cell-based signaling center is another strategy, which involves a group of cells that releases signaling factors. In this method, the cells are modified to express the target signal molecules and inserted at one pole of the EBs to further specify the following differentiation. For instance, BMP4 treated aggregates were combined with the untreated larger aggregates while the former acts as the signaling center (42). The signaling induced the expression of the downstream pathway, Wnt and Nodal, at one side and resulted in symmetry breaking of the EB, generating embryonic-like structures with the specified axis. Another example utilized Chd3- and Wnt3A-expressing HEK cells, which enable the spontaneous formation of a cell cluster and induction of the Wnt/β-catenin pathway, respectively (43). The cell cluster was attached to the EB and guided the patterned mesoderm-specification at the attached site. Both studies showed that a cell-based signaling center is a promising technique to recapitulate the biochemical environment throughout development.
Inducible systems for manipulating signaling with high spatiotemporal resolution: The signaling dynamics during brain development are precisely confined in space and time. To mimic such signaling activities in a spatiotemporally-restricted manner, an inducible gene expression system is now utilized on top of a cell-based signaling center. A chemical-inducible system, in which the addition of chemicals such as doxycycline can regulate the expression of the target signaling molecules, has been shown to direct positional identities in brain organoids (44). For this, a cluster of cells with a chemically-indu-cible system for SHH was embedded at one side of the forebrain organoid; its expression was controlled with doxycycline to generate a gradient of SHH. Such asymmetric SHH signals led to the assembly of the dorsoventral forebrain axis in the brain organoid.
Besides chemicals, researchers also attempted to use a light-inducible system instead, which may provide a greater degree of freedom in stimulation with respect to space and time. In a study, the light activable Wnt system was used to drive mesoderm-specific differentiation of hPSCs along with the induction of subpopulation-wise self-organization of cells in a 3D culture (45). Similarly, optical control of curvature in neuroectodermal organoids was reported, indicating the possibility to modulate brain organoid morphogenesis using a light-activable system (46). A more recent study has utilized a light-inducible Cre/Lox recombination system to activate the expression of SHH only in the photo-stimulated area within the neural organoid, resulting in the establishment of the dorsoventral positional identity (47). Possibly, a light-inducible system with high flexibility and reversibility, such as a light-activable CRISPR/Cas9 system (48), may provide a more versatile platform for brain organoid manipulation.
Recapitulating the neural ECM in brain organoid culture
The neural extracellular matrix (ECM) has essential roles throughout the stages of the developing brain. Composed primarily of hyaluronic acid (HA), reelin, chondroitin sulfate proteoglycans, link proteins, tenascins, and laminins and with low levels of fibrous proteins, the neural ECM serves as a matrix of biochemical and biophysical cues that regulate the proliferation, differentiation, and migration of cells within the brain (51-53). Therefore, the selection of an appropriate ECM is critical for brain organoid systems because it provides the mechanical and signal-rich context for organoids in vitro culture systems (Fig. 3C).
Naturally derived biomaterials such as Matrigel and laminin are the most widely-used ECM for currently established protocols of cerebral organoid generation (5, 7, 54, 55). In particular, Matrigel is a murine sarcoma-derived reconstituted basement membrane that consists of a complex cocktail of about 2,000 unique proteins (56). However, while it is commercially available and easy to use, it is undefined and inconsistent among different batches and lacks many neural ECM components such as glycoproteins (56-58). Furthermore, studies report that Matrigel may bias neural differentiation by affecting neuronal yield and neurite length compared to other ECM materials (59, 60). Therefore, other natural and synthetically engineered biomaterials have been developed that may better recapitulate the in vivo environment of the developing human brain and provide a defined physical scaffold to guide growth and differentiation.
As alternatives to Matrigel, other naturally-derived biomaterials and cocktails may provide an improved culture environment for brain organoids. In particular, decellularized ECM (dECM) extracted from brain tissue more robustly mimics the neural ECM (61). For example, culturing human neural stem cells in a mixture of decellularized porcine brain ECM and laminin-coated silk scaffolds supplemented with type I collagen not only produces functionally mature neurons and astrocytes but also reduces reactive astrogliogenesis (61). The use of HA or modified HA may also promote differentiation of human pluripotent cells towards neural fates (62-64). Notably, HA-based hydrogels (HA and chitosan in a dextran-sodium chloride solution) can be used to differentiate induced pluripotent stem cells (iPSCs) into NPCs in the absence of any neural induction components (64). Density gradients can also be created using 3D layered hydrogels of methacrylate-modi-fied HA to closely observe neuronal migration and accelerate neuronal maturation (62). Using this approach, a study differentiated human iPSC-derived NPCs within a density gradient and reported migration defects caused by mutations associated with a neurodevelopmental disorder (62). These efforts to incorporate natural biomaterials to guide neuronal culture may be applied to 3D cultures to enhance brain organoid polarization and cortical layer formation.
As opposed to natural biomaterials, the use of synthetic biomaterials as the ECM for brain organoid culture enables for customization and control. The fine-tunable properties of extracellular matrices include ligand concentration, stiffness, degradability, and composition of the neural ECM components such as laminin and fibronectin. For example, modulating concentrations of adhesive ligands such as RGF, YIGSR, and IKVAV in matrices can increase cell survival and improve neuronal morphology and marker expression (65). Furthermore, PEG hydrogels can be modified to optimal stiffness and combined with other ECM components to promote neural differentiation and control organoid size and polarity (66, 67). Efforts through screening experiments to optimize the various parameters of synthetic neural extracellular matrices have demonstrated the potential to customize and “design” matrices best suited for the particular purpose of the brain organoid culture.
In summary, while natural biomaterials may provide improved organoid growth and architecture, they may also have high variability within batches. In contrast, synthetic biomaterials such as PEG provide a reproducible, well-defined microenvironment with the ability to fine-tune various parameters for optimal brain organoid culture. However, they lack the complex network of biochemical cues essential for cell physiology present in the neural ECM and partially in naturally derived biomaterials. To achieve a highly reproducible and optimal growth environment for culturing brain organoids that accurately model neurogenesis, future work should focus on characterizing the individual and interactive contributions of the ECM components and the impacts of the cell-secreted ECM in the designed matrix.
Overcoming the diffusion limit in brain organoids
Long-term maintenance of organoids that sufficiently mimic the physiological functions of the postnatal and adult brain is one of the major challenges of organoid technology. Previous studies have shown that the majority of brain organoids can only reach a transcriptomic “age limit” of a second-trimester fetal brain (12, 33, 67). A recent study reported that brain organoids grown for 250 to 300 days could exhibit developmental signatures similar to that of the postnatal brain (11). Regardless, the in vitro culture environment poses many issues regarding organoid maturation and function, such as increased cellular stress and impaired cell-type specification (11, 12). In particular, a lack of even diffusion of oxygen and nutrients is one of the main limiting factors of brain organoid maturation in long-term culture. This eventually causes a necrotic core to build up inside the organoids, thus limiting further size increases. Different approaches have been used to address these issues with diffusion and to enhance the uptake of oxygen and nutrients (Fig. 3D).
A simple technological improvement that can vastly improve nutrient and oxygen absorption is to use spinning bioreactors. 3D printing technology has enabled the design and production of more accessible and cost-efficient spinning bioreactors for use in brain organoid cultures (5, 7, 68). Utilizing miniaturized spinning bioreactors such as SpinΩ produces brain organoids with reduced heterogeneity and may more robustly recapitulate primate-specific features of early neurodevelopment such as the presence of the outer subventricular zone and outer radial glial-like cells (7, 68). Additionally, rotating wall bioreactors may also be used to improve organoid culture, as can be seen in previous studies with retinal organoids (69-71). Incorporating spinning bioreactors in organoid culture can accelerate and improve growth and differentiation by enabling the efficient uptake of oxygen and nutrients.
Slice organoid culture at the air-liquid interface may also substantially enhance the long-term culture and maturation of brain organoids. Most brain organoid models are sphere-like structures, and therefore the diffusion limit is unavoidable without integrating an active circulation system. In contrast, a slice organoid culture at the air-liquid interface involves sectioning the organoid into disks to improve surface diffusion into the interior of the organoid. This method of slice culture at the air-liquid interface reduces cell death within the organoids and enables the expansion of the cortical architecture (72-74). Improved maturation and long-term culture through slice cultures may be particularly useful in modeling late human neurodevelopment, a stage that is critically important to study adult-onset diseases such as ALS/FTD (73).
An alternative to the previous methods of overcoming the diffusion limit is to introduce a perfusion system to the brain organoids through engineering or biological techniques. 3D printing is the basis behind the majority of the engineering methods, whether it be through the printing of biomaterial-derived casts for creating vascular networks (75), or direct printing of vasculature and/or vascular scaffolds (76). It is also possible to directly introduce a perfusion system to brain organoids through biological techniques. In vitro approaches may consist of coculturing brain organoids with endothelial cells (77, 78), using genetic engineering to induce vasculature-like network formation during brain organoid growth (79), or incorporating of the external perfusable device (80-82). One study ectopically expressed human ETS variant 2 (ETV2) in hESCs to induce the formation of vascularized networks during a brain organoid culture (79). Another study combined a human brain tissue-derived dECM and microfluidic systems to recapitulate human brain microenvironment and to improve the nutrient and oxygen supply (80). In vivo approaches consist of grafting brain organoids into a host animal brain, making the host act as a bioreactor to provide a perfusion system (83). Brain organoids transplanted into a host such as a mouse brain not only develop functional vasculature and graft-to-host synaptic connections, but they also show increased cell survival, alleviation of stress pathways, and enhanced cell type specification compared to in vitro culture (12, 83). Regardless of which approach is used, future models of brain organoids may require the incorporation of vascular networks to better emulate the context of the human brain.
Reconstruction of missing cell types
Besides the perfusion of oxygen and nutrients, another critical component in the successful maturation of brain organoids is the development of connections among the various cellular subtypes. Current protocols fail to fully capture the complex interactions within the human nervous system, such as the interaction between neuronal and non-neuronal cell subtypes. Various studies have addressed these issues by incorporating missing cell types through small molecule induction or coculture (Fig. 3E).
One important example is the incorporation of microglia, the main source of immune defense in the central nervous system (CNS) and also a regulator of neurodevelopment (84, 85). Because microglia are derived from the mesoderm lineage, most brain organoid protocols that direct differentiation towards neuroectodermal lineages prevent the formation of microglia (86). However, microglia develop innately in brain organoids that are grown in neural organoid media without dual-SMAD inhibitors to enable for mesoderm differentiation (86). Microglia can also be incorporated through coculture, as demonstrated in a recent study that developed a rigorous model enabling the control of the ratio of human microglia in brain organoid culture (87). Incorporating microglia in brain organoid cultures is crucial to produce fully mature brain organoid models to study human-specific aspects of microglia during neurodevelopment.
In vitro organoids also lack the distinct boundaries provided by the membranous layers surrounding the CNS in the human brain. Consisting of the dura, arachnoid, and pia mater, these meningeal layers provide a physical boundary that protects and nourishes brain tissue and acts as a signal-rich niche required for brain cell maturation (88). For example, dorsal forebrain meninges control the timing of neurogenesis by the secretion of retinoic acid (88). Dorsal forebrain meninges interact with radial glial endfeet to influence the cell cycle exit of neuron progenitors, neuron production, and neuroepithelium elongation (88). The physical boundary that the meninges present may also help create and maintain concentration gradients of factors secreted by cortical layer neurons. Reelin, an extracellular matrix glycoprotein secreted by cells of the marginal zone in the developing forebrain, is known to affect neuronal migration and layer formation(89-91). In contrast, the lack of a closed boundary in typical organoid culture conditions, where they are suspended in liquid media, may impede the formation of these concentration gradients. This explains how brain organoids exhibit less complex and spatially distinct cortical layers compared to in vivo brain tissue. Looking forward, a possible approach to mimic the meninges will be to develop biomaterials encasing organoids that provide an appropriate ECM context and closed boundary conditions. Alternatively, another approach will be to coculture meningeal cells derived from neural crest lineages with neural cells to incorporate meninges into brain organoid culture.
Choroid plexus (ChP) and cerebrospinal fluid (CSF) production is another aspect of the meninges-brain interacting environment that needs to be considered in brain organoid cultures. The choroid plexus is found within the pia mater, the innermost layer, where it secretes the nutrient and signaling molecule-rich CSF. It also forms the blood-CSF barrier (B-CSF-B), which, together with the blood-brain barrier (BBB), works as a protective barrier blocking the entry of toxic substances to the CNS. These two main functions of CSF production and blood-CSF barrier formation are vital for brain function, and recent studies have established protocols to model the ChP in vitro through ChP organoids (28, 92). ChP organoids develop the ChP epithelium, express known biomarkers, and produce a CSF-like fluid (28, 92). Importantly, one study showed that their ChP organoids can quantitatively predict the CNS permeability of known molecules (28, 92), demonstrating their potential to be used for drug screening. Furthermore, another study conducted transcriptome analysis on organoids generated by their ChP organoid protocol and found that their ChP organoids showed high transcriptome correlation levels to adult human choroid plexus tissue (28, 92). A notable finding from studies using the ChP organoid model is that SARS-CoV-2 productively infects the ChP compared to other brain regions in organoid models (28, 92, 93). The establishment of 3D ChP models is an exciting development that will provide effective platforms for drug testing and exploration of human-specific disease mechanisms related to CNS protective barriers.
Lastly, brain organoid models represent only a portion of the human central nervous system. However, the nervous system consists of both the peripheral and central nervous systems, and models that accurately represent the complex CNS-PNS interactions have yet to be developed. An exception is the development of neuromuscular organoids (NMO), which generate functional neuromuscular junctions and can contract (94). Further efforts are required to generate rigorous models of the interplay between the CNS and PNS, including the connections along the corticospinal tract. Accurate representation of all parts of the nervous system in brain organoid culture will enable advanced maturation of brain organoids in vitro and provide powerful tools to study the interactions between neuronal and non-neuronal regions during neurogenesis.
Go to :
Advanced Analysis Tools for Brain Organoids
Although brain organoids provide new opportunities for modeling human brain development and brain disorders, the complex cell-cell interaction among diverse cell types in the 3D environment makes it difficult to precisely analyze the functions of cells residing in brain organoids. Recently developed tools such as CRISPR-based barcoding and multielectrode recording devices are promising breakthroughs to overcome these limitations (Fig. 4). This section will focus on recent advances in new technologies to analyze the complex nature of brain organoid systems.
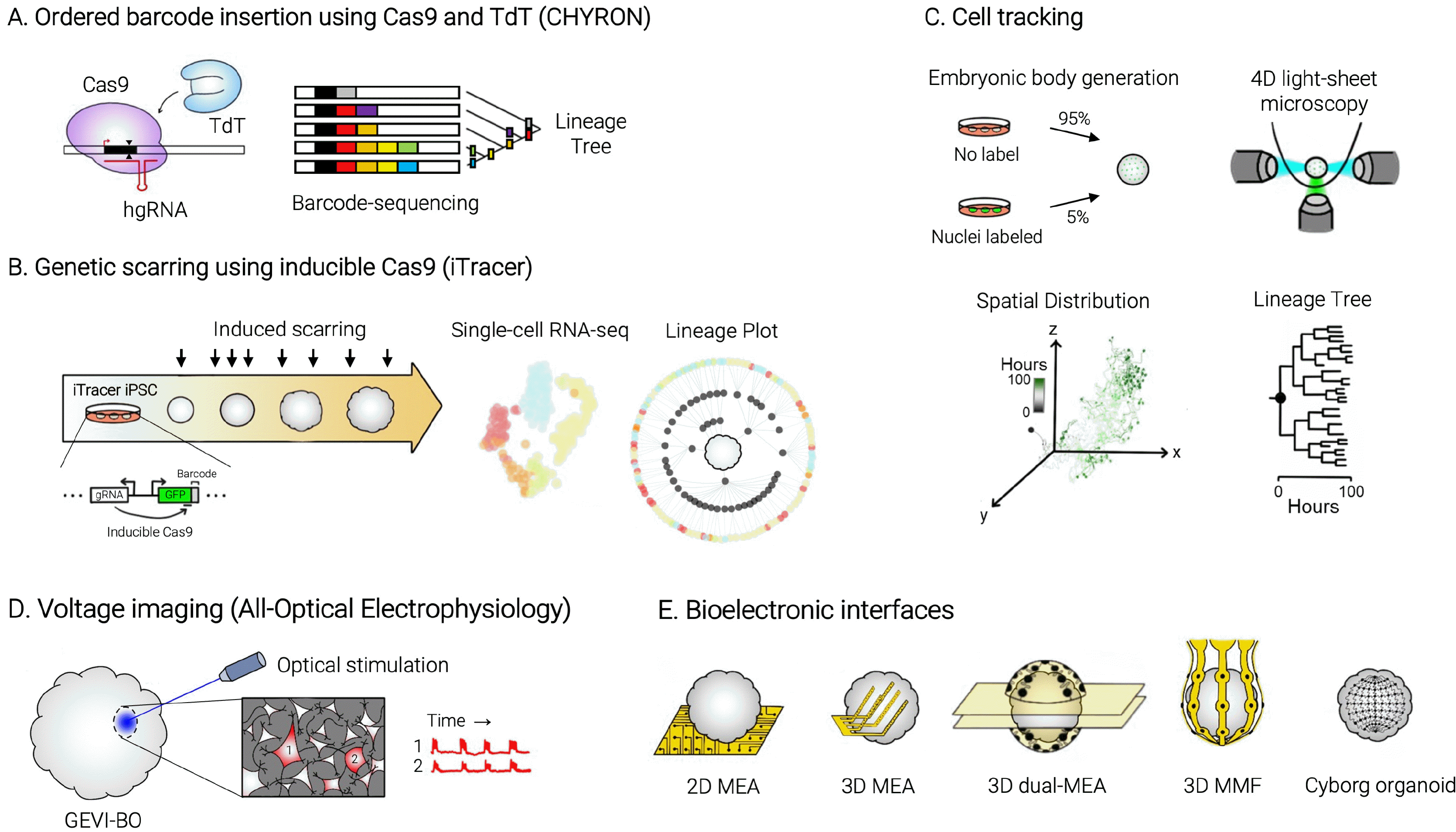 | Fig. 4Advanced tools to analyze the functional properties of the brain organoid system. (A∼C) Lineage tracing tools to elucidate spatiotemporal dynamics of developing organoids, including dynamic lineage tracers and cell tracking. (D, E) Electrophysiological methods to monitor electrical activity in brain organoids, including voltage imaging and bioelectronic interfaces.
|
Monitoring the trajectory of cell-type specification using lineage tracing tools
In the developing brain, neural stem cells produce diverse types of cells depending on the spatiotemporal context. Lineage is a developmental path connecting the founder cell to its descendant cells. If a single progenitor cell is genetically marked and the mark is transmitted to all the progenies that originated from the founder cell, lineage information about the number of progenies, their location, and their differentiation status could be traced. Construction of lineage trees throughout brain development is beneficial and informative in modeling differentiation processes in vitro, opening the possibility of manipulating cell fates in vivo or identifying the origin of developmental pathologies or cancer (95).
Imaging-based pre-genomic methods such as a tracer dye, cell transplantation, and in toto confocal microscopy have been used to visualize cell lineages (96, 97). However, the temporal and molecular resolution of imaging-based techniques is not sufficient to fully capture the details of complex lineage relationships. Alternatively, lineage information can be extracted from genetic changes such as naturally-occurring somatic mutations or artificially induced genetic mutations (98-101). In this genetic lineage tracing, lineages of individual cells are permanently recorded in the genome. Current strategies that induce and analyze genetic manipulation at a specific locus are classified as dynamic lineage tracing, which can be divided into three approaches: transposon integration, Cre-loxP recombination, and CRISPR-Cas9 editing (102). Transposon-based recording usually inserts a fluorescent reporter cassette whose 3’ UTR carries a short sequence working as a barcode (103-106). Cre-loxP recombination sites or CRISPR-Cas9 target sites can also function as a barcode, usually introduced by homologous recombination or lentiviral transduction (107). After barcode integration and recording, lineage barcodes must be recovered by DNA or mRNA sequencing to construct a lineage tree. When edited sequences are predictable, in situ imaging such as seqFISH (sequential single-molecule fluorescence in situ hybridization) can be used as a visual readout, rather reaching a much higher recovery (108).
Dynamic lineage tracing tools have been actively applied to zebrafish and mouse embryos to monitor cell fate specification during embryogenesis. For instance, CRISPR-based GESTALT (Genome Editing of Synthetic Target Arrays for Lineage Tracing) revealed that few embryonic progenitor cells give rise to each organ of the entire zebrafish by implementing a barcode array consisting of 9 to 12 Cas9 targets (109). Cre-based barcoding called Polylox implemented an artificial recombination locus with ten loxP sites in alternating orientations and demonstrated the multilineage priming of mouse hematopoietic stem cells (HSCs) (107). Such sophistication of the barcode composition was followed by combination with single-cell transcriptomics, for example scGESTALT, becoming capable of incorporating cell state information into differentiation trajectories (110). Single-cell lineage tracing has been assisted by vigorous development of barcode systems such as ScarTrace (111), LINNAEUS (112), MARC1 (113), CellTagging (106), LARRY (105), etc. (For more detail of these technologies, refer to this review (114).
Recently, CHYRON (Cell History Recording by Ordered Insertion) emerged as a cutting-edge lineage recorder based on the CRISPR-Cas9 system (Fig. 4A) (115). Unlike existing methods relying on indel mutations of the barcode, Loveless et al. (2021) minimized deletion and facilitated insertion mutations at the barcode by combining Cas9, a homing guide RNA (hgRNA), and the template-independent DNA polymerase terminal deoxynucleotidyl transferase (TdT). Cas9 targets a specific sequence in the genome under the guard of guide RNA and generates a double-strand break. TdT can insert random nucleotides without the template DNA in repairing this double-strand break. Homing guide RNA, a self-targeting guide RNA, keeps taking Cas9 to the hgRNA-encoding locus so that insertion occurs in the same locus repeatedly. Thus, ordered insertions could be successfully recorded in the barcode and then safely reconstructed as cell population lineages through barcode-sequencing. By engineering Cas9 and TdT to be expressed only under the stimuli-of-interest (e.g., hypoxia), CHYRON could also capture the extent and timing of exposure to the biological stimuli. This technology will provide rich information about brain organoids, investigating lineage relationships between various cell types or cell populations that experienced particular stimuli to what extent.
Yet, the only lineage tracing tool applied to the brain organoid study has been iTracer (Fig. 4B) (116). Previous approaches were based on computational analyses, using the Monocle package, to construct pseudo-time trajectories from single-cell RNA-seq data in brain organoids (33, 117-120). iTracer is based on the sleeping beauty transposon system, integrating the 3’-barcoded GFP and the barcode-targeting guide RNA into multiple genomic loci. Cerebral organoids were generated with iTracer iPSCs containing an inducible Cas9 cassette. Cas9 expression is induced by introducing doxycycline at different developmental time points, resulting in genetic scars at the barcode. Editing may mirror versatile biological responses by modulating scar-inducing signals. Cells expressing lineage barcodes were subjected to lineage-coupled single-cell and spatial transcriptomics, which were both the first use in the brain organoid field. Transcriptome analyses revealed differentiation and regionalization trajectories of the iPSC clones by connecting different time points and distinct regions. Still, there are some technological limitations left to be solved, including transgene silencing, single-cell dropout, and sampling depth, for the sake of comprehensive lineage reconstruction from the iTracer system.
He et al. (2020) emphasized how powerful it will be to use the iTracer system together with 4D light-sheet microscopy, which is one of the breakthrough technologies that can secure temporal resolution of imaging-based lineage tracing. They demonstrated long-term live imaging of developing cerebral organoids by tracking individual fluorescently-tagged nuclei and observed local amplification of the initial clones. This concept is in line with ‘cell tracking,’ which follows each developing cell in real-time and draws complete lineage trees to monitor spatiotemporal developmental dynamics (Fig. 4C) (121).
By identifying single stem cell lineages during cell fate specification, we can decipher complicated developmental processes such as discovering unique and transient cell types. Brain organoids came about as a potent in vitro model that can provide direct evidence in the investigation of the differentiation dynamics in human brain development. Lineage-coupled single-cell and spatial transcriptomics along with long-term time-lapse live imaging of barcoded organoids provide an unprecedented opportunity to overcome doubts in transcriptomic and spatial resemblance to the real human brain. As in human colon and retinal organoids, analyzing lineage relationships of diverse cell populations will contribute to the understanding of developmental effects of mutations and the modeling of cancer progression (122-125).
Systemic analysis of neural activity in brain organoids
As we discussed, brain organoids have the potential to fill the gap between human monolayer cell culture models and animal models such as rodents. Although effective and accurate electrophysiological approaches have been developed for two-dimensional cultures, functional analysis of electrical activities of brain organoids is limited (126). Classical patch-clamp electrophysiology enables the recording of a single cell with a high temporal resolution but has difficulties grasping the complex activity of a group of cells due to its invasive and laborious nature. Thus, recent advances in the past several years to overcome limitations posed by the three-dimensional structure of brain organoids will be discussed here.
The classical methods for measuring electrophysiological activities, such as patch-clamp, offer high temporal resolution yet suffer from low spatial resolution to record whole-organoid activity (23). The imaging-based methods, such as calcium imaging, may provide larger-scale recording but lack of high temporal resolution (5). Lastly, microelectrode arrays (MEAs) (72) can provide both network-scale and relatively high temporal resolution but currently do not support the recording in three-dimension, which is in increasing need to accurately analyze electrophysiology in brain organoid.
Calcium imaging has been used to monitor neuronal activities at the single-cell level and their synchronization but suffered from its limited temporal resolution (25, 130). The problematic penetration depth is expected to be improved by the development of two-photon and light-sheet microscopy, expanding the region-of-interest from the organoid surfaces to deeper layers (127-129). In comparison, whole-cell patch-clamp recording can measure the detailed properties of one neuron at a time in millisecond temporal resolution while lacking network connectivity (23). Thus, these two techniques can be used together comple-mentarily. For example, Xiang et al. (2017) confirmed the neuronal activity and connectivity in fused MGE (medial ganglionic eminence)-cortical organoids (25).
Similar to calcium imaging that uses genetically encoded calcium indicators (GECIs), voltage imaging that uses genetically encoded voltage indicators (GEVIs) was developed as another option. Originally, voltage-sensitive dyes have been used because of their good speed and sensitivity but it’s hard to restrict them to specific cell populations (131, 132). Hochbaum et al. (2014) solved this problem by applying spectrally orthogonal GEVI and an optogenetic actuator, which is called all-optical electrophysiology that can visualize membrane voltage changes upon optical stimulation (Fig. 4D) (133). Recently, a synthetic NIR voltage reporter was invented by Puppo et al. (2021) for easy and safe delivery without triggering any bias (134). However, the technologies mentioned so far are incompatible with long-term stable measurements. The accuracy and resolution of calcium and voltage imaging are disrupted by the decaying dynamics of the indicators and increasing noise as the neuronal firing rate increases. Patch-clamp recording lacks long-term reliability due to difficulties in the maintenance of the subject, which is also confined to organoid edges or sliced cultures (101). This is where bioelectronics can have a big part, enabling noninvasive access to electrophysiological information whenever we want (Fig. 4E).
One of the most frequently used devices in electrophysiological measurements of brain organoids is a microelectrode array (MEA), which is a neural interface in which interconnected microelectrodes record electric signals from neighboring cells. 2D MEAs have been widely applied to the analysis of brain organoids, due to their good temporal and network resolution and larger data scale. For instance, Giandomenico et al. (2019) confirmed the functionality of long-range neural circuits and the connection between several regions within the organoid (72). Trujillo et al. (2019) conducted a comparison study that examined the electrical activities of developing forebrain organoids by weekly MEA recordings for 10 months and EEG features of preterm infants, suggesting that the evolution of neural networks in the developing organoids resembles that of preterm infants to some extent (135). In combination with single-cell RNA-seq and immunohisto-chemistry data, Fair et al. (2020) elucidated the electrophysiological maturation of forebrain organoids correlated with transcriptomic and morphological features (136).
Despite the progress that 2D MEAs have made, their planar structure cannot record the inner part of 3D brain organoids. Therefore, 3D MEAs were developed by folding the electrode array into a hinged form so that the vertically aligned part of the probes can monitor the inner part as well. Soscia et al. (2020) developed flexible MEA probes using polyimide layers that can be naturally positioned to the inner part of the organoid by seeding the cells (e.g., iPSC-derived neurons) embedded in conditioned ECM on the probes and then growing the cells to establish a 3D structure around the probes (137). There are a lot of improvements being made, such as implementing high-density MEAs that enable single-cell analysis and 3D multifunctional MEAs that can both monitor and modulate neural activities in real time (138-142). However, MEAs still have some problems including their limited contact with the organoids and the inconvenience arising from organoid culture protocols that require frequent media exchange. Shim et al. (2020) recently fabricated a 3D dual-MEA to overcome these problems by sandwiching brain organoids between two stretchable polyimide layers, in which high stretchability was engineered with serpentine interconnects (143).
Another approach in bioelectronics has been proposed to outperform MEA technologies. By enhancing the flexibility of polyimide arrays to softly envelop the entire brain organoids, Park et al. (2021) designed a 3D flexible neural interface precisely matching the shape of cortical spheroids; therefore, gold microelectrodes are positioned proximal to the surface of the organoids (144). They named this 3D multifunctional mesoscale framework (3D MMF), where additional functional elements such as an optical device, thermal actuator, or electrochemical oxygen sensor enable simultaneous stimulation and measurement. Likewise, many different materials and electrode designs have been engineered to improve biocompatibility, such as printing MEAs on soft substrates mechanically similar to brain tissue and manufacturing electrodes using titanium nitride or ruthenium oxide (145-147). Furthermore, mesh nanoelectronics expedites the electrical characterization of organoids throughout the entire developmental time course. Li et al. (2019) first demonstrated ‘cyborg organoids,’ by integrating a stretchable layer containing electrodes in a mesh structure to a cardiomyocyte monolayer prepared for a cardiac organoid generation (148). During the organoid genesis, electrodes could be evenly spaced across the entire organoid, and electrophysiological measurements could be conducted all the time in single-cell resolution. Cyborg brain organoids were implemented into the brain organoid system to enable long-term non-invasive recording of neuronal activities (149). These approaches are part of bioinspired flexible electronics, enabling the safe establishment of neural interfaces and chronic recording from the entire organoid (150). This also includes the engineering of nanoelectronic scaffold (nanoES), invented by Tian et al. (2012), which provides structural and bioactive support for better mimicry of tissue organization and simultaneous monitoring of electrical activity (151). Moreover, neuron-like electronics (NeuE) can be applied to brain organoids (152), incorporating neural probes that resemble neurons in size, shape, and mechanical properties into neural tissues for high-resolution electrophysiological recording.
Long-term stable recording of brain organoid electrophysiology became a reality thanks to the development of various bioelectronic interfaces. However, some issues still remain in common. First, spatial mapping of electrophysiological activity is yet blurred. Recently, Miccoli et al. (2019) visualized the spatial distribution of electrical activity in neuronal monolayer culture with a 2D multimodal complementary metal-oxide-semiconductor (CMOS)-based MEA (153). This high-resolution MEA measured the impedance of each unit during the electrical recording. Impedance variation was calculated and used as an indicator of electrical activity, later constructed as an electrical map, demonstrating label-free, real-time and non-invasive visualization of the electrical activity in a neuronal network. Similar approaches in the 3D MEAs may clarify this map in the brain organoids. Second, cell-type specificity is lost in bioelectronic platforms. Li et al. (2021) recently succeeded to integrate single-cell electrophysiology with in situ sequencing, proposing in situ electro-seq (154). Utilizing the cyborg organoid platform, they labeled each electrode in the flexible mesh electronics with a unique fluorescent electronic barcode (E-barcode) by photolithography to represent the location of the electrode across the organoid. Whole organoid-electronics hybrid was fixed after electrical recording and cleared for in situ sequencing, leading to the explication of spatially-resolved RNA expression at the single-cell level. Optogenetics may enrich the knowledge in this field by manipulating the electrical activity of specific neurons (155, 156). Chemo-genetics, represented by DREADDs (designer receptor exclusively activated by designer drugs), may also enable the functional segregation of neuronal subtypes in brain organoids (157, 158). Taken together, thorough and systemic electrical characterization of brain organoids will contribute to our understanding of the process of neural circuitry formation in human brain development and brain disorders.
Go to :
Conclusions
Brain organoids nonetheless have opened up a new era of human neurobiology with unprecedented opportunities to study human brain development and disease in a dish. The breakthroughs of brain organoid technology in the past years have witnessed its magnificent potential not only as a neurodevelopmental model but also to resemble mature neuronal circuitry and functionality for the postnatal stages of the brain. In addition, the cutting-edge analysis tools to dissect the molecular, cellular, and functional aspects of brain organoids have further empowered researchers to unravel the mysteries of the complicated nature of the brain. We envision that the synergistic combination of bioengineering and stem cell technology will undoubtfully provide a more rigorous in vitro model of the brain in the near future. Lastly, we must keep in mind that while the upcoming brain organoid technology starts to emulate the human brain, to levels of reaching genuine consciousness, the technology may put itself at the edge of ethical standards. Still, it is an exciting avenue in neuroscience to better understand our brain and pave a way for treating brain disorders formerly incurable.
Go to :
 XML Download
XML Download